Gut Microbiota Interventions to Retain Residual Kidney Function



Abstract
:1. Introduction
2. Residual Kidney Function in CKD
3. Gut Microbiota
4. Gut Microbiota Composition in CKD: Is Dysbiosis a Problem?
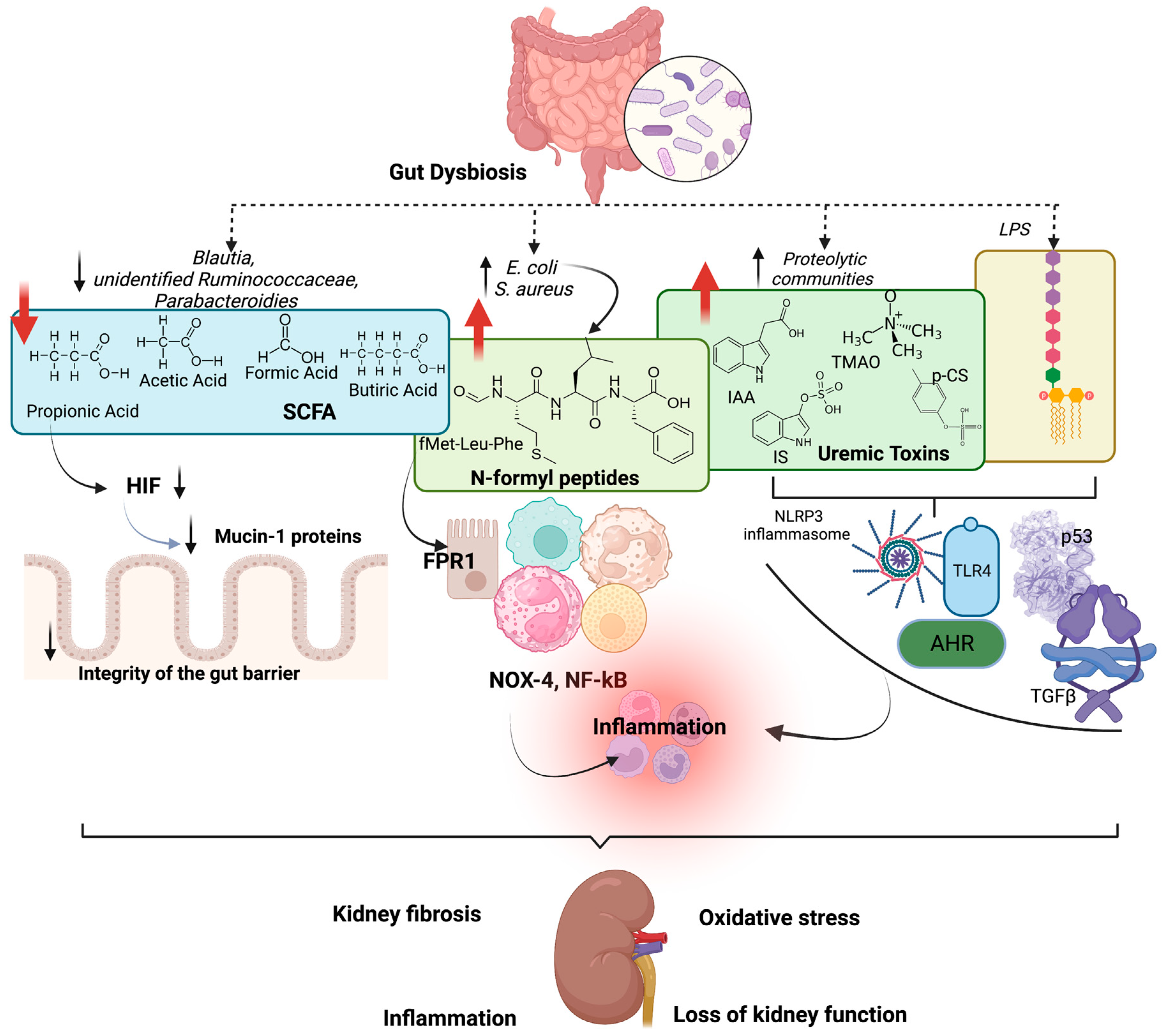
5. Effects of Gut Dysbiosis on Kidney Function
6. Gut Microbiota Modulation and Effects on Residual Kidney Function
6.1. Fibres
6.2. Probiotics
6.3. Synbiotics
6.4. Dietary Proteins
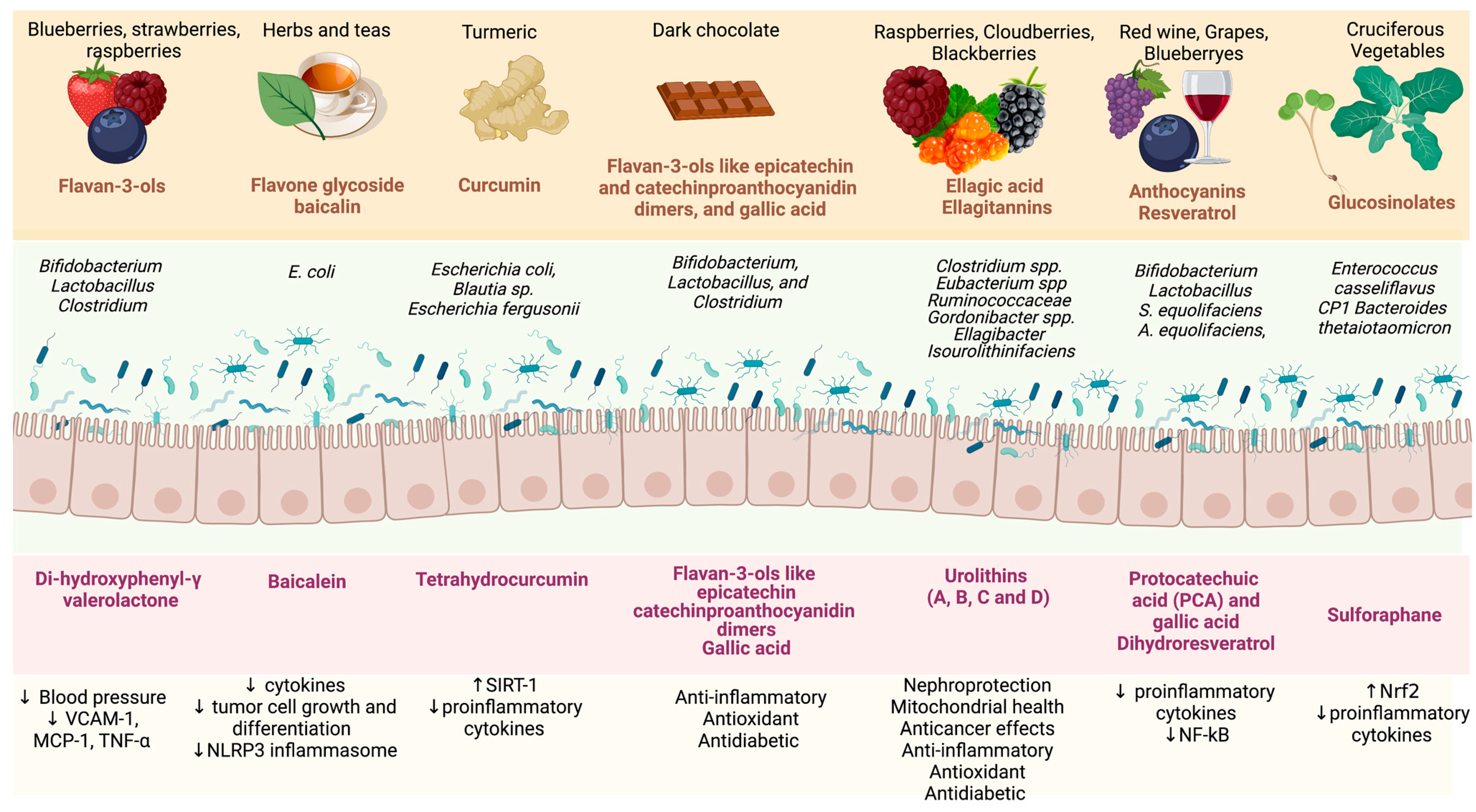
6.5. Bioactive Compounds
6.6. Faecal Microbiota Transplantation
6.7. Medications
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AhR | Aryl hydrocarbon receptor |
| AKI | Acute kidney injury |
| CKD | Chronic kidney disease |
| CVD | Cardiovascular disease |
| eGFR | Estimated glomerular filtration rate |
| fMLF | Fmet-Leu-Phe |
| fMIFL | Fmet-Ile-Phe-Leu |
| FMT | Faecal microbiota transplant |
| FOS | Fructooligosaccharides |
| FRP1 | Formyl peptide receptor 1 |
| HD | Haemodialysis |
| IS | Indoxyl sulfate |
| IL | Interleukin |
| IAA | Indol-3-acetic acid |
| LPS | Lipopolysaccharides |
| MCP-1 | Monocyte chemoattractant protein-1 |
| NOX-4 | NADPH-oxidase 4 |
| NF-κB | Nuclear factor-κB |
| NLRP3 | NLR family pyrin domain-containing 3 |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| PAGln | Phenylacetylglutamine |
| PAGly | Phenylacetylglycine |
| p-CS | P-cresyl sulfate |
| PS | Phenyl sulfate |
| PD | Peritoneal dialysis |
| RAAS | Renin–angiotensin–aldosterone system |
| SCFAs | Short-chain fatty acids |
| TMAO | Trimethylamine N-oxide |
| SGLT2 | Sodium-glucose cotransporter-2 |
| SIRT-1 | Sirtuin-1 |
| TMA | Trimethylamine |
| TLR-4 | Toll-like receptor-4 |
| TGF-β | Transforming growth factor beta |
| TNF-∝ | Tumour necrosis factor |
| TWEAK | TNF-like weak inducer of apoptosis |
| VCAM-1 | Vascular cell adhesion molecule-1 |
References
- Krukowski, H.; Valkenburg, S.; Madella, A.M.; Garssen, J.; van Bergenhenegouwen, J.; Overbeek, S.A.; Huys, G.R.B.; Raes, J.; Glorieux, G. Gut microbiome studies in CKD: Opportunities, pitfalls and therapeutic potential. Nat. Rev. Nephrol. 2023, 19, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Mafra, D.; Kalantar-Zadeh, K.; Moore, L.W. New Tricks for Old Friends: Treating Gut Microbiota of Patients with CKD. J. Ren. Nutr. 2021, 31, 433–437. [Google Scholar] [CrossRef]
- Graboski, A.L.; Redinbo, M.R. Gut-Derived Protein-Bound Uremic Toxins. Toxins 2020, 12, 590. [Google Scholar] [CrossRef]
- Jiang, N.; Zhang, C.; Feng, H.; Yuan, J.; Ding, L.; Fang, W.; Gu, A.; Huang, J.; Li, N.; Gu, L.; et al. Clinical Characteristics Associated with the Properties of Gut Microbiota in Peritoneal Dialysis Patients. Perit. Dial. Int. J. Int. Soc. Perit. Dial. 2021, 41, 298–306. [Google Scholar] [CrossRef]
- Cao, C.; Zhu, H.; Yao, Y.; Zeng, R. Gut Dysbiosis and Kidney Diseases. Front. Med. 2022, 9, 829349. [Google Scholar] [CrossRef]
- Rysz, J.; Franczyk, B.; Ławiński, J.; Olszewski, R.; Ciałkowska-Rysz, A.; Gluba-Brzózka, A. The Impact of CKD on Uremic Toxins and Gut Microbiota. Toxins 2021, 13, 252. [Google Scholar] [CrossRef] [PubMed]
- Wehedy, E.; Shatat, I.F.; Al Khodor, S. The Human Microbiome in Chronic Kidney Disease: A Double-Edged Sword. Front. Med. 2022, 8, 790783. [Google Scholar] [CrossRef]
- Tourountzis, T.; Lioulios, G.; Fylaktou, A.; Moysidou, E.; Papagianni, A.; Stangou, M. Microbiome in Chronic Kidney Disease. Life 2022, 12, 1513. [Google Scholar] [CrossRef]
- Arora, A.; Behl, T.; Sehgal, A.; Singh, S.; Sharma, N.; Bhatia, S.; Sobarzo-Sanchez, E.; Bungau, S. Unravelling the Involvement of Gut Microbiota in Type 2 Diabetes Mellitus. Life Sci. 2021, 273, 119311. [Google Scholar] [CrossRef] [PubMed]
- Mafra, D.; Borges, N.A.; Lindholm, B.; Shiels, P.G.; Evenepoel, P.; Stenvinkel, P. Food as Medicine: Targeting the Uraemic Phenotype in Chronic Kidney Disease. Nat. Rev. Nephrol. 2021, 17, 153–171. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.; Zhang, P.; Guo, S.; Zou, J.; Ji, J.; Ding, X.; Yu, X. Altered Gut Microbiota and Gut-Derived p-Cresyl Sulfate Serum Levels in Peritoneal Dialysis Patients. Front. Cell. Infect. Microbiol. 2022, 12, 639624. [Google Scholar] [CrossRef] [PubMed]
- Tanriover, C.; Ucku, D.; Basile, C.; Tuttle, K.R.; Kanbay, M. On the Importance of the Interplay of Residual Renal Function with Clinical Outcomes in End-Stage Kidney Disease. J. Nephrol. 2022, 35, 2191–2204. [Google Scholar] [CrossRef] [PubMed]
- Manns, B.; Hemmelgarn, B.; Lillie, E.; Dip, S.C.P.G.; Cyr, A.; Gladish, M.; Large, C.; Silverman, H.; Toth, B.; Wolfs, W.; et al. Setting Research Priorities for Patients on or Nearing Dialysis. Clin. J. Am. Soc. Nephrol. 2014, 9, 1813–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikizler, T.A.; Burrowes, J.D.; Byham-Gray, L.D.; Campbell, K.L.; Carrero, J.J.; Chan, W.; Fouque, D.; Friedman, A.N.; Ghaddar, S.; Goldstein-Fuchs, D.J.; et al. KDOQI Clinical Practice Guideline for Nutrition in CKD: 2020 Update. Am. J. Kidney Dis. 2020, 76, S1–S107. [Google Scholar] [CrossRef]
- Kalantar-Zadeh, K.; Joshi, S.; Schlueter, R.; Cooke, J.; Brown-Tortorici, A.; Donnelly, M.; Schulman, S.; Lau, W.L.; Rhee, C.M.; Streja, E.; et al. Plant-Dominant Low-Protein Diet for Conservative Management of Chronic Kidney Disease. Nutrients 2020, 12, 1931. [Google Scholar] [CrossRef]
- Joshi, S.; McMacken, M.; Kalantar-Zadeh, K. Plant-Based Diets for Kidney Disease: A Guide for Clinicians. Am. J. Kidney Dis. 2021, 77, 287–296. [Google Scholar] [CrossRef]
- Mathew, A.T.; Fishbane, S.; Obi, Y.; Kalantar-Zadeh, K. Preservation of Residual Kidney Function in Hemodialysis Patients: Reviving an Old Concept. Kidney Int. 2016, 90, 262–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecking, M.; McCullough, K.P.; Port, F.K.; Bieber, B.; Morgenstern, H.; Yamamoto, H.; Suri, R.S.; Jadoul, M.; Gesualdo, L.; Perl, J.; et al. Self-Reported Urine Volume in Hemodialysis Patients: Predictors and Mortality Outcomes in the International Dialysis Outcomes and Practice Patterns Study (DOPPS). Am. J. Kidney Dis. 2019, 74, 425–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nongnuch, A.; Assanatham, M.; Panorchan, K.; Davenport, A. Strategies for Preserving Residual Renal Function in Peritoneal Dialysis Patients. Clin. Kidney J. 2015, 8, 202–211. [Google Scholar] [CrossRef] [Green Version]
- Bargman, J.M.; Thorpe, K.E.; Churchill, D.N. Relative Contribution of Residual Renal Function and Peritoneal Clearance to Adequacy of Dialysis: A Reanalysis of the CANUSA Study. J. Am. Soc. Nephrol. 2001, 12, 2158–2162. [Google Scholar] [CrossRef]
- Te Liao, C.; Chen, Y.M.; Shiao, C.C.; Hu, F.C.; Huang, J.W.; Kao, T.W.; Chuang, H.F.; Hung, K.Y.; Wu, K.D.; Tsai, T.J. Rate of Decline of Residual Renal Function Is Associated with All-Cause Mortality and Technique Failure in Patients on Long-Term Peritoneal Dialysis. Nephrol. Dial. Transplant. 2009, 24, 2909–2914. [Google Scholar] [CrossRef] [Green Version]
- Jansen, M.A.M.; Hart, A.A.M.; Korevaar, J.C.; Dekker, F.W.; Boeschoten, E.W.; Krediet, R.T.; Birnie, R.J.; Boekhout, M.; Boer, W.H.; van Buren, M.; et al. Predictors of the Rate of Decline of Residual Renal Function in Incident Dialysis Patients. Kidney Int. 2002, 62, 1046–1053. [Google Scholar] [CrossRef] [Green Version]
- Canaud, B.; Kooman, J.P.; Selby, N.M.; Taal, M.W.; Francis, S.; Maierhofer, A.; Kopperschmidt, P.; Collins, A.; Kotanko, P. Dialysis-Induced Cardiovascular and Multiorgan Morbidity. Kidney Int. Rep. 2020, 5, 1856–1869. [Google Scholar] [CrossRef]
- Mccafferty, K.; Fan, S.; Davenport, A. Extracellular Volume Expansion, Measured by Multifrequency Bioimpedance, Does Not Help Preserve Residual Renal Function in Peritoneal Dialysis Patients. Kidney Int. 2014, 85, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, N.; Yan, Y.; Chen, N.; Xu, S.; Chu, R.; Wang, M.; Duan, S.; Ren, H.; Song, S.; Wang, L.; et al. Relationship between Gut Microbiota and Nutritional Status in Patients on Peritoneal Dialysis. Sci. Rep. 2023, 13, 1572. [Google Scholar] [CrossRef]
- Caria, S.; Cupisti, A.; Sau, G.; Bolasco, P. The Incremental Treatment of ESRD: A Low-Protein Diet Combined with Weekly Hemodialysis May Be Beneficial for Selected Patients. BMC Nephrol. 2014, 15, 172. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Qian, J.; Sun, W.; Lin, A.; Cao, L.; Wang, Q.; Ni, Z.; Wan, Y.; Linholm, B.; Axelsson, J.; et al. Better Preservation of Residual Renal Function in Peritoneal Dialysis Patients Treated with a Low-Protein Diet Supplemented with Keto Acids: A Prospective, Randomized Trial. Nephrol. Dial. Transplant. 2009, 24, 2551–2558. [Google Scholar] [CrossRef] [Green Version]
- Riccio, P.; Rossano, R. The Human Gut Microbiota Is Neither an Organ nor a Commensal. FEBS Lett. 2020, 594, 3262–3271. [Google Scholar] [CrossRef]
- Lazar, V.; Ditu, L.M.; Pircalabioru, G.G.; Picu, A.; Petcu, L.; Cucu, N.; Chifiriuc, M.C. Gut Microbiota, Host Organism, and Diet Trialogue in Diabetes and Obesity. Front. Nutr. 2019, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, G.J.; Domingues, S. We Are Never Alone: Living with the Human Microbiota. Front. Young Minds 2017, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voroneanu, L.; Burlacu, A.; Brinza, C.; Covic, A.; Balan, G.G.; Nistor, I.; Popa, C.; Hogas, S.; Covic, A. Gut Microbiota in Chronic Kidney Disease: From Composition to Modulation towards Better Outcomes—A Systematic Review. J. Clin. Med. 2023, 12, 1948. [Google Scholar] [CrossRef]
- Cresci, G.A.; Bawden, E. Gut Microbiome: What We Do and Don’t Know. Nutr. Clin. Pract. 2015, 30, 734–746. [Google Scholar] [CrossRef] [Green Version]
- Lazar, V.; Holban, A.M.; Curutiu, C.; Ditu, L.M. Modulation of Gut Microbiota by Essential Oils and Inorganic Nanoparticles: Impact in Nutrition and Health. Front. Nutr. 2022, 9, 920413. [Google Scholar] [CrossRef]
- Tiffany, C.R.; Bäumler, A.J. Dysbiosis: From Fiction to Function. Am. J. Physiol.-Gastrointest. Liver Physiol. 2019, 317, G602–G608. [Google Scholar] [CrossRef]
- Martinez, J.E.; Kahana, D.D.; Ghuman, S.; Wilson, H.P.; Wilson, J.; Kim, S.C.J.; Lagishetty, V.; Jacobs, J.P.; Sinha-Hikim, A.P.; Friedman, T.C. Unhealthy Lifestyle and Gut Dysbiosis: A Better Understanding of the Effects of Poor Diet and Nicotine on the Intestinal Microbiome. Front. Endocrinol. 2021, 12, 649. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The Healthy Human Microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Khodor, S.; Shatat, I.F. Gut Microbiome and Kidney Disease: A Bidirectional Relationship. Pediatr. Nephrol. 2017, 32, 921. [Google Scholar] [CrossRef] [Green Version]
- Simenhoff, M.L.; Saukkonen, J.J.; Burke, F.; Wesson, L.G.; Schaedler, R.W.; Gordon, S.J. Bacterial Populations of the Small Intestine in Uremia. Nephron 1978, 22, 63–68. [Google Scholar] [CrossRef]
- Hida, M.; Aiba, Y.; Sawamura, S.; Suzuki, N.; Satoh, T.; Koga, Y. Inhibition of the Accumulation of Uremic Toxins in the Blood and Their Precursors in the Feces after Oral Administration of Lebenin®, a Lactic Acid Bacteria Preparation, to Uremic Patients Undergoing Hemodialysis? Nephron 1996, 74, 349–355. [Google Scholar] [CrossRef]
- Wang, I.K.; Lai, H.C.; Yu, C.J.; Liang, C.C.; Chang, C.T.; Kuo, H.L.; Yang, Y.F.; Lin, C.C.; Lin, H.H.; Liu, Y.L.; et al. Real-Time PCR Analysis of the Intestinal Microbiotas in Peritoneal Dialysis Patients. Appl. Environ. Microbiol. 2012, 78, 1107–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaziri, N.D.; Wong, J.; Pahl, M.; Piceno, Y.M.; Yuan, J.; Desantis, T.Z.; Ni, Z.; Nguyen, T.H.; Andersen, G.L. Chronic Kidney Disease Alters Intestinal Microbial Flora. Kidney Int. 2013, 83, 308–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabatino, A.; Regolisti, G.; Brusasco, I.; Cabassi, A.; Morabito, S.; Fiaccadori, E. Alterations of Intestinal Barrier and Microbiota in Chronic Kidney Disease. Nephrol. Dial. Transplant. 2015, 30, 924–933. [Google Scholar] [CrossRef] [Green Version]
- Kanbay, M.; Onal, E.M.; Afsar, B.; Dagel, T.; Yerlikaya, A.; Covic, A.; Vaziri, N.D. The Crosstalk of Gut Microbiota and Chronic Kidney Disease: Role of Inflammation, Proteinuria, Hypertension, and Diabetes Mellitus. Int. Urol. Nephrol. 2018, 50, 1453–1466. [Google Scholar] [CrossRef] [Green Version]
- Bhargava, S.; Merckelbach, E.; Noels, H.; Vohra, A.; Jankowski, J. Homeostasis in the Gut Microbiota in Chronic Kidney Disease. Toxins 2022, 14, 648. [Google Scholar] [CrossRef]
- Evenepoel, P.; Poesen, R.; Meijers, B. The Gut-Kidney Axis. Pediatr. Nephrol. 2017, 32, 2005–2014. [Google Scholar] [CrossRef]
- Wong, J.; Piceno, Y.M.; DeSantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of Urease- and Uricase-Containing, Indole- and p-Cresol-Forming and Contraction of Short-Chain Fatty Acid-Producing Intestinal Microbiota in ESRD. Am. J. Nephrol. 2014, 39, 230–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lun, H.; Yang, W.; Zhao, S.; Jiang, M.; Xu, M.; Liu, F.; Wang, Y. Altered Gut Microbiota and Microbial Biomarkers Associated with Chronic Kidney Disease. Microbiologyopen 2019, 8, e00678. [Google Scholar] [CrossRef]
- Hu, Q.; Wu, K.; Pan, W.; Zeng, Y.; Hu, K.; Chen, D.; Huang, X.; Zhang, Q. Intestinal Flora Alterations in Patients with Early Chronic Kidney Disease: A Case-Control Study among the Han Population in Southwestern China. J. Int. Med. Res. 2020, 48, 1–12. [Google Scholar] [CrossRef]
- Li, F.X.; Wang, M.H.; Wang, J.P.; Li, R.S.; Zhang, Y.Q. Alterations to the Gut Microbiota and Their Correlation with Inflammatory Factors in Chronic Kidney Disease. Front. Cell. Infect. Microbiol. 2019, 9, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esgalhado, M.; Kemp, J.A.; Damasceno, N.R.T.; Fouque, D.; Mafra, D. Short-Chain Fatty Acids: A Link between Prebiotics and Microbiota in Chronic Kidney Disease. Future Microbiol. 2017, 12, 1413–1425. [Google Scholar] [PubMed]
- Magliocca, G.; Mone, P.; Di Iorio, B.R.; Heidland, A.; Marzocco, S. Short-Chain Fatty Acids in Chronic Kidney Disease: Focus on Inflammation and Oxidative Stress Regulation. Int. J. Mol. Sci. 2022, 23, 354. [Google Scholar] [CrossRef]
- Kim, J.E.; Kim, H.E.; Park, J.I.; Cho, H.; Kwak, M.J.; Kim, B.Y.; Yang, S.H.; Lee, J.P.; Kim, D.K.; Joo, K.W.; et al. The Association between Gut Microbiota and Uremia of Chronic Kidney Disease. Microorganisms 2020, 8, 907. [Google Scholar] [CrossRef]
- McIntyre, C.W.; Harrison, L.E.A.; Eldehni, M.T.; Jefferies, H.J.; Szeto, C.C.; John, S.G.; Sigrist, M.K.; Burton, J.O.; Hothi, D.; Korsheed, S.; et al. Circulating Endotoxemia: A Novel Factor in Systemic Inflammation and Cardiovascular Disease in Chronic Kidney Disease. Clin. J. Am. Soc. Nephrol. 2011, 6, 133. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.; Bosch, J.A.; Phillips, A.C.; Chin, S.H.; Antonysunil, A.; Inston, N.; Moore, S.; Kaur, O.; McTernan, P.G.; Borrows, R. The Associations of Endotoxemia With Systemic Inflammation, Endothelial Activation, and Cardiovascular Outcome in Kidney Transplantation. J. Ren. Nutr. 2018, 28, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Lim, Y.J.; Sidor, N.A.; Tonial, N.C.; Che, A.; Urquhart, B.L. Uremic Toxins in the Progression of Chronic Kidney Disease and Cardiovascular Disease: Mechanisms and Therapeutic Targets. Toxins 2021, 13, 142. [Google Scholar] [CrossRef]
- Jankowski, J.; Floege, J.; Fliser, D.; Böhm, M.; Marx, N. Cardiovascular Disease in Chronic Kidney Disease. Circulation 2021, 143, 1157–1172. [Google Scholar] [CrossRef]
- Lin, T.Y.; Wu, P.H.; Lin, Y.T.; Hung, S.C. Gut Dysbiosis and Mortality in Hemodialysis Patients. npj Biofilms Microbiomes 2021, 7, 20. [Google Scholar] [CrossRef]
- Saranya, G.R.; Viswanathan, P. Gut Microbiota Dysbiosis in AKI to CKD Transition. Biomed. Pharmacother. 2023, 161, 114447. [Google Scholar] [CrossRef]
- Chen, G.; Wang, X.; Liao, Q.; Ge, Y.; Jiao, H.; Chen, Q.; Liu, Y.; Lyu, W.; Zhu, L.; van Zundert, G.C.P.; et al. Structural Basis for Recognition of N-Formyl Peptides as Pathogen-Associated Molecular Patterns. Nat. Commun. 2022, 13, 5232. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.S.; Bae, Y.S. Formyl Peptide Receptors in the Mucosal Immune System. Exp. Mol. Med. 2020, 52, 1694–1704. [Google Scholar] [CrossRef] [PubMed]
- Salamah, M.F.; Ravishankar, D.; Vaiyapuri, R.; Moraes, L.A.; Patel, K.; Perretti, M.; Gibbins, J.M.; Vaiyapuri, S. The Formyl Peptide FMLF Primes Platelet Activation and Augments Thrombus Formation. J. Thromb. Haemost. 2019, 17, 1120–1133. [Google Scholar] [CrossRef] [Green Version]
- Anders, H.J.; Andersen, K.; Stecher, B. The Intestinal Microbiota, a Leaky Gut, and Abnormal Immunity in Kidney Disease. Kidney Int. 2013, 83, 1010–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briskey, D.; Tucker, P.; Johnson, D.W.; Coombes, J.S. The Role of the Gastrointestinal Tract and Microbiota on Uremic Toxins and Chronic Kidney Disease Development. Clin. Exp. Nephrol. 2017, 21, 7–15. [Google Scholar] [CrossRef]
- Glorieux, G.; Nigam, S.K.; Vanholder, R.; Verbeke, F. Role of the Microbiome in Gut-Heart-Kidney Cross Talk. Circ. Res. 2023, 132, 1064–1083. [Google Scholar] [CrossRef]
- Brito, J.S.; Borges, N.A.; Esgalhado, M.; Magliano, D.C.; Soulage, C.O.; Mafra, D. Aryl Hydrocarbon Receptor Activation in Chronic Kidney Disease: Role of Uremic Toxins. Nephron. 2017, 137, 1–7. [Google Scholar] [CrossRef]
- Shimizu, H.; Yisireyili, M.; Nishijima, F.; Niwa, T. Indoxyl Sulfate Enhances P53-TGF-Β1-Smad3 Pathway in Proximal Tubular Cells. Am. J. Nephrol. 2013, 37, 97–103. [Google Scholar] [CrossRef]
- Ichii, O.; Otsuka-Kanazawa, S.; Nakamura, T.; Ueno, M.; Kon, Y.; Chen, W.; Rosenberg, A.Z.; Kopp, J.B. Podocyte Injury Caused by Indoxyl Sulfate, a Uremic Toxin and Aryl-Hydrocarbon Receptor Ligand. PLoS ONE 2014, 9, e108448. [Google Scholar] [CrossRef]
- Balint, L.; Socaciu, C.; Socaciu, A.I.; Vlad, A.; Gadalean, F.; Bob, F.; Milas, O.; Cretu, O.M.; Suteanu-Simulescu, A.; Glavan, M.; et al. Metabolite Profiling of the Gut-Renal-Cerebral Axis Reveals a Particular Pattern in Early Diabetic Kidney Disease in T2DM Patients. Int. J. Mol. Sci. 2023, 24, 6212. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Sanchez-Niño, M.D.; Glorieux, G.; Sanz, A.B.; Egido, J.; Vanholder, R.; Ortiz, A. P-Cresyl Sulphate Has pro-Inflammatory and Cytotoxic Actions on Human Proximal Tubular Epithelial Cells. Nephrol. Dial. Transplant. 2014, 29, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, L.; Sallée, M.; Cerini, C.; Poitevin, S.; Gondouin, B.; Jourde-Chiche, N.; Fallague, K.; Brunet, P.; Calaf, R.; Dussol, B.; et al. The Cardiovascular Effect of the Uremic Solute Indole-3 Acetic Acid. J. Am. Soc. Nephrol. 2015, 26, 876–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Wang, X.; Liu, X.; Wang, L.; Ren, F.; Wang, X.; Leng, X. Hippuric Acid Promotes Renal Fibrosis by Disrupting Redox Homeostasis via Facilitation of NRF2-KEAP1-CUL3 Interactions in Chronic Kidney Disease. Antioxidants 2020, 9, 783. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Zheng, B.; Liu, N.; Liu, J.; Liu, W.; Huang, X.; Zeng, X.; Chen, L.; Li, Z.; Ouyang, D. Trimethylamine N-Oxide Exacerbates Renal Inflammation and Fibrosis in Rats with Diabetic Kidney Disease. Front. Physiol. 2021, 12, 682482. [Google Scholar] [CrossRef]
- Sun, C.Y.; Cheng, M.L.; Pan, H.C.; Lee, J.H.; Lee, C.C. Protein-Bound Uremic Toxins Impaired Mitochondrial Dynamics and Functions. Oncotarget 2017, 8, 77722. [Google Scholar] [CrossRef] [Green Version]
- Mafra, D.; Borges, N.A.; Lindholm, B.; Stenvinkel, P. Mitochondrial Dysfunction and Gut Microbiota Imbalance: An Intriguing Relationship in Chronic Kidney Disease. Mitochondrion 2019, 47, 206–209. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, X.; Chen, J.; Hu, Y.; Bai, Y.; Xu, W.; He, L.; Wang, Y.; Chen, C.; Chen, J. Evaluation of the Contribution of Gut Microbiome Dysbiosis to Cardiac Surgery-Associated Acute Kidney Injury by Comparative Metagenome Analysis. Front. Microbiol. 2023, 14, 1119959. [Google Scholar] [CrossRef]
- Gryp, T.; Huys, G.R.B.; Joossens, M.; Van Biesen, W.; Glorieux, G.; Vaneechoutte, M. Isolation and Quantification of Uremic Toxin Precursor-Generating Gut Bacteria in Chronic Kidney Disease Patients. Int. J. Mol. Sci. 2020, 21, 1986. [Google Scholar] [CrossRef] [Green Version]
- Ranganathan, N.; Anteyi, E. The Role of Dietary Fiber and Gut Microbiome Modulation in Progression of Chronic Kidney Disease. Toxins 2022, 14, 183. [Google Scholar] [CrossRef]
- Cigarrán Guldris, S.; Latorre Catalá, J.A.; Sanjurjo Amado, A.; Menéndez Granados, N.; Piñeiro Varela, E. Fibre Intake in Chronic Kidney Disease: What Fibre Should We Recommend? Nutrients 2022, 14, 4419. [Google Scholar] [CrossRef]
- Mazidi, M.; Katsiki, N.; Banach, M. Higher Plasma Levels of Valerate Produced by Gut Microbiota May Have a Beneficial Impact on Renal Function. J. Am. Nutr. Assoc. 2023, 42, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Huang, Y.F.; Wang, M.Q.; Chen, D.X.; Wan, H.; Wei, L.B.; Xiao, W. Dietary Fiber Intake Is Associated with Chronic Kidney Disease (CKD) Progression and Cardiovascular Risk, but Not Protein Nutritional Status, in Adults with CKD. Asia Pac. J. Clin. Nutr. 2017, 26, 598–605. [Google Scholar] [CrossRef]
- Ebrahim, Z.; Proost, S.; Tito, R.Y.; Raes, J.; Glorieux, G.; Moosa, M.R.; Blaauw, R. The Effect of SS-Glucan Prebiotic on Kidney Function, Uremic Toxins and Gut Microbiome in Stage 3 to 5 Chronic Kidney Disease (CKD) Predialysis Participants: A Randomized Controlled Trial. Nutrients 2022, 14, 805. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Dong, X.; Li, X.; Jia, R.; Zhang, H.L. Benefits of Resistant Starch Type 2 for Patients with End-Stage Renal Disease under Maintenance Hemodialysis: A Systematic Review and Meta-Analysis. Int. J. Med. Sci. 2021, 18, 811. [Google Scholar] [CrossRef] [PubMed]
- Younes, H.; Rémésy, C.; Behr, S.; Demigné, C. Fermentable Carbohydrate Exerts a Urea-Lowering Effect in Normal and Nephrectomized Rats. Am. J. Physiol. 1997, 272, G515–G521. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Qin, X.; Yang, C.; Sabatino, A.; Kelly, J.T.; Avesani, C.M.; Carrero, J.J. Fiber Intake and Health in People with Chronic Kidney Disease. Clin. Kidney J. 2021, 15, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Halloran, K.; Underwood, M.A. Probiotic Mechanisms of Action. Early Hum. Dev. 2019, 135, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Miraghajani, M.; Zaghian, N.; Dehkohneh, A.; Mirlohi, M.; Ghiasvand, R. Probiotic Soy Milk Consumption and Renal Function Among Type 2 Diabetic Patients with Nephropathy: A Randomized Controlled Clinical Trial. Probiotics Antimicrob. Proteins 2019, 11, 124–132. [Google Scholar] [CrossRef] [PubMed]
- de Faria Barros, A.; Borges, N.A.; Nakao, L.S.; Dolenga, C.J.; do Carmo, F.L.; de Carvalho Ferreira, D.; Stenvinkel, P.; Bergman, P.; Lindholm, B.; Mafra, D. Effects of Probiotic Supplementation on Inflammatory Biomarkers and Uremic Toxins in Non-Dialysis Chronic Kidney Patients: A Double-Blind, Randomized, Placebo-Controlled Trial. J. Funct. Foods 2018, 46, 378–383. [Google Scholar] [CrossRef]
- Borges, N.A.; Carmo, F.L.; Stockler-Pinto, M.B.; de Brito, J.S.; Dolenga, C.J.; Ferreira, D.C.; Nakao, L.S.; Rosado, A.; Fouque, D.; Mafra, D. Probiotic Supplementation in Chronic Kidney Disease: A Double-Blind, Randomized, Placebo-Controlled Trial. J. Ren. Nutr. 2018, 28, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.-T.; Zhou, J.; Zhu, J.-H.; Wu, C.-Y.; Shen, H.; Zhang, W.; Zhou, S.-S.; Xu, J.-D.; Mao, Q.; Zhang, Y.-Q.; et al. Gut Microbiota Mediates the Protective Effects of Traditional Chinese Medicine Formula Qiong-Yu-Gao against Cisplatin-Induced Acute Kidney Injury. Microbiol. Spectr. 2022, 10, e0075922. [Google Scholar] [CrossRef] [PubMed]
- Mafra, D.; Borges, N.A.; Alvarenga, L.; Ribeiro, M.; Fonseca, L.; Leal, V.O.; Shiels, P.G.; Stenvinkel, P. Fermented Food: Should Patients with Cardiometabolic Diseases Go Back to an Early Neolithic Diet? Crit. Rev. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef]
- Xi, Y.; Huang, Y.; Li, Y.; Yan, J.; Shi, Z. Fermented Feed Supplement Relieves Caecal Microbiota Dysbiosis and Kidney Injury Caused by High-Protein Diet in the Development of Gosling Gout. Animals 2020, 10, 2139. [Google Scholar] [CrossRef]
- He, L.X.; Abdolmaleky, H.M.; Yin, S.; Wang, Y.; Zhou, J.R. Dietary Fermented Soy Extract and Oligo-Lactic Acid Alleviate Chronic Kidney Disease in Mice via Inhibition of Inflammation and Modulation of Gut Microbiota. Nutrients 2020, 12, 2376. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.C.; Kok, K.E.; Huang, K.F.; Weng, Y.L.; Chung, Y.C. Effects of Fermented Red Bean Extract on Nephropathy in Streptozocin-Induced Diabetic Rats. Food Nutr. Res. 2020, 64, 4272. [Google Scholar] [CrossRef]
- Tao, S.; Tao, S.; Cheng, Y.; Liu, J.; Ma, L.; Fu, P. Effects of Probiotic Supplements on the Progression of Chronic Kidney Disease: A Meta-Analysis. Nephrology 2019, 24, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Lempert, K.D. Probiotics and CKD Progression: Are Creatinine-Based Estimates of GFR Applicable? Am. J. Kidney Dis. 2019, 74, 429–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mafra, D.; Borges, N.; Alvarenga, L.; Esgalhado, M.; Cardozo, L.; Lindholm, B.; Stenvinkel, P. Dietary Components That May Influence the Disturbed Gut Microbiota in Chronic Kidney Disease. Nutrients 2019, 11, 496. [Google Scholar] [CrossRef] [Green Version]
- Zaramela, L.S.; Martino, C.; Alisson-Silva, F.; Rees, S.D.; Diaz, S.L.; Chuzel, L.; Ganatra, M.B.; Taron, C.H.; Secrest, P.; Zuñiga, C.; et al. Gut Bacteria Responding to Dietary Change Encode Sialidases That Exhibit Preference for Red Meat-Associated Carbohydrates. Nat. Microbiol. 2019, 4, 2082–2089. [Google Scholar] [CrossRef]
- Tian, N.; Li, L.; Ng, J.K.C.; Li, P.K.T. The Potential Benefits and Controversies of Probiotics Use in Patients at Different Stages of Chronic Kidney Disease. Nutrients 2022, 14, 4044. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef]
- Nakabayashi, I.; Nakamura, M.; Kawakami, K.; Ohta, T.; Kato, I.; Uchida, K.; Yoshida, M. Effects of Synbiotic Treatment on Serum Level of P-Cresol in Haemodialysis Patients: A Preliminary Study. Nephrol. Dial. Transplant. 2011, 26, 1094–1098. [Google Scholar] [CrossRef] [Green Version]
- Guida, B.; Germanò, R.; Trio, R.; Russo, D.; Memoli, B.; Grumetto, L.; Barbato, F.; Cataldi, M. Effect of Short-Term Synbiotic Treatment on Plasma p-Cresol Levels in Patients with Chronic Renal Failure: A Randomized Clinical Trial. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Johnson, D.W.; Morrison, M.; Pascoe, E.M.; Coombes, J.S.; Forbes, J.M.; Szeto, C.C.; McWhinney, B.C.; Ungerer, J.P.J.; Campbell, K.L. Synbiotics Easing Renal Failure by Improving Gut Microbiology (SYNERGY): A Randomized Trial. Clin. J. Am. Soc. Nephrol. 2016, 11, 223–231. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, C.; Krishnasamy, R.; Stanton, T.; Savill, E.; Snelson, M.; Mihala, G.; Kelly, J.T.; Morrison, M.; Johnson, D.W.; Campbell, K.L. Synbiotics Easing Renal Failure by Improving Gut Microbiology Ii (Synergy Ii): A Feasibility Randomized Controlled Trial. Nutrients 2021, 13, 4481. [Google Scholar] [CrossRef]
- Cosola, C.; Rocchetti, M.T.; Di Bari, I.; Acquaviva, P.M.; Maranzano, V.; Corciulo, S.; Di Ciaula, A.; Di Palo, D.M.; La Forgia, F.M.; Fontana, S.; et al. An Innovative Synbiotic Formulation Decreases Free Serum Indoxyl Sulfate, Small Intestine Permeability and Ameliorates Gastrointestinal Symptoms in a Randomized Pilot Trial in Stage IIIb-IV CKD Patients. Toxins 2021, 13, 334. [Google Scholar] [CrossRef]
- Lydia, A.; Indra, T.A.; Rizka, A.; Abdullah, M. The Effects of Synbiotics on Indoxyl Sulphate Level, Constipation, and Quality of Life Associated with Constipation in Chronic Haemodialysis Patients: A Randomized Controlled Trial. BMC Nephrol. 2022, 23, 259. [Google Scholar] [CrossRef]
- Nguyen, T.T.U.; Kim, H.W.; Kim, W. Effects of Probiotics, Prebiotics, and Synbiotics on Uremic Toxins, Inflammation, and Oxidative Stress in Hemodialysis Patients: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Clin. Med. 2021, 10, 4456. [Google Scholar] [CrossRef]
- Massy, Z.A.; Drueke, T.B. Diet-Microbiota Interaction and Kidney Disease Progression. Kidney Int. 2021, 99, 797–800. [Google Scholar] [CrossRef]
- Lauriola, M.; Farré, R.; Evenepoel, P.; Overbeek, S.A.; Meijers, B. Food-Derived Uremic Toxins in Chronic Kidney Disease. Toxins 2023, 15, 116. [Google Scholar] [CrossRef]
- Wu, S.; Bhat, Z.F.; Gounder, R.S.; Ahmed, I.A.M.; Al-Juhaimi, F.Y.; Ding, Y.; Bekhit, A.E.D.A. Effect of Dietary Protein and Processing on Gut Microbiota-A Systematic Review. Nutrients 2022, 14, 453. [Google Scholar] [CrossRef]
- Lobel, L.; Cao, Y.G.; Fenn, K.; Glickman, J.N.; Garrett, W.S. Diet Posttranslationally Modifies the Mouse Gut Microbial Proteome to Modulate Renal Function. Science 2020, 369, 1518–1524. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Fu, M.; Chen, X.; Guo, J.; Chen, B.; Tao, X. Dietary Methionine Restriction Attenuates Renal Ischaemia/Reperfusion-Induced Myocardial Injury by Activating the CSE/H2S/ERS Pathway in Diabetic Mice. J. Cell. Mol. Med. 2020, 24, 9890–9897. [Google Scholar] [CrossRef]
- Osterholt, T.; Gloistein, C.; Todorova, P.; Becker, I.; Arenskrieger, K.; Melka, R.; Koehler, F.C.; Faust, M.; Wahlers, T.; Benzing, T.; et al. Preoperative Short-Term Restriction of Sulfur-Containing Amino Acid Intake for Prevention of Acute Kidney Injury After Cardiac Surgery: A Randomized, Controlled, Double-Blind, Translational Trial. J. Am. Heart Assoc. 2022, 11, e025229. [Google Scholar] [CrossRef] [PubMed]
- Delgadillo-Puga, C.; Sánchez-Castillo, D.R.; Cariño-Cervantes, Y.Y.; Torre-Villalvazo, I.; Tovar-Palacio, C.; Vásquez-Reyes, S.; Furuzawa-Carballeda, J.; Acevedo-Carabantes, J.A.; Camacho-Corona, M.d.R.; Guzmán-Mar, J.L.; et al. Vachellia Farnesiana Pods or a Polyphenolic Extract Derived from Them Exert Immunomodulatory, Metabolic, Renoprotective, and Prebiotic Effects in Mice Fed a High-Fat Diet. Int. J. Mol. Sci. 2023, 24, 7984. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ma, Y.; Xu, Q.; Shikov, A.N.; Pozharitskaya, O.N.; Flisyuk, E.V.; Liu, M.; Li, H.; Vargas-Murga, L.; Duez, P. Flavonoids and Saponins: What Have We Got or Missed? Phytomedicine 2023, 109, 154580. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Miao, B.; Cao, H.; Tian, X.; Shen, L.; Yang, Z.; Yuan, F.; Ding, Y. Monkfish (Lophius litulon) Peptides Ameliorate High-Fat-Diet-Induced Nephrotoxicity by Reducing Oxidative Stress and Inflammation via Regulation of Intestinal Flora. Molecules 2022, 28, 245. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.; Alvarenga, L.; Cardozo, L.F.M.F.; Chermut, T.R.; Sequeira, J.; de Souza Gouveia Moreira, L.; Teixeira, K.T.R.; Shiels, P.G.; Stenvinkel, P.; Mafra, D. From the Distinctive Smell to Therapeutic Effects: Garlic for Cardiovascular, Hepatic, Gut, Diabetes and Chronic Kidney Disease. Clin. Nutr. 2021, 40, 4807–4819. [Google Scholar] [CrossRef] [PubMed]
- Verediano, T.A.; Stampini Duarte Martino, H.; Dias Paes, M.C.; Tako, E. Effects of Anthocyanin on Intestinal Health: A Systematic Review. Nutrients 2021, 13, 1331. [Google Scholar] [CrossRef]
- Coutinho-Wolino, K.S.; Melo, M.F.S.; Mota, J.C.; Mafra, D.; Guimarães, J.T.; Stockler-Pinto, M.B. Blueberry, Cranberry, Raspberry, and Strawberry as Modulators of the Gut Microbiota: Target for Treatment of Gut Dysbiosis in Chronic Kidney Disease? From Current Evidence to Future Possibilities. Nutr. Rev. 2023. [Google Scholar] [CrossRef]
- Ma, M.; Wang, Y.; Fan, S.; Huang, Y.; Su, X.; Lu, C. Urolithin A Alleviates Colitis in Mice by Improving Gut Microbiota Dysbiosis, Modulating Microbial Tryptophan Metabolism, and Triggering AhR Activation. J. Agric. Food Chem. 2023, 71, 7710–7722. [Google Scholar] [CrossRef]
- Moreira, L.D.S.G.; Brum Da Costa, I.D.S.; De Vargas Reis, D.C.M.; Trugilho, L.; Chermut, T.R.; Esgalhado, M.; Cardozo, L.F.M.F.; Stenvinkel, P.; Shiels, P.G.; Mafra, D. Cinnamon: An Aromatic Condiment Applicable to Chronic Kidney Disease. Kidney Res. Clin. Pract. 2023, 42, 4–26. [Google Scholar] [CrossRef]
- Moreira, L.D.S.G.; Fanton, S.; Cardozo, L.; Borges, N.A.; Combet, E.; Shiels, P.G.; Stenvinkel, P.; Mafra, D. Pink Pressure: Beetroot (Beta Vulgaris Rubra) as a Possible Novel Medical Therapy for Chronic Kidney Disease. Nutr. Rev. 2022, 80, 1041–1061. [Google Scholar] [CrossRef] [PubMed]
- Chermut, T.R.; Fonseca, L.; Figueiredo, N.; de Oliveira Leal, V.; Borges, N.A.; Cardozo, L.F.M.F.; Correa Leite, P.E.; Alvarenga, L.; Regis, B.; Delgado, A.; et al. Effects of Propolis on Inflammation Markers in Patients Undergoing Hemodialysis: A Randomized, Double-Blind Controlled Clinical Trial. Complement. Ther. Clin. Pract. 2023, 51, 101732. [Google Scholar] [CrossRef] [PubMed]
- Baptista, B.G.; Fanton, S.; Ribeiro, M.; Cardozo, L.F.; Regis, B.; Alvarenga, L.; Ribeiro-Alves, M.; Berretta, A.A.; Shiels, P.G.; Mafra, D. The Effect of Brazilian Green Propolis Extract on Inflammation in Patients with Chronic Kidney Disease on Peritoneal Dialysis: A Randomised Double-Blind Controlled Clinical Trial. Phytomedicine 2023, 114, 154731. [Google Scholar] [CrossRef]
- Alvarenga, L.; Cardozo, L.F.M.F.; Borges, N.A.; Chermut, T.R.; Ribeiro, M.; Leite, M.; Shiels, P.G.; Stenvinkel, P.; Mafra, D. To Bee or Not to Bee? The Bee Extract Propolis as a Bioactive Compound in the Burden of Lifestyle Diseases. Nutrition 2021, 83, 111094. [Google Scholar] [CrossRef] [PubMed]
- Salarolli, R.T.; Alvarenga, L.; Cardozo, L.F.M.F.; Teixeira, K.T.R.; Moreira, L.d.S.G.; Lima, J.D.; Rodrigues, S.D.; Nakao, L.S.; Fouque, D.; Mafra, D. Can Curcumin Supplementation Reduce Plasma Levels of Gut-Derived Uremic Toxins in Hemodialysis Patients? A Pilot Randomized, Double-Blind, Controlled Study. Int. Urol. Nephrol. 2021, 53, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Kwon, C.; Ediriweera, M.K.; Kim Cho, S. Interplay between Phytochemicals and the Colonic Microbiota. Nutrients 2023, 15, 1989. [Google Scholar] [CrossRef] [PubMed]
- Mithul Aravind, S.; Wichienchot, S.; Tsao, R.; Ramakrishnan, S.; Chakkaravarthi, S. Role of Dietary Polyphenols on Gut Microbiota, Their Metabolites and Health Benefits. Food Res. Int. 2021, 142, 110189. [Google Scholar] [CrossRef] [PubMed]
- Borody, T. Fecal Microbiota Transplantation: Treatment of the Gut Microbiome. In Gastrointestinal Diseases and Their Associated Infections; Eslick, G.D., Ed.; Elsevier: St Louis, MO, USA, 2019; pp. 249–261. [Google Scholar]
- Varga, A.; Kocsis, B.; Sipos, D.; Kása, P.; Vigvári, S.; Pál, S.; Dembrovszky, F.; Farkas, K.; Péterfi, Z. How to Apply FMT More Effectively, Conveniently and Flexible—A Comparison of FMT Methods. Front. Cell. Infect. Microbiol. 2021, 11, 657320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Luo, W.; Shi, Y.; Fan, Z.; Ji, G. Should We Standardize the 1,700-Year-Old Fecal Microbiota Transplantation? Am. J. Gastroenterol. 2012, 107, 1755. [Google Scholar] [CrossRef]
- Caggiano, G.; Stasi, A.; Franzin, R.; Fiorentino, M.; Cimmarusti, M.T.; Deleonardis, A.; Palieri, R.; Pontrelli, P.; Gesualdo, L. Fecal Microbiota Transplantation in Reducing Uremic Toxins Accumulation in Kidney Disease: Current Understanding and Future Perspectives. Toxins 2023, 15, 115. [Google Scholar] [CrossRef] [PubMed]
- Bian, J.; Liebert, A.; Bicknell, B.; Chen, X.M.; Huang, C.; Pollock, C.A. Faecal Microbiota Transplantation and Chronic Kidney Disease. Nutrients 2022, 14, 2528. [Google Scholar] [CrossRef]
- Wang, X.; Yang, S.; Li, S.; Zhao, L.; Hao, Y.; Qin, J.; Zhang, L.; Zhang, C.; Bian, W.; Zuo, L.; et al. Original Research: Aberrant Gut Microbiota Alters Host Metabolome and Impacts Renal Failure in Humans and Rodents. Gut 2020, 69, 2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Su, X.; Gao, Y.; Lv, C.; Gao, Z.; Liu, Y.; Wang, Y.; Li, S.; Wang, Z. The Potential Role of the Gut Microbiota in Modulating Renal Function in Experimental Diabetic Nephropathy Murine Models Established in Same Environment. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165764. [Google Scholar] [CrossRef] [PubMed]
- Barba, C.; Soulage, C.O.; Caggiano, G.; Glorieux, G.; Fouque, D.; Koppe, L. Effects of Fecal Microbiota Transplantation on Composition in Mice with CKD. Toxins 2020, 12, 741. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.B.; Lu, J.; Chen, P.P.; Lu, C.C.; Zhang, J.X.; Li, X.Q.; Yuan, B.Y.; Huang, S.J.; Ruan, X.Z.; Liu, B.C.; et al. Dysbiosis of Intestinal Microbiota Mediates Tubulointerstitial Injury in Diabetic Nephropathy via the Disruption of Cholesterol Homeostasis. Theranostics 2020, 10, 2803. [Google Scholar] [CrossRef]
- Bastos, R.M.C.; Simplício-Filho, A.; Sávio-Silva, C.; Oliveira, L.F.V.; Cruz, G.N.F.; Sousa, E.H.; Noronha, I.L.; Mangueira, C.L.P.; Quaglierini-Ribeiro, H.; Josefi-Rocha, G.R.; et al. Fecal Microbiota Transplant in a Pre-Clinical Model of Type 2 Diabetes Mellitus, Obesity and Diabetic Kidney Disease. Int. J. Mol. Sci. 2022, 23, 3842. [Google Scholar] [CrossRef]
- Zhou, G.; Zeng, J.; Peng, L.; Wang, L.; Zheng, W.; Wu, D.; Yang, Y. Fecal Microbiota Transplantation for Membranous Nephropathy. CEN Case Rep. 2021, 10, 261. [Google Scholar] [CrossRef]
- Zhao, J.; Bai, M.; Yang, X.; Wang, Y.; Li, R.; Sun, S. Alleviation of Refractory IgA Nephropathy by Intensive Fecal Microbiota Transplantation: The First Case Reports. Ren. Fail. 2021, 43, 928. [Google Scholar] [CrossRef]
- Weersma, R.K.; Zhernakova, A.; Fu, J. Interaction between Drugs and the Gut Microbiome. Gut 2020, 69, 1510–1519. [Google Scholar] [CrossRef]
- Ramirez, J.; Guarner, F.; Bustos Fernandez, L.; Maruy, A.; Sdepanian, V.L.; Cohen, H. Antibiotics as Major Disruptors of Gut Microbiota. Front. Cell. Infect. Microbiol. 2020, 10, 572912. [Google Scholar] [CrossRef] [PubMed]
- Cuisiniere, T.; Calvé, A.; Fragoso, G.; Oliero, M.; Hajjar, R.; Gonzalez, E.; Santos, M.M. Oral Iron Supplementation after Antibiotic Exposure Induces a Deleterious Recovery of the Gut Microbiota. BMC Microbiol. 2021, 21, 259. [Google Scholar] [CrossRef]
- Ribeiro, M.; Fonseca, L.; Anjos, J.S.; Capo-chichi, J.C.C.; Borges, N.A.; Burrowes, J.; Mafra, D. Oral Iron Supplementation in Patients with Chronic Kidney Disease: Can It Be Harmful to the Gut Microbiota? Nutr. Clin. Pract. 2022, 37, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Imhann, F.; Bonder, M.J.; Vila, A.V.; Fu, J.; Mujagic, Z.; Vork, L.; Tigchelaar, E.F.; Jankipersadsing, S.A.; Cenit, M.C.; Harmsen, H.J.M.; et al. Proton Pump Inhibitors Affect the Gut Microbiome. Gut 2016, 65, 740–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiraga, Y.; Kubota, T.; Katoh, M.; Horai, Y.; Suzuki, H.; Yamashita, Y.; Hirata, R.; Moroi, M. AST-120 Treatment Alters the Gut Microbiota Composition and Suppresses Hepatic Triglyceride Levels in Obese Mice. Endocr. Res. 2021, 46, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Yoshifuji, A.; Wakino, S.; Irie, J.; Matsui, A.; Hasegawa, K.; Tokuyama, H.; Hayashi, K.; Itoh, H. Oral Adsorbent AST-120 Ameliorates Gut Environment and Protects against the Progression of Renal Impairment in CKD Rats. Clin. Exp. Nephrol. 2018, 22, 1069. [Google Scholar] [CrossRef] [Green Version]
- Sato, E.; Hosomi, K.; Sekimoto, A.; Mishima, E.; Oe, Y.; Saigusa, D.; Ito, S.; Abe, T.; Sato, H.; Kunisawa, J.; et al. Effects of the Oral Adsorbent AST-120 on Fecal p-Cresol and Indole Levels and on the Gut Microbiota Composition. Biochem. Biophys. Res. Commun. 2020, 525, 773–779. [Google Scholar] [CrossRef]
- Hsu, C.K.; Su, S.C.; Chang, L.C.; Yang, K.J.; Lee, C.C.; Hsu, H.J.; Chen, Y.T.; Sun, C.Y.; Wu, I.W. Oral Absorbent AST-120 Is Associated with Compositional and Functional Adaptations of Gut Microbiota and Modification of Serum Short and Medium-Chain Fatty Acids in Advanced CKD Patients. Biomedicines 2022, 10, 2234. [Google Scholar] [CrossRef]
- Lau, W.L.; Vaziri, N.D.; Nunes, A.C.F.; Comeau, A.M.; Langille, M.G.I.; England, W.; Khazaeli, M.; Suematsu, Y.; Phan, J.; Whiteson, K. The Phosphate Binder Ferric Citrate Alters the Gut Microbiome in Rats with Chronic Kidney Disease. J. Pharmacol. Exp. Ther. 2018, 367, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Iguchi, A.; Yamamoto, S.; Oda, A.; Tanaka, K.; Kazama, J.J.; Saeki, T.; Yamazaki, H.; Ishioka, K.; Suzutani, T.; Narita, I. Effect of Sucroferric Oxyhydroxide on Gastrointestinal Microbiome and Uremic Toxins in Patients with Chronic Kidney Disease Undergoing Hemodialysis. Clin. Exp. Nephrol. 2020, 24, 725–733. [Google Scholar] [CrossRef]
- Abdelbary, M.M.H.; Kuppe, C.; Michael, S.S.Y.; Krüger, T.; Floege, J.; Conrads, G. Impact of Sucroferric Oxyhydroxide on the Oral and Intestinal Microbiome in Hemodialysis Patients. Sci. Rep. 2022, 12, 9614. [Google Scholar] [CrossRef] [PubMed]
- Merino-Ribas, A.; Araujo, R.; Bancu, I.; Graterol, F.; Vergara, A.; Noguera-Julian, M.; Paredes, R.; Bonal, J.; Sampaio-Maia, B. Gut Microbiome in Hemodialysis Patients Treated with Calcium Acetate or Treated with Sucroferric Oxyhydroxide: A Pilot Study. Int. Urol. Nephrol. 2022, 54, 2015. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| References | Sample/Design | Intervention | Results |
|---|---|---|---|
| Polyphenols | |||
| Delgadillo-Puga et al. (2023) [114] | Male C57BL/6 mice and INS-1E cells or C2C12 myotube | Groups: control diet, HFD, HFD + 10% VFP (Vachellia farnesiana pods), or 0.5%, 1%, or 2% or VFPE (Vachellia farnesiana polyphenolic extract) for 14 weeks Cells treated with 0, 5, or 10 mg/mL of VFPE for 2 h | VFP (Vachellia farnesiana pods) or VFPE (Vachellia farnesiana polyphenolic extract: Prevented glomerular damage, insulin resistance, hepatic steatosis, ↓ Abundance of Desulfovibrionaceae, Enterobacteriales, Gram-positive bacteria Erysipelotrichales, Chloroplast, Mollicutes, and Mycoplasmatales INS-1E cells: ↓ insulin secretion C2C12 myotubes: ↑ mitochondrial activity |
| Ren et al. (2022) [116] | C57BL/6J mice treated with Lophius litulon peptides (LPs) isolated from Monkfish | HFD (saline), HFD + LP–100 mg/kg and HFD + LP–200 mg/kg | LP: ↓ kidney damage ↓UA, Cr, BUN ↓ IL-1β, IL-6, and TNF-α ↓ TLR4/NF-κB pathway ↑ Nrf2/Keap1 pathway ↑ out and bacterial diversity |
| Zou et al. (2022) [90] | Male C57BL/6 mice cisplatin-induced AKI | Control group; cisplatin group; Qiong-Yu-Gao (QYG) group | ↓ gene expression of kidney injury markers ↓ BUN ↓ fibrosis induced by cisplatin and inflammation diversity of bacteria Akkermansia, Faecalibaculum, Bifidobacterium and Lachnospiraceae_NK4A136 SCFA-producing bacteria |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mafra, D.; Kemp, J.A.; Borges, N.A.; Wong, M.; Stenvinkel, P. Gut Microbiota Interventions to Retain Residual Kidney Function. Toxins 2023, 15, 499. https://doi.org/10.3390/toxins15080499
Mafra D, Kemp JA, Borges NA, Wong M, Stenvinkel P. Gut Microbiota Interventions to Retain Residual Kidney Function. Toxins. 2023; 15(8):499. https://doi.org/10.3390/toxins15080499
Chicago/Turabian StyleMafra, Denise, Julie A. Kemp, Natalia A. Borges, Michelle Wong, and Peter Stenvinkel. 2023. "Gut Microbiota Interventions to Retain Residual Kidney Function" Toxins 15, no. 8: 499. https://doi.org/10.3390/toxins15080499
APA StyleMafra, D., Kemp, J. A., Borges, N. A., Wong, M., & Stenvinkel, P. (2023). Gut Microbiota Interventions to Retain Residual Kidney Function. Toxins, 15(8), 499. https://doi.org/10.3390/toxins15080499