Multi-Omic Identification of Venom Proteins Collected from Artificial Hosts of a Parasitoid Wasp

 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
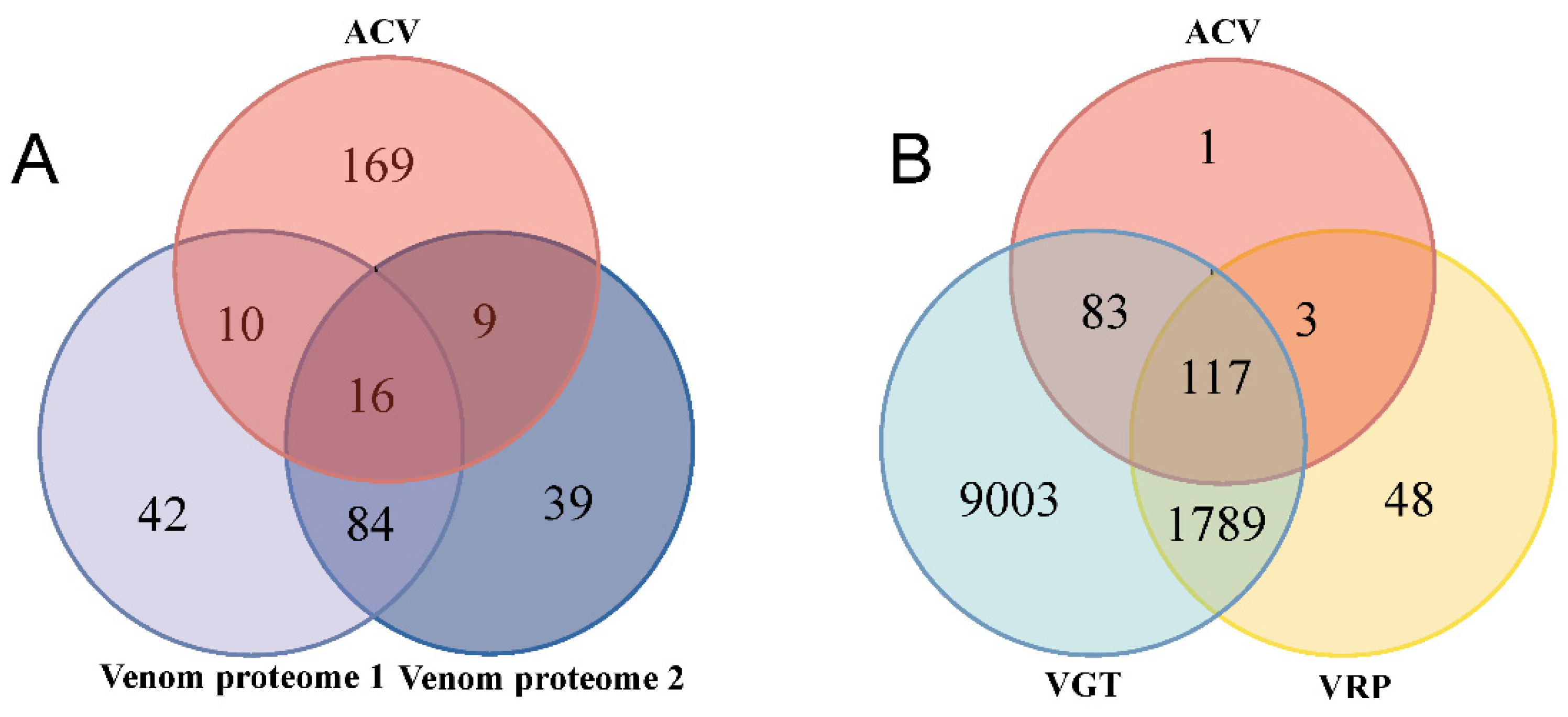
2.1. Identification and Analysis of Artificially Collected Venom Proteins Using Method 1
2.2. Identification of Putative Venom Proteins Using Method 2
2.3. Identification of Putative Venom Proteins Using Method 3
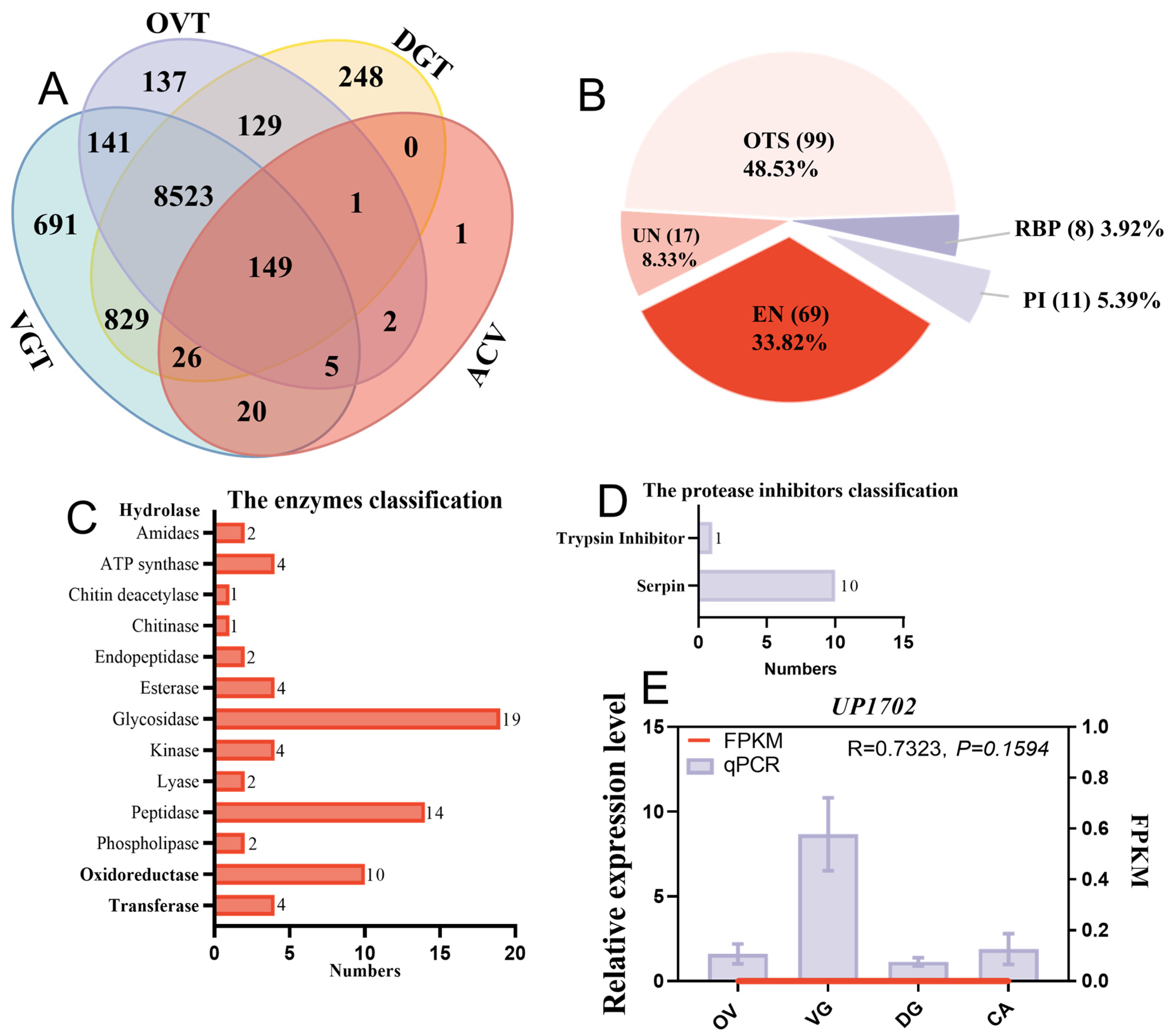
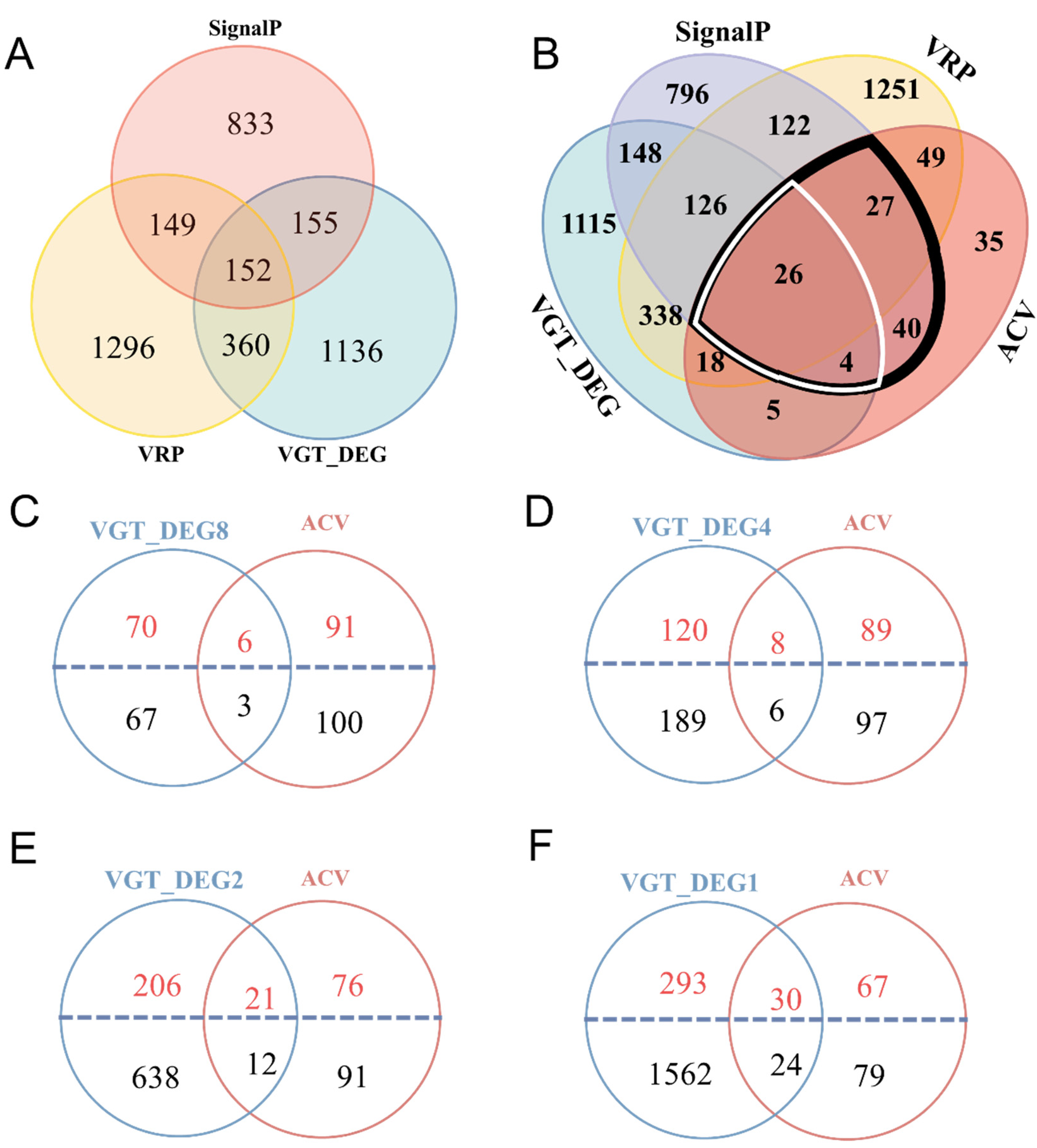
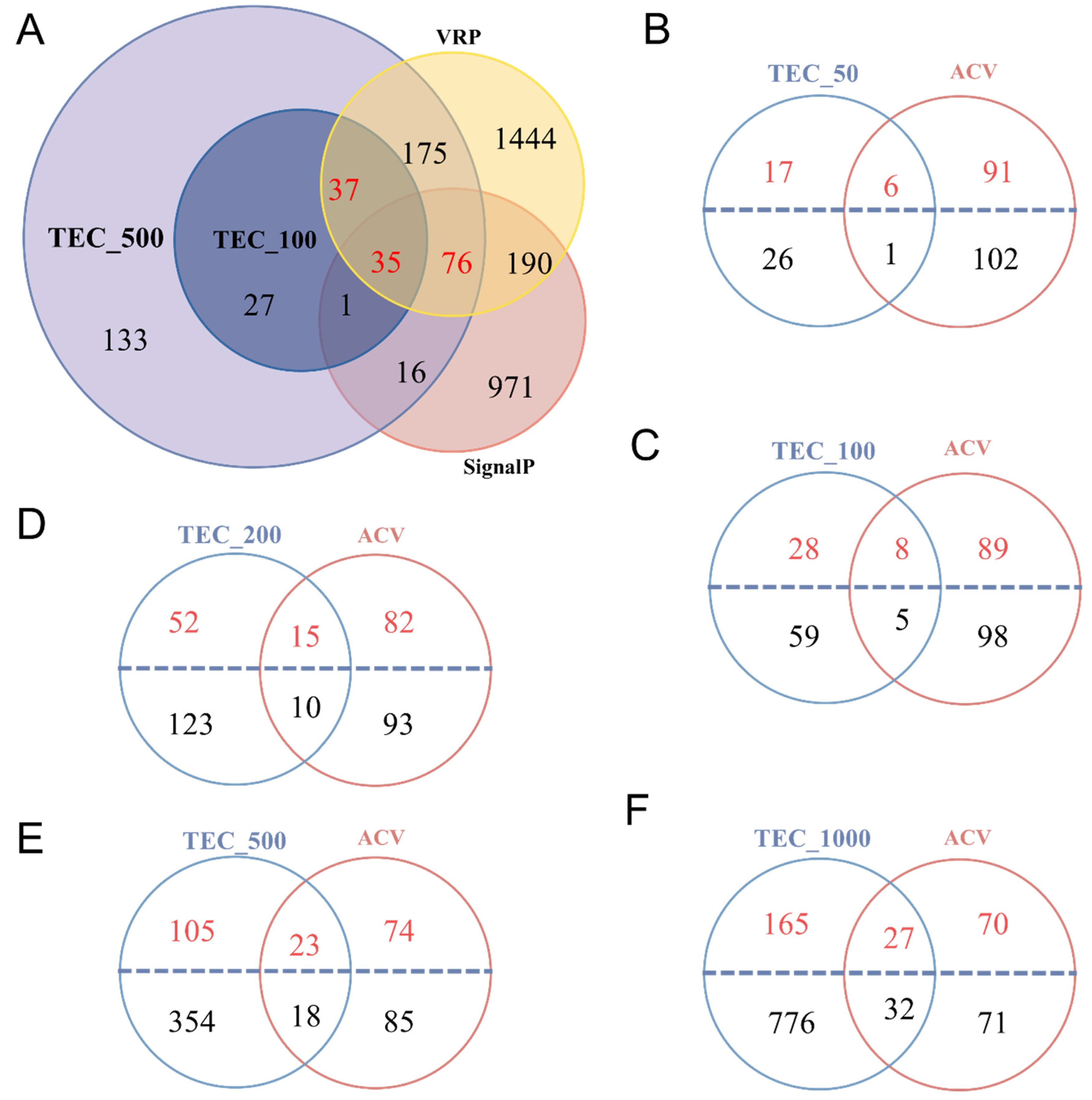
2.4. Comparison of Venom Putative Proteins Identified Using Different Methods
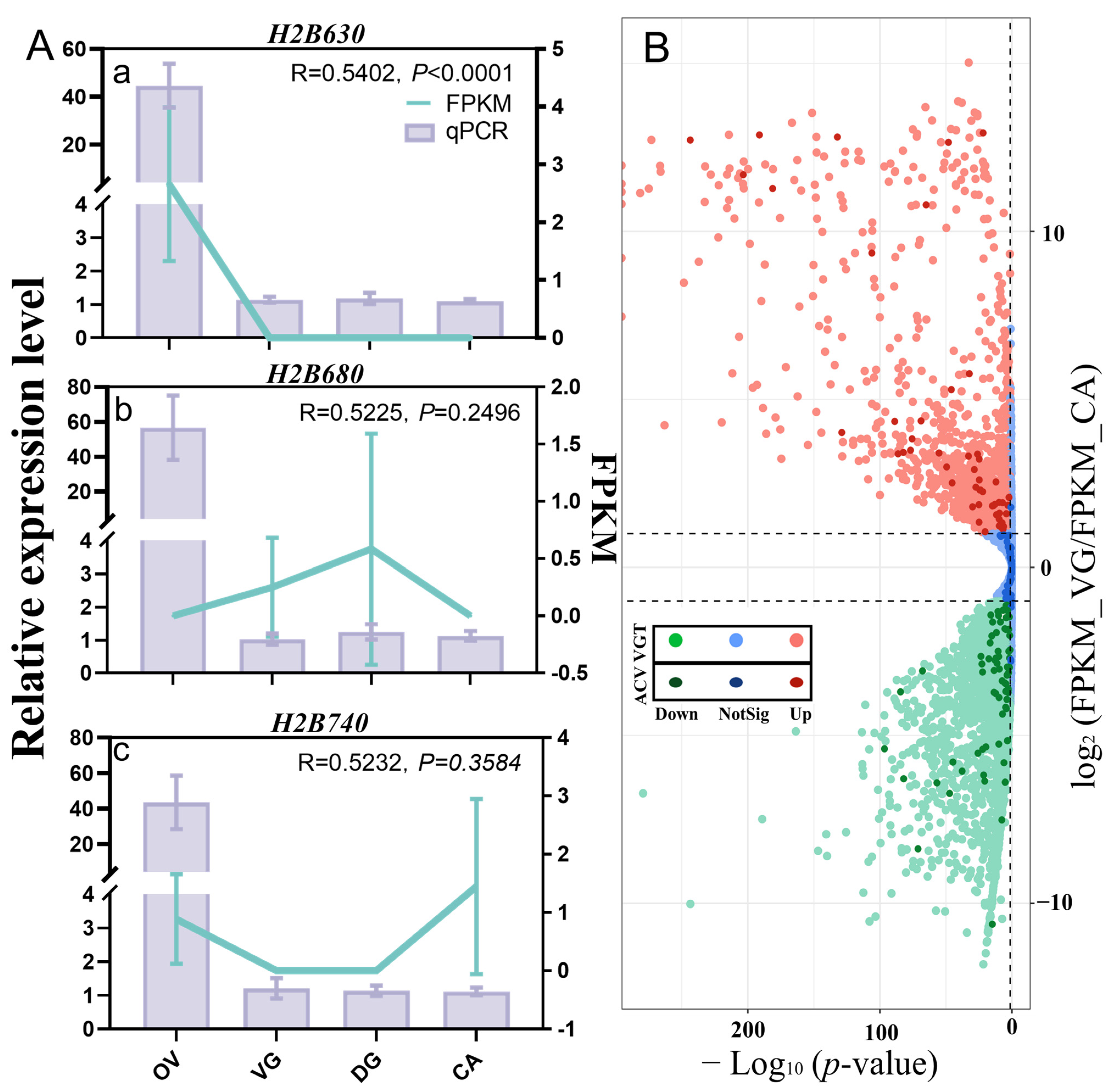
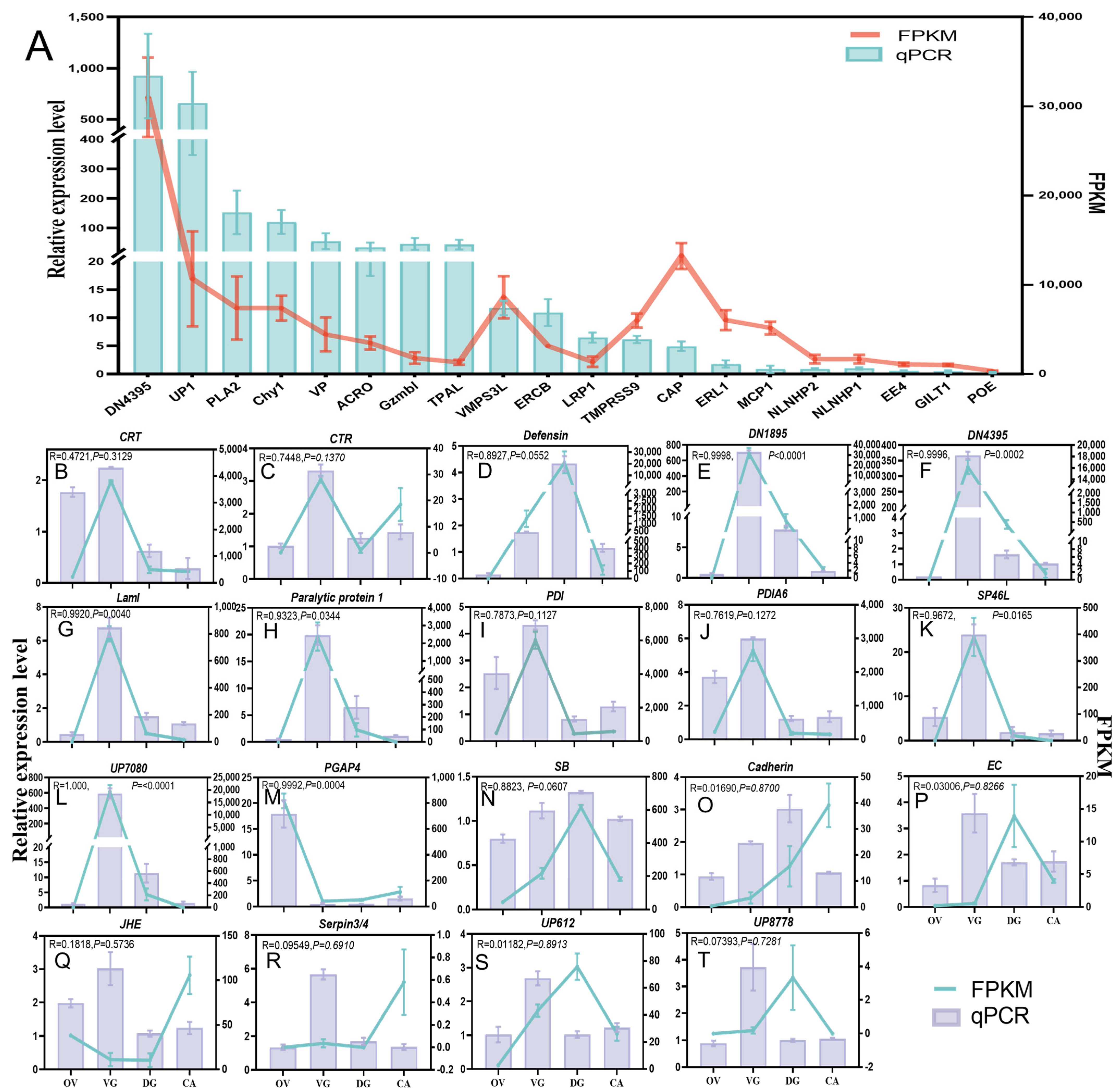
2.5. qPCR and PCR for Gene Accuracy
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Insect Breeding and Parasitization
5.2. RNA Preparation, Complementary DNA (cDNA) Library Construction, and Illumina® Sequencing
5.3. Read Assembly and Unigene Annotation
5.4. Transcriptomic Data Analysis
5.5. Protein Collection
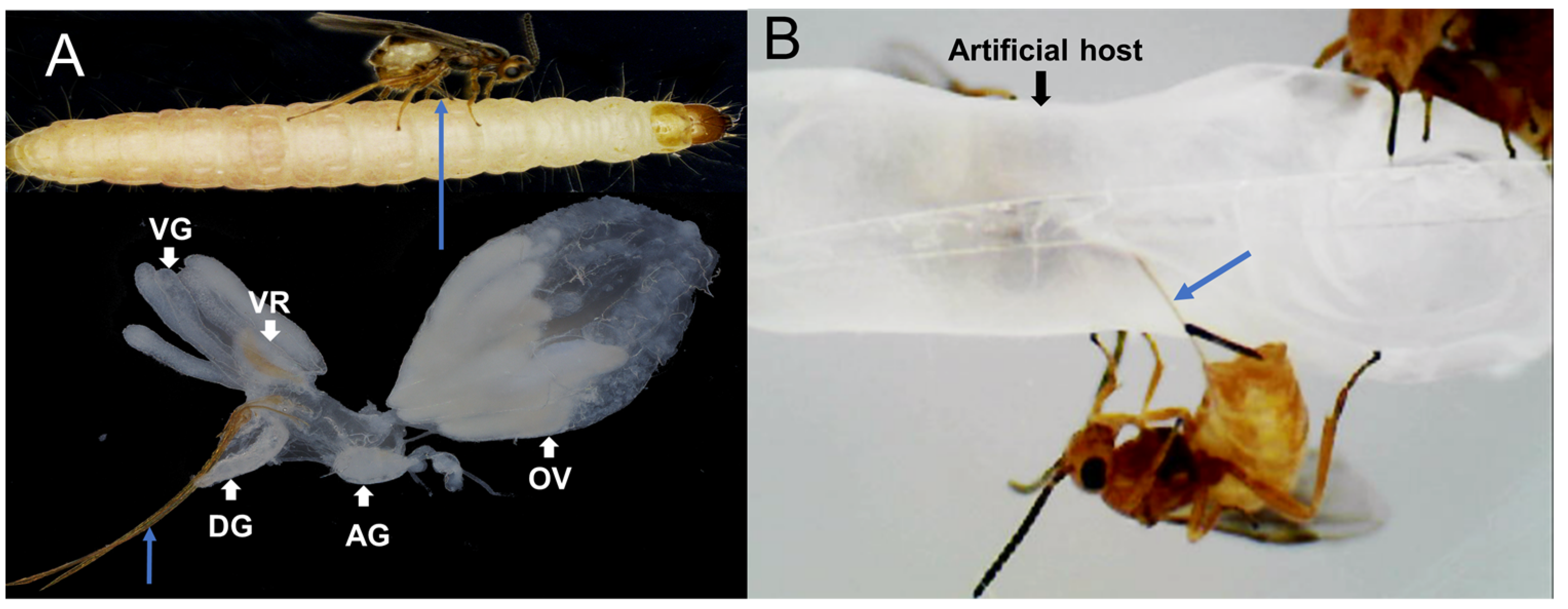
5.6. Artificial Host Production and Venom Protein Collection
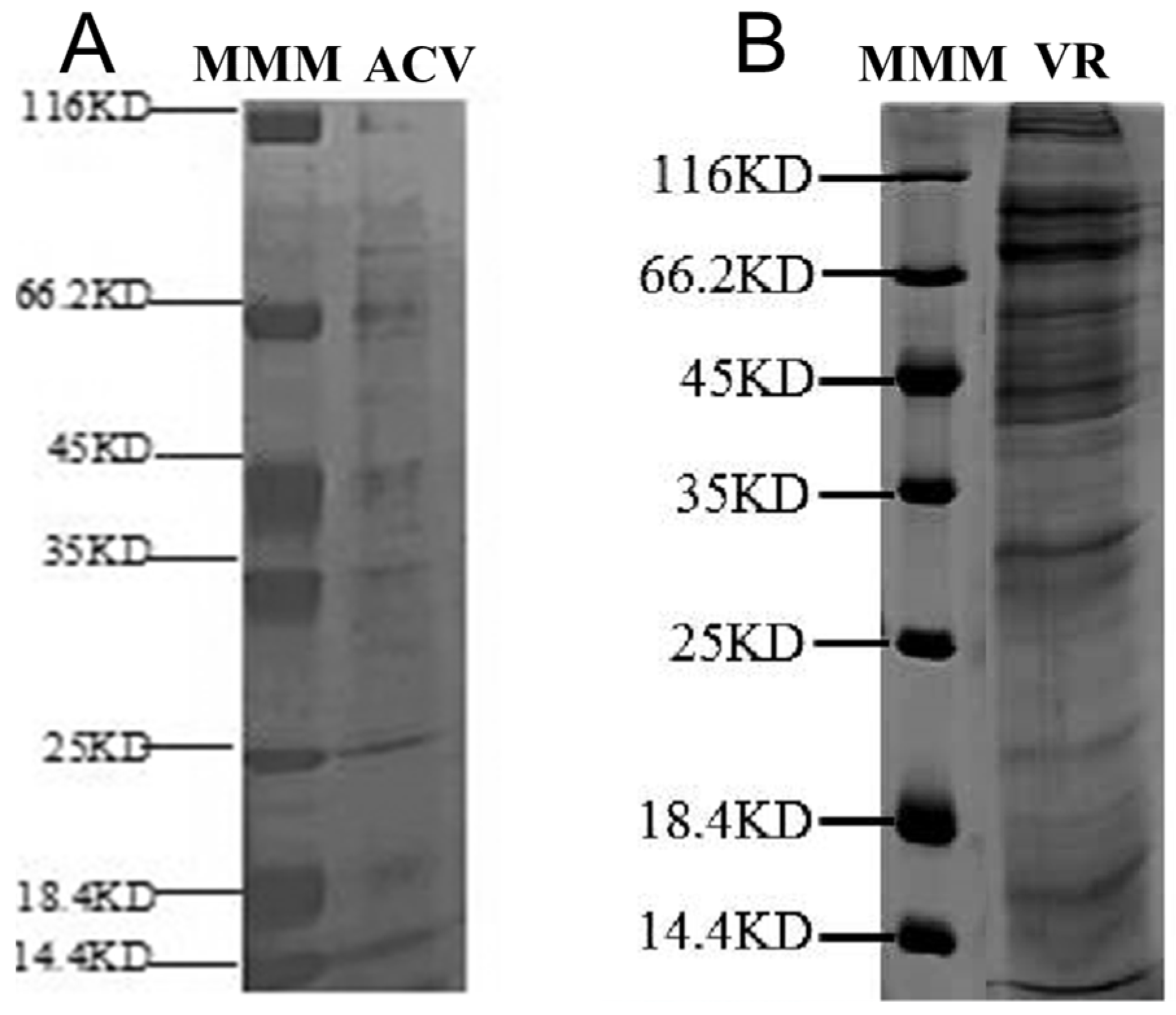
5.7. Analysis of Proteins from VR and ACV Using SDS-PAGE and LC-MS/MS
5.8. Venom Protein Identification
5.9. Compare with the Venom Proteins Screened by Method 2 from Six Parasitoid Wasps
5.10. Predict the Venom Proteins Families
5.11. Multiple Sequence Matching
5.12. qPCR
5.13. Sequencing Confirmation
5.14. Crude Venom Injection
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darling, D.C.; Sharkey, M.J. Order hymenoptera. Bull. Am. Mus. Nat. Hist. 1990, 195, 123–153. [Google Scholar]
- Laurino, S.; Grossi, G.; Pucci, P.; Flagiello, A.; Bufo, S.A.; Bianco, G.; Salvia, R.; Vinson, S.B.; Vogel, H.; Falabella, P. Identification of major Toxoneuron nigriceps venom proteins using an integrated transcriptomic/proteomic approach. Insect Biochem. Mol. Biol. 2016, 76, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Özbek, R.; Wielsch, N.; Vogel, H.; Lochnit, G.; Foerster, F.; Vilcinskas, A.; von Reumont, B.M. Proteo-transcriptomic characterization of the venom from the endoparasitoid wasp Pimpla turionellae with aspects on its biology and evolution. Toxins 2019, 11, 721. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, J.J.; Fernandez-Triana, J.L.; Smith, M.A.; Janzen, D.H.; Hallwachs, W.; Erwin, T.L.; Whitfield, J.B. Extrapolations from field studies and known faunas converge on dramatically increased estimates of global microgastrine parasitoid wasp species richness (Hymenoptera: Braconidae). Insect Conserv. Divers. 2013, 6, 530–536. [Google Scholar] [CrossRef]
- Wang, Z.-Z.; Liu, Y.-Q.; Shi, M.; Huang, J.-H.; Chen, X.-X. Parasitoid wasps as effective biological control agents. J. Integr. Agric. 2019, 18, 705–715. [Google Scholar] [CrossRef] [Green Version]
- von Reumont, B.M.; Campbell, L.I.; Jenner, R.A. Quo vadis venomics? A roadmap to neglected venomous invertebrates. Toxins 2014, 6, 3488–3551. [Google Scholar] [CrossRef] [Green Version]
- Kote, S.; Faktor, J.; Dapic, I.; Mayordomo, M.Y.; Kocikowski, M.; Kagansky, A.; Goodlett, D.; Vojtesek, B.; Hupp, T.; Wilcockson, D.; et al. Analysis of venom sac constituents from the solitary, aculeate wasp Cerceris rybyensis. Toxicon 2019, 169, 1–4. [Google Scholar] [CrossRef]
- Wang, H.-Z.; Zhong, X.; Gu, L.; Li, S.-S.; Zhang, G.-R.; Liu, X. Analysis of the Gynaephora qinghaiensis pupae immune transcriptome in response to parasitization by Thektogaster sp. Arch. Insect Biochem. Physiol. 2019, 100, e21533. [Google Scholar] [CrossRef]
- Beckage, N.E.; Gelman, D.B. Wasp parasitoid disruption of host development: Implications for new biologically based strategies for insect control. Annu. Rev. Entomol. 2004, 49, 299–330. [Google Scholar] [CrossRef] [Green Version]
- Moreau, S.J.M.; Asgari, S. Venom proteins from parasitoid wasps and their biological functions. Toxins 2015, 7, 2385–2412. [Google Scholar] [CrossRef] [Green Version]
- Elias, L.G.; Silva, D.B.; Silva, R.; Peng, Y.-Q.; Yang, D.-R.; Lopes, N.P.; Pereira, R.A.S. A comparative venomic fingerprinting approach reveals that galling and non-galling fig wasp species have different venom profiles. PLoS ONE 2018, 13, e0207051. [Google Scholar] [CrossRef]
- Asgari, S.; Rivers, D.B. Venom proteins from endoparasitoid wasps and their role in host-parasite interactions. Annu. Rev. Entomol. 2011, 56, 313–335. [Google Scholar] [CrossRef] [PubMed]
- Piek, T.; Veenendaal, R.L.; Mantel, P. The pharmacology of Microbracon venom. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1982, 72, 303–309. [Google Scholar] [CrossRef]
- Richards, E.H.; Edwards, J.P. Larvae of the ectoparasitic wasp, Eulophus pennicornis, release factors which adversely affect haemocytes of their host, Lacanobia oleracea. J. Insect Physiol. 2002, 48, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Rivers, D.B.; Ruggiero, L.; Hayes, M. The ectoparasitic wasp Nasonia vitripennis (Walker) (Hymenoptera: Pteromalidae) differentially affects cells mediating the immune response of its flesh fly host, Sarcophaga bullata Parker (Diptera: Sarcophagidae). J. Insect Physiol. 2002, 48, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Martinson, E.O.; Wheeler, D.; Wright, J.; Mrinalini; Siebert, A.L.; Werren, J.H. Nasonia vitripennis venom causes targeted gene expression changes in its fly host. Mol. Ecol. 2014, 23, 5918–5930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrinalini; Siebert, A.L.; Wright, J.; Martinson, E.; Wheeler, D.; Werren, J.H. Parasitoid venom induces metabolic cascades in fly hosts. Metabolomics 2015, 11, 350–366. [Google Scholar] [CrossRef] [Green Version]
- Asgari, S. Venom proteins from polydnavirus-producing endoparasitoids: Their role in host-parasite interactions. Arch. Insect Biochem. Physiol. 2006, 61, 146–156. [Google Scholar] [CrossRef]
- Teng, Z.-W.; Xiong, S.-J.; Xu, G.; Gan, S.-Y.; Chen, X.; Stanley, D.; Yan, Z.-C.; Ye, G.-Y.; Fang, Q. Protein discovery: Combined transcriptomic and proteomic analyses of venom from the endoparasitoid Cotesia chilonis (Hymenoptera: Braconidae). Toxins 2017, 9, 135. [Google Scholar] [CrossRef] [Green Version]
- Moreau, S.J.M.; Guillot, S. Advances and prospects on biosynthesis, structures and functions of venom proteins from parasitic wasps. Insect Biochem. Mol. Biol. 2005, 35, 1209–1223. [Google Scholar] [CrossRef]
- Mendonça, A.; Marchiotti, R.C.B.; Firmino, E.L.B.; Santos, P.P.; Antonio, D.S.; Serrão, J.E.; Cardoso, C.A.L.; Antonialli Junior, W.F. Proteomic analysis of the venom of the social wasp Apoica pallens (Hymenoptera: Vespidae). Rev. Bras. Entomol. 2019, 63, 322–330. [Google Scholar] [CrossRef]
- Wilson, D.; Daly, N.L. Venomics: A mini-review. High-Throughput 2018, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- von Reumont, B.M.; Anderluh, G.; Antunes, A.; Ayvazyan, N.; Beis, D.; Caliskan, F.; Crnkovic, A.; Damm, M.; Dutertre, S.; Ellgaard, L.; et al. Modern venomics-current insights, novel methods, and future perspectives in biological and applied animal venom research. Gigascience 2022, 11, giac048. [Google Scholar] [CrossRef]
- Vincent, B.; Kaeslin, M.; Roth, T.; Heller, M.; Poulain, J.; Cousserans, F.; Schaller, J.; Poirié, M.; Lanzrein, B.; Drezen, J.-M.; et al. The venom composition of the parasitic wasp Chelonus inanitus resolved by combined expressed sequence tags analysis and proteomic approach. BMC Genom. 2010, 11, 693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorémus, T.; Urbach, S.; Jouan, V.; Cousserans, F.; Ravallec, M.; Demettre, E.; Wajnberg, E.; Poulain, J.; Azéma-Dossat, C.; Darboux, I.; et al. Venom gland extract is not required for successful parasitism in the polydnavirus-associated endoparasitoid Hyposoter didymator (Hym. Ichneumonidae) despite the presence of numerous novel and conserved venom proteins. Insect Biochem. Mol. Biol. 2013, 43, 292–307. [Google Scholar] [CrossRef]
- Colinet, D.; Anselme, C.; Deleury, E.; Mancini, D.; Poulain, J.; Azéma-Dossat, C.; Belghazi, M.; Tares, S.; Pennacchio, F.; Poirié, M.; et al. Identification of the main venom protein components of Aphidius ervi, a parasitoid wasp of the aphid model Acyrthosiphon pisum. BMC Genom. 2014, 15, 342. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.C.; Fang, Q.; Wang, L.; Liu, J.; Zhu, Y.; Wang, F.; Li, F.; Werren, J.H.; Ye, G. Insights into the venom composition and evolution of an endoparasitoid wasp by combining proteomic and transcriptomic analyses. Sci. Rep. 2016, 6, 19604–19616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Yang, Y.; Liu, M.-M.; Yan, Z.-C.; Qiu, L.-M.; Fang, Q.; Wang, F.; Werren, J.H.; Ye, G.-Y. Identification and comparative analysis of venom proteins in a pupal ectoparasitoid, Pachycrepoideus vindemmiae. Front. Physiol. 2020, 11, 9–27. [Google Scholar] [CrossRef]
- Scieuzo, C.; Salvia, R.; Franco, A.; Pezzi, M.; Cozzolino, F.; Chicca, M.; Scapoli, C.; Vogel, H.; Monti, M.; Ferracini, C.; et al. An integrated transcriptomic and proteomic approach to identify the main Torymus sinensis venom components. Sci. Rep. 2021, 11, 5032. [Google Scholar] [CrossRef] [PubMed]
- de Graaf, D.C.; Aerts, M.; Brunain, M.; Desjardins, C.A.; Jacobs, F.J.; Werren, J.H.; Devreese, B. Insights into the venom composition of the ectoparasitoid wasp Nasonia vitripennis from bioinformatic and proteomic studies. Insect Mol. Biol. 2010, 19 (Suppl. S1), 11–26. [Google Scholar] [CrossRef] [Green Version]
- Martinson, E.O.; Mrinalini; Kelkar, Y.D.; Chang, C.-H.; Werren, J.H. The evolution of venom by co-option of single-copy genes. Curr. Biol. 2017, 27, 2007–2013.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heavner, M.E.; Gueguen, G.; Rajwani, R.; Pagan, P.E.; Small, C.; Govind, S. Partial venom gland transcriptome of a Drosophila parasitoid wasp, Leptopilina heterotoma, reveals novel and shared bioactive profiles with stinging Hymenoptera. Gene 2013, 526, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Zhang, L.; Fang, Y.; Han, B.; Lu, X.; Zhou, T.; Feng, M.; Li, J. Proteome and phosphoproteome analysis of honeybee (Apis mellifera) venom collected from electrical stimulation and manual extraction of the venom gland. BMC Genom. 2013, 14, 766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turillazzi, F.; Pieraccini, G.; Turillazzi, S.; Battaglini, N.O.; Severino, M. Venom collection by electrical stimulation in the invasive species Polistes dominula reared using a vespiculture regime. Molecules 2022, 27, 8821. [Google Scholar] [CrossRef] [PubMed]
- Kryukova, N.A.; Chertkova, E.A.; Semenova, A.D.; Glazachev, Y.I.; Slepneva, I.A.; Glupov, V.V. Venom from the ectoparasitic wasp Habrobracon hebetor activates calcium-dependent degradation of Galleria mellonella Larval hemocytes. Arch. Insect Biochem. Physiol. 2015, 90, 117–130. [Google Scholar] [CrossRef]
- Ghimire, M.N.; Phillips, T.W. Mass rearing of Habrobracon hebetor Say (Hymenoptera: Braconidae) on larvae of the Indian meal moth, Plodia interpunctella (Lepidoptera: Pyralidae): Effects of host density, parasitoid density, and rearing containers. J. Stored Prod. Res. 2010, 46, 214–220. [Google Scholar] [CrossRef]
- Mbata, G.N.; Warsi, S. Habrobracon hebetor and Pteromalus cerealellae as tools in post-harvest integrated pest management. Insects 2019, 10, 85. [Google Scholar] [CrossRef] [Green Version]
- Quistad, G.B.; Nguyen, Q.; Bernasconi, P.; Leisy, D.J. Purification and characterization of insecticidal toxins from venom glands of the parasitic wasp, Bracon hebetor. Insect Biochem. Mol. Biol. 1994, 24, 955–961. [Google Scholar] [CrossRef]
- Manzoor, A.; Ul Abdin, Z.; Webb, B.A.; Arif, M.J.; Jamil, A. De novo sequencing and transcriptome analysis of female venom glands of ectoparasitoid Bracon hebetor (Say.) (Hymenoptera: Braconidae). Comp. Biochem. Physiol. Part D Genom. Proteom. 2016, 20, 101–110. [Google Scholar] [CrossRef]
- Hussain, F.; Ul Abdin, Z.; Arif, M.J.; Jamil, A.; Li, X. Isolation and characterization of immune suppressive genes through bioinformatic analysis of venom glands transcriptome of Bracon hebetor (Hymenoptera: Braconidae). Int. J. Agric. Biol. 2019, 21, 1189–1196. [Google Scholar] [CrossRef]
- Ye, X.H.; Yang, Y.; Tian, Z.Y.; Xu, L.; Yu, K.L.; Xiao, S.; Yin, C.L.; Xiong, S.J.; Fang, Q.; Chen, H.; et al. A high-quality de novo genome assembly from a single parasitoid wasp. BioRxiv 2020. [Google Scholar] [CrossRef]
- Quicke, D.L.J.; Butcher, B.A. Review of venoms of non-polydnavirus carrying Ichneumonoid wasps. Biology 2021, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Eng, J.K.; Mccormack, A.L.; Yates, J.R. An approach to correlate tandem mass spectral data of peptides with amino acid sequences in a protein database. J. Am. Soc. Mass Spectrom. 1994, 5, 976–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Wang, R.-J.; Cheng, Y.; Du, J.; Volovych, O.; Han, L.-B.; Li, J.-C.; Hu, Y.; Lu, Z.-Y.; Lu, Z.; et al. Insights into the venom protein components of Microplitis mediator, an endoparasitoid wasp. Insect Biochem. Mol. Biol. 2019, 105, 33–42. [Google Scholar] [CrossRef]
- Jones, D.; Sawicki, G.; Wozniak, M. Sequence, structure, and expression of a wasp venom protein with a negatively charged signal peptide and a novel repeating internal structure. J. Biol. Chem. 1992, 267, 14871–14878. [Google Scholar] [CrossRef]
- Ferrarese, R.; Morales, J.; Fimiarz, D.; Webb, B.A.; Govind, S. A supracellular system of actin-lined canals controls biogenesis and release of virulence factors in parasitoid venom glands. J. Exp. Biol. 2009, 212, 2261–2268. [Google Scholar] [CrossRef] [Green Version]
- Aili, S.R.; Touchard, A.; Hayward, R.; Robinson, S.D.; Pineda, S.S.; Lalagüe, H.; Mrinalini; Vetter, I.; Undheim, E.A.B.; Kini, R.M.; et al. An integrated proteomic and transcriptomic analysis reveals the venom complexity of the bullet ant Paraponera clavata. Toxins 2020, 12, 324. [Google Scholar] [CrossRef]
- Uçkan, F.; Ergin, E.; Rivers, D.B.; Gençer, N. Age and diet influence the composition of venom from the endoparasitic wasp Pimpla turionellae L. (Hymenoptera: Ichneumonidae). Arch. Insect Biochem. Physiol. 2006, 63, 177–187. [Google Scholar] [CrossRef]
- Silva, L.M.; Botelho, A.C.C.; Nacif-Pimenta, R.; Martins, G.F.; Alves, L.C.; Brayner, F.A.; Fortes-Dias, C.L.; Pimenta, P.F.P. Structural analysis of the venom glands of the armed spider Phoneutria nigriventer (Keyserling, 1891): Microanatomy, fine structure and confocal observations. Toxicon 2008, 51, 693–706. [Google Scholar] [CrossRef]
- Wan, B.; Poirié, M.; Gatti, J.-L. Parasitoid wasp venom vesicles (venosomes) enter Drosophila melanogaster lamellocytes through a flotillin/lipid raft-dependent endocytic pathway. Virulence 2020, 11, 1512–1521. [Google Scholar] [CrossRef] [PubMed]
- Windass, J.D.; Christian, P.D.; Duncan, R.E.; Baule, V.J. Toxins from the Wasp Bracon hebetor. International Application No. PCT/GB1995/002720, 30 May 1996. [Google Scholar]
- Hou, C.S.; Guo, L.Q.; You, L.F.; Wang, J.R.; Lin, J.F.; Wu, W.H.; Wu, C.S.; Wang, T. Molecular cloning and characterization of a venom phospholipase A2 from Apis mellifera spp. J. Entomol. Res. Soc. 2014, 16, 55–66. [Google Scholar]
- Xin, B.; Liu, P.; Xu, X.; Zhang, S.; Zheng, Y. Identification of venom proteins of the indigenous endoparasitoid Chouioia cunea (Hymenoptera: Eulophidae). J. Econ. Entomol. 2017, 110, 2022–2030. [Google Scholar] [CrossRef] [PubMed]
- Leisy, D.J.; Quistad, G.B. Insecticidal Toxins from the Parastic Wasp, Bracon hebetor. International Application No. PCT/EP1993/000431, 16 September 1993. [Google Scholar]
- Zurovec, M.; Martinkova, B.; Zaloudikova, A.; Shaik, H.A.; Konik, P.; Strnad, H.; Sehadova, H.; Kodrik, D. Isolation and Characterization of the Gene Encoding a Paralytic Protein from the Wasp Habrobracon hebetor. Unpublished Submitted (30-DEC-2017) Genetics, Biology Centre, CAS, Institute of Entomology, Branisovska 31, Ceske Budejovice, Czech Republic 37005, Czech Republic. 2017. Available online: https://www.ncbi.nlm.nih.gov/protein/?term=Isolation%20and%20Characterization%20of%20the%20gene%20encoding%20a%20paralytic%20protein%20from%20the%20wasp%20Habrobracon%20hebetor (accessed on 14 May 2023).
- Colinet, D.; Mathé-Hubert, H.; Allemand, R.; Gatti, J.-L.; Poirié, M. Variability of venom components in immune suppressive parasitoid wasps: From a phylogenetic to a population approach. J. Insect Physiol. 2013, 59, 205–212. [Google Scholar] [CrossRef]
- Becchimanzi, A.; Avolio, M.; Bostan, H.; Colantuono, C.; Cozzolino, F.; Mancini, D.; Chiusano, M.L.; Pucci, P.; Caccia, S.; Pennacchio, F. Venomics of the ectoparasitoid wasp Bracon nigricans. BMC Genom. 2020, 21, 34. [Google Scholar] [CrossRef]
- Laskowski, M.; Kato, I. Protein inhibitors of proteinases. Annu. Rev. Biochem. 1980, 49, 593–626. [Google Scholar] [CrossRef]
- Liu, N.-Y.; Huang, J.-M.; Ren, X.-M.; Xu, Z.-W.; Yan, N.-S.; Zhu, J.-Y. Superoxide dismutase from venom of the ectoparasitoid Scleroderma guani inhibits melanization of hemolymph. Arch. Insect Biochem. Physiol. 2018, 99, e21503. [Google Scholar] [CrossRef]
- Li, X.; Qin, Q.; Ding, C.; Liu, H. Characterization of Microplitis mediator polydnavirus. Virol. Sin. 2001, 16, 373–376. [Google Scholar]
- Wu, S.-F.; Sun, F.-D.; Qi, Y.-X.; Yao, Y.; Fang, Q.; Huang, J.; Stanley, D.; Ye, G.-Y. Parasitization by Cotesia chilonis influences gene expression in fatbody and hemocytes of Chilo suppressalis. PLoS ONE 2013, 8, e74309. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.A.; Robinson, S.D.; Yeates, D.K.; Jin, J.; Baumann, K.; Dobson, J.; Fry, B.G.; King, G.F. Entomo-venomics: The evolution, biology and biochemistry of insect venoms. Toxicon 2018, 154, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Saba, E.; Shafeeq, T.; Irfan, M.; Lee, Y.Y.; Kwon, H.-W.; Seo, M.G.; Park, S.-J.; Lee, K.-Y.; Rhee, M.H. Anti-inflammatory activity of crude venom isolated from parasitoid wasp, Bracon hebetor Say. Mediat. Inflamm. 2017, 2017, 6978194. [Google Scholar] [CrossRef] [Green Version]
- Phillips, T.W.; Strand, M.R. Larval secretions and food odors affect orientation in female Plodia interpunctella. Entomol. Exp. Appl. 1994, 71, 185–192. [Google Scholar] [CrossRef]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–653. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.C.; Ren, X.Y.; Li, Y.X. A Method for Extracting Parasitic Wasp Venom Using Artificial Hosts. International Application No. CN111149772A, 15 May 2020. [Google Scholar]
- Li, L.; Stoeckert, C.J., Jr.; Roos, D.S. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, K.; Chen, J.; Bai, X.; Xiong, S.; Ye, X.; Yang, Y.; Yao, H.; Wang, F.; Fang, Q.; Song, Q.; et al. Multi-Omic Identification of Venom Proteins Collected from Artificial Hosts of a Parasitoid Wasp. Toxins 2023, 15, 377. https://doi.org/10.3390/toxins15060377
Yu K, Chen J, Bai X, Xiong S, Ye X, Yang Y, Yao H, Wang F, Fang Q, Song Q, et al. Multi-Omic Identification of Venom Proteins Collected from Artificial Hosts of a Parasitoid Wasp. Toxins. 2023; 15(6):377. https://doi.org/10.3390/toxins15060377
Chicago/Turabian StyleYu, Kaili, Jin Chen, Xue Bai, Shijiao Xiong, Xinhai Ye, Yi Yang, Hongwei Yao, Fang Wang, Qi Fang, Qisheng Song, and et al. 2023. "Multi-Omic Identification of Venom Proteins Collected from Artificial Hosts of a Parasitoid Wasp" Toxins 15, no. 6: 377. https://doi.org/10.3390/toxins15060377
APA StyleYu, K., Chen, J., Bai, X., Xiong, S., Ye, X., Yang, Y., Yao, H., Wang, F., Fang, Q., Song, Q., & Ye, G. (2023). Multi-Omic Identification of Venom Proteins Collected from Artificial Hosts of a Parasitoid Wasp. Toxins, 15(6), 377. https://doi.org/10.3390/toxins15060377