Microcystin-LR Induces and Aggravates Colitis through NLRP3 Inflammasome-Mediated Pyroptosis in Mice


 and
and 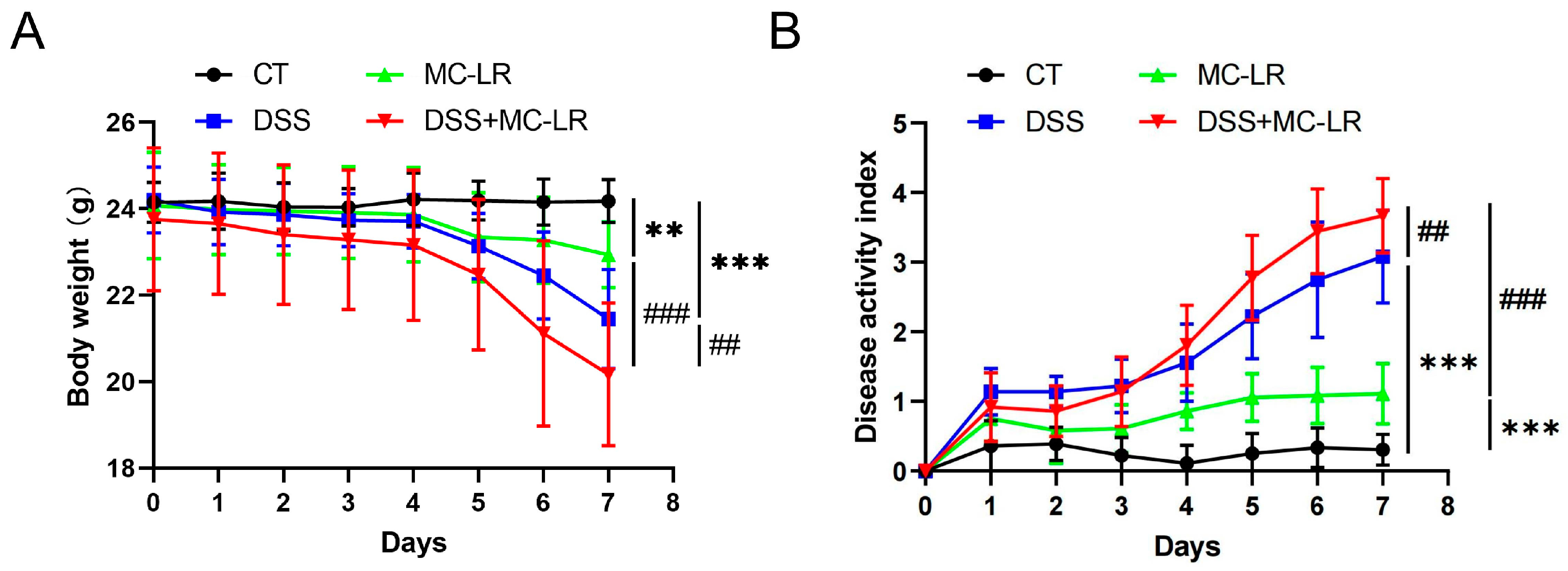{kind=link}
{kind=link}
{kind=link}
{kind=link}
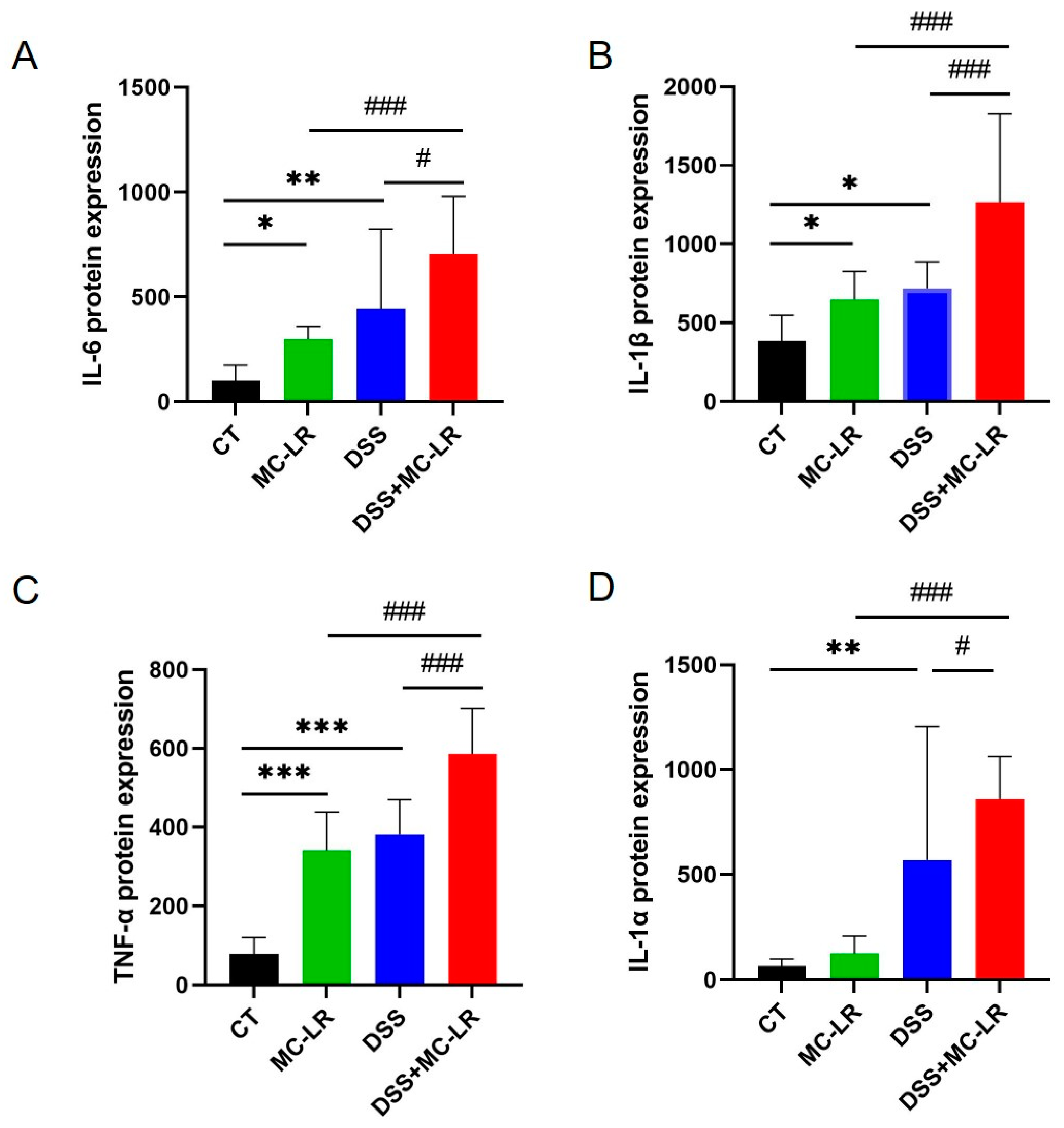{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Body Weight and DAI Score
2.2. Colon Length
2.3. Histological and Cytological Damage
2.4. The Expression Levels of Pro-Inflammatory Cytokines
2.5. NLRP3-Related Proteins
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Reagents and Antibodies
5.2. Mice
5.3. Modeling and Experimental Design
5.4. Histology
5.5. Enzyme-Linked Immunosorbent Assay (ELISA)
5.6. Western Blotting (WB)
5.7. TUNEL Staining
5.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flynn, S.; Eisenstein, S. Inflammatory bowel disease presentation and diagnosis. Surg. Clin. 2019, 99, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Lukas, M. Inflammatory bowel disease as a risk factor for colorectal cancer. Dig. Dis. 2010, 28, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Coward, S.; Clement, F.; Benchimol, E.; Bernstein, C.; Bitton, A.; Carroll, M.; Hazlewood, G.; Jelinski, S.; Jones, J.; Kuenzig, E.; et al. Analyzing the rising prevalence of IBD: Predicting the prevalence in 2030 by age group. Gastroenterology 2018, 154, S992–S993. [Google Scholar] [CrossRef]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.; Chan, F.K. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef]
- Kaplan, G.G. The global burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef]
- Benchimol, E.I.; Bernstein, C.N.; Bitton, A.; Murthy, S.K.; Nguyen, G.C.; Lee, K.; Cooke-Lauder, J.; Siddiq, S.; Windsor, J.W.; Carroll, M.W. The impact of inflammatory bowel disease in Canada 2018: A scientific report from the Canadian Gastro-Intestinal Epidemiology Consortium to Crohn’s and Colitis Canada. J. Can. Assoc. Gastroenterol. 2019, 2, S1–S5. [Google Scholar] [CrossRef] [Green Version]
- Podolsky, D.K. Inflammatory bowel disease. N. Engl. J. Med. 2002, 347, 417–429. [Google Scholar] [CrossRef]
- Kaplan, G.G.; Ng, S.C. Globalisation of inflammatory bowel disease: Perspectives from the evolution of inflammatory bowel disease in the UK and China. Lancet Gastroenterol. Hepatol. 2016, 1, 307–316. [Google Scholar] [CrossRef]
- Kaplan, G.G.; Ng, S.C. Understanding and Preventing the Global Increase of Inflammatory Bowel Disease. Gastroenterology 2017, 152, 313–321.e312. [Google Scholar] [CrossRef] [Green Version]
- Chorus, I. Introduction: Cyanotoxins—Research for environmental safety and human health. In Cyanotoxin; Springer: Berlin/Heidelberg, Germany, 2001; pp. 1–4. [Google Scholar]
- Campos, A.; Vasconcelos, V. Molecular mechanisms of microcystin toxicity in animal cells. Int. J. Mol. Sci. 2010, 11, 268–287. [Google Scholar] [CrossRef] [Green Version]
- Lone, Y.; Koiri, R.K.; Bhide, M. An overview of the toxic effect of potential human carcinogen Microcystin-LR on testis. Toxicol. Rep. 2015, 2, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Sedan, D.; Laguens, M.; Copparoni, G.; Aranda, J.O.; Giannuzzi, L.; Marra, C.A.; Andrinolo, D. Hepatic and intestine alterations in mice after prolonged exposure to low oral doses of Microcystin-LR. Toxicon 2015, 104, 26–33. [Google Scholar] [CrossRef]
- Su, R.C.; Blomquist, T.M.; Kleinhenz, A.L.; Khalaf, F.K.; Dube, P.; Lad, A.; Breidenbach, J.D.; Mohammed, C.J.; Zhang, S.; Baum, C.E. Exposure to the harmful algal bloom (HAB) toxin microcystin-LR (MC-LR) prolongs and increases severity of dextran sulfate sodium (DSS)-induced colitis. Toxins 2019, 11, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Wang, H.; Wang, X.; Chen, L.; Liu, W.; Cai, D.; Deng, S.; Chu, H.; Liu, Y.; Feng, X.; et al. Long-term environmental levels of microcystin-LR expo-sure induces colorectal chronic inflammation, fibrosis and barrier disruption via CSF1R/Rap1b signaling pathway. J. Hazard. Mater. 2022, 440, 129793. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Hoeger, S.J.; Stemmer, K.; Feurstein, D.J.; Knobeloch, D.; Nussler, A.; Dietrich, D.R. The role of organic anion transporting polypeptides (OATPs/SLCOs) in the toxicity of different microcystin congeners in vitro: A comparison of primary human hepatocytes and OATP-transfected HEK293 cells. Toxicol. Appl. Pharm. 2010, 245, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Valerio, E.; Vasconcelos, V.; Campos, A. New Insights on the Mode of Action of Microcystins in Animal Cells—A Review. Mini Rev. Med. Chem. 2016, 16, 1032–1041. [Google Scholar] [CrossRef] [PubMed]
- Bouaicha, N.; Maatouk, I. Microcystin-LR and nodularin induce intracellular glutathione alteration, reactive oxygen species production and lipid peroxidation in primary cultured rat hepatocytes. Toxicol. Lett. 2004, 148, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Kitani, A.; Strober, W.; Fuss, I.J. The role of NLRP3 and IL-1β in the pathogenesis of inflammatory bowel disease. Front. Immunol. 2018, 9, 2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, L.; Li, Y.W.; Liu, B.L.; Zhao, H.M.; Li, H.; Cai, Q.Y.; Mo, C.H.; Wong, M.H.; Li, Q.X. High ecological and human health risks from microcystins in vegetable fields in southern China. Environ. Int. 2019, 133, 105142. [Google Scholar] [CrossRef] [PubMed]
- Greer, B.; Meneely, J.P.; Elliott, C.T. Uptake and accumulation of Microcystin-LR based on exposure through drinking water: An animal model assessing the human health risk. Sci. Rep. UK 2018, 8, 4913. [Google Scholar] [CrossRef] [Green Version]
- Lun, Z.; Hai, Y.; Kun, C. Relationship between microcystin in drinking water and colorectal cancer. Biomed. Environ. Sci. 2002, 15, 166–171. [Google Scholar]
- Dong, S.; Zhu, M.; Wang, K.; Zhao, X.; Hu, L.; Jing, W.; Lu, H.; Wang, S. Dihydromyricetin improves DSS-induced colitis in mice via modulation of fecal-bacteria-related bile acid metabolism. Pharm. Res. 2021, 171, 105767. [Google Scholar] [CrossRef]
- Su, R.C.; Warner, E.A.; Breidenbach, J.D.; Lad, A.; Blomquist, T.M.; Kleinhenz, A.L.; Modyanov, N.; Malhotra, D.; Kennedy, D.J.; Haller, S.T. CD40 receptor knockout protects against microcystin-LR (MC-LR) prolongation and exacerbation of dextran sulfate sodium (DSS)-induced colitis. Biomedicines 2020, 8, 149. [Google Scholar] [CrossRef]
- Bakr, A.; Alzain, M.N.; Alzamel, N.M.; Loutfy, N. Accumulation of Microcystin from Oscillatoria limnetica Lemmermann and Microcystis aeruginosa (Kützing) in Two Leafy Green Vegetable Crop Plants Lactuca sativa L. and Eruca sativa. Plants 2022, 11, 1733. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xie, P.; Lin, J.; He, J.; Zeng, C.; Chen, J. Effects of microcystin-LR on gut microflora in different gut regions of mice. J. Toxicol. Sci. 2015, 40, 485–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Giesy, J.P.; Xie, P. The dose makes the poison. Sci. Total. Environ. 2018, 621, 649–653. [Google Scholar] [CrossRef]
- Jia, J.; Luo, W.; Lu, Y.; Giesy, J.P. Bioaccumulation of microcystins (MCs) in four fish species from Lake Taihu, China: Assessment of risks to humans. Sci. Total. Environ. 2014, 487, 224–232. [Google Scholar] [CrossRef]
- Leppkes, M.; Neurath, M.F. Cytokines in inflammatory bowel diseases–update 2020. Pharmacol. Res. 2020, 158, 104835. [Google Scholar] [CrossRef] [PubMed]
- Tracey, D.; Klareskog, L.; Sasso, E.H.; Salfeld, J.G.; Tak, P.P. Tumor necrosis factor antagonist mechanisms of action: A comprehensive review. Pharm. Ther. 2008, 117, 244–279. [Google Scholar] [CrossRef]
- Blandizzi, C.; Gionchetti, P.; Armuzzi, A.; Caporali, R.; Chimenti, S.; Cimaz, R.; Cimino, L.; Lapadula, G.; Lionetti, P.; Marchesoni, A. The role of tumour necrosis factor in the pathogenesis of immune-mediated diseases. Int. J. Immunopathol. Pharmacol. 2014, 27, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Anders, H. Of inflammasomes and alarmins: IL-1β and IL-1α in kidney disease. J. Am. Soc. Nephrol. 2016, 27, 2564–2575. [Google Scholar] [CrossRef] [PubMed]
- Coccia, M.; Harrison, O.J.; Schiering, C.; Asquith, M.J.; Becher, B.; Powrie, F.; Maloy, K.J. IL-1β mediates chronic intestinal inflammation by promoting the accumulation of IL-17A secreting innate lymphoid cells and CD4+ Th17 cells. J. Exp. Med. 2012, 209, 1595–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanneganti, T.D. Inflammatory Bowel Disease and the NLRP3 Inflammasome. N. Engl. J. Med. 2017, 377, 694–696. [Google Scholar] [CrossRef] [PubMed]
- Tourkochristou, E.; Aggeletopoulou, I.; Konstantakis, C.; Triantos, C. Role of NLRP3 inflammasome in inflammatory bowel diseases. World J. Gastroenterol. 2019, 25, 4796. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Fukuyama, S.; Sakai-Tagawa, Y.; Takashita, E.; Shoemaker, J.E.; Kawaoka, Y. C646, a novel p300/CREB-binding protein-specific inhibitor of histone acetyltransferase, attenuates influenza A virus infection. Antimicrob. Agents Chemother. 2016, 60, 1902–1906. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Li, J.; Long, X.; Tao, S.; Yu, X.; Ruan, X.; Zhao, K.; Tian, L. C646 Protects Against DSS-Induced Colitis Model by Targeting NLRP3 Inflammasome. Front. Pharmacol. 2021, 12, 707610. [Google Scholar] [CrossRef]
- Long, X.; Yu, X.; Gong, P.; Wang, X.; Tian, L. Identification of WT161 as a Potent Agent for the Treatment of Colitis by Targeting the Nucleotide-Binding Domain-Like Receptor Family Pyrin Domain Containing 3 Inflammasome. Front. Pharmacol. 2022, 13, 780179. [Google Scholar] [CrossRef]
- Martinon, F.; Mayor, A.; Tschopp, J. The inflammasomes: Guardians of the body. Annu. Rev. Immunol. 2009, 27, 229–265. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef]
- Alex, P.; Zachos, N.C.; Nguyen, T.; Gonzales, L.; Chen, T.E.; Conklin, L.S.; Centola, M.; Li, X. Distinct cytokine pat-terns identified from multiplex profiles of murine DSS and TNBS-induced colitis. Inflamm. Bowel Dis. 2009, 15, 341–352. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Gong, P.; Long, X.; Jiang, Y.; Ye, M.; Tao, S.; Su, Y.; Yang, F.; Tian, L. Microcystin-LR Induces and Aggravates Colitis through NLRP3 Inflammasome-Mediated Pyroptosis in Mice. Toxins 2023, 15, 447. https://doi.org/10.3390/toxins15070447
Yang Y, Gong P, Long X, Jiang Y, Ye M, Tao S, Su Y, Yang F, Tian L. Microcystin-LR Induces and Aggravates Colitis through NLRP3 Inflammasome-Mediated Pyroptosis in Mice. Toxins. 2023; 15(7):447. https://doi.org/10.3390/toxins15070447
Chicago/Turabian StyleYang, Yue, Pan Gong, Xiuyan Long, Yuanjuan Jiang, Mingmei Ye, Sifan Tao, Yahui Su, Fei Yang, and Li Tian. 2023. "Microcystin-LR Induces and Aggravates Colitis through NLRP3 Inflammasome-Mediated Pyroptosis in Mice" Toxins 15, no. 7: 447. https://doi.org/10.3390/toxins15070447
APA StyleYang, Y., Gong, P., Long, X., Jiang, Y., Ye, M., Tao, S., Su, Y., Yang, F., & Tian, L. (2023). Microcystin-LR Induces and Aggravates Colitis through NLRP3 Inflammasome-Mediated Pyroptosis in Mice. Toxins, 15(7), 447. https://doi.org/10.3390/toxins15070447