Characterization of Sodium Channel Peptides Obtained from the Venom of the Scorpion Centruroides bonito

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Venom LD50 Determination
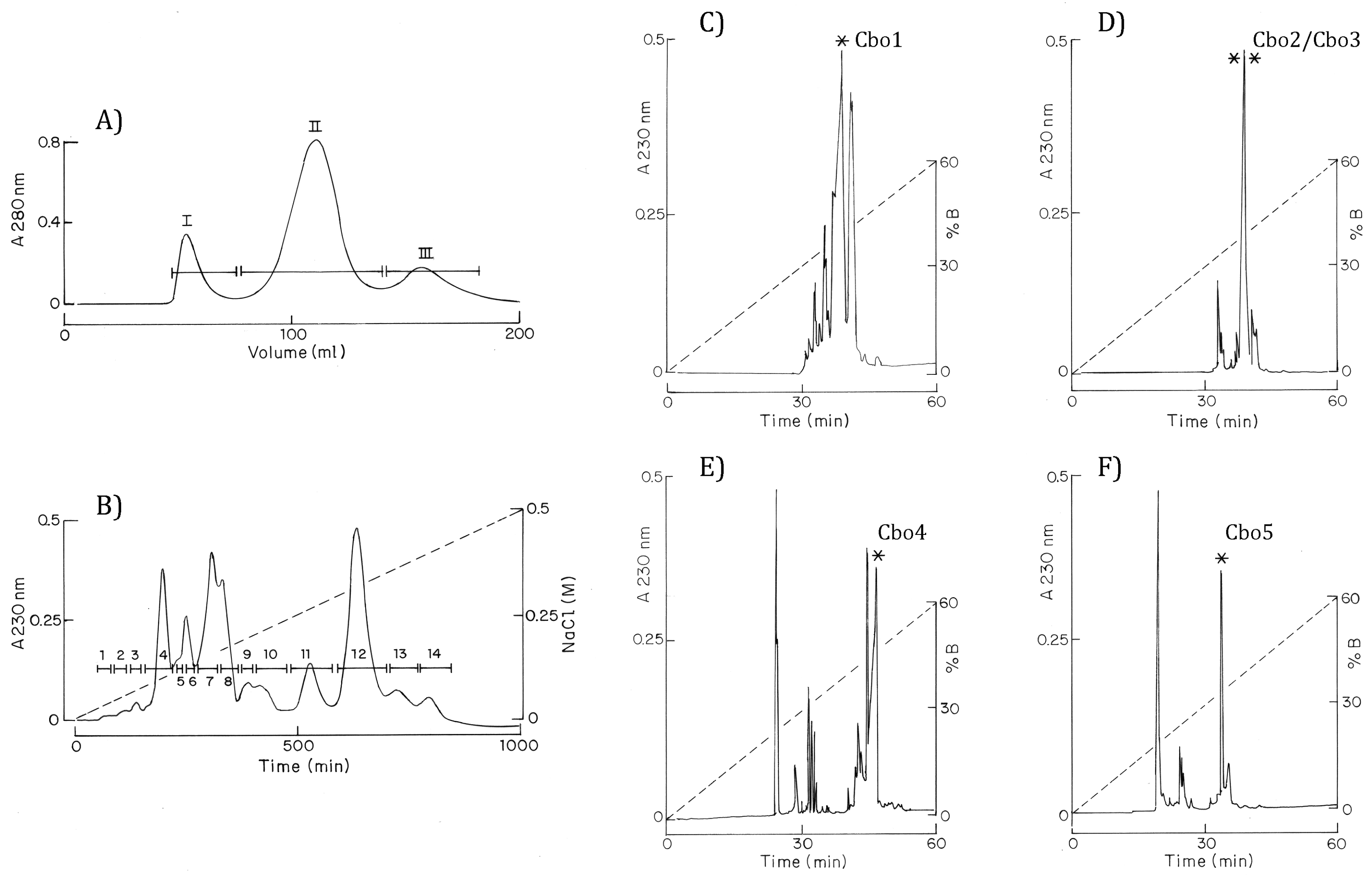
2.2. Purification and Sequencing
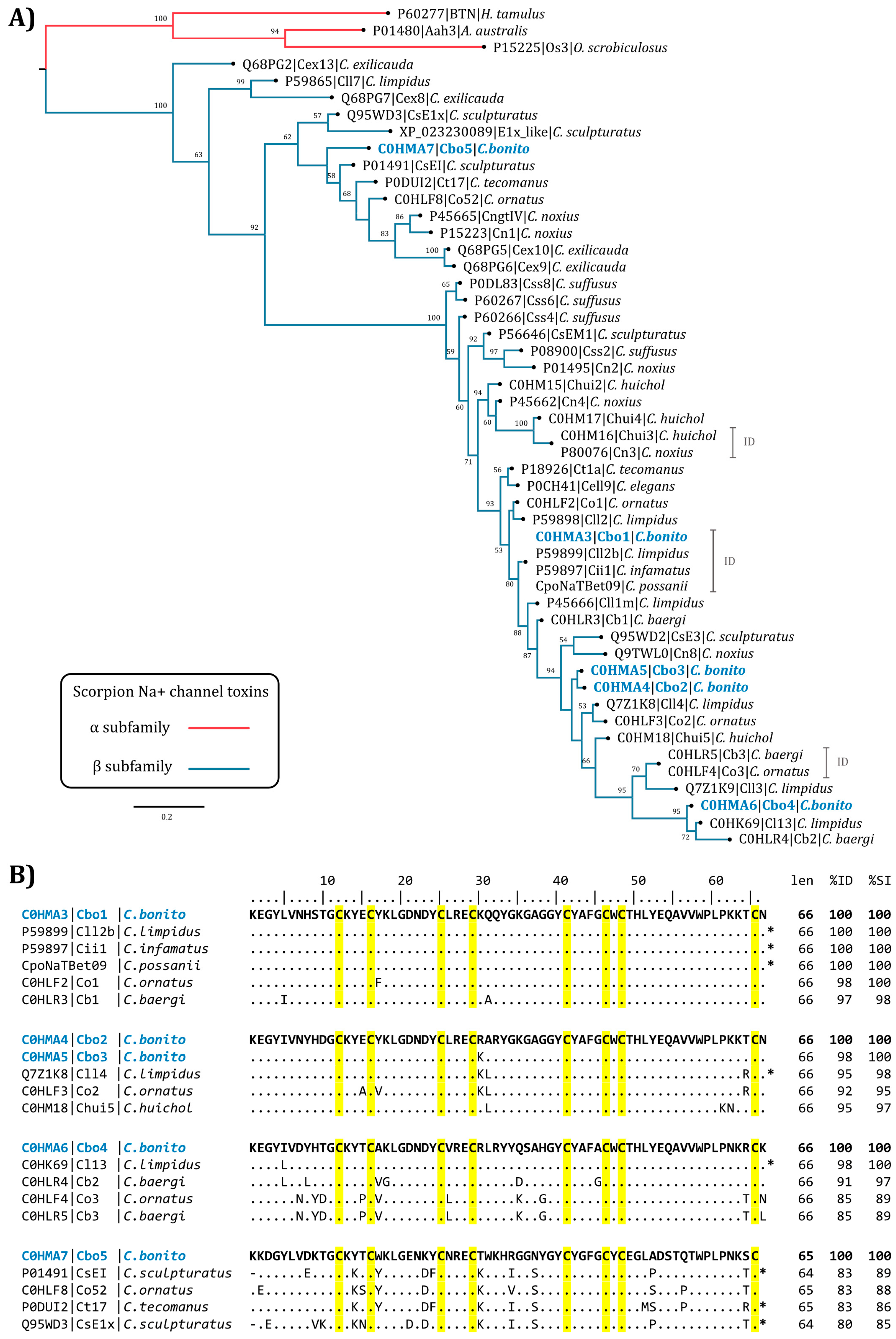
2.3. Phylogenetic Analysis
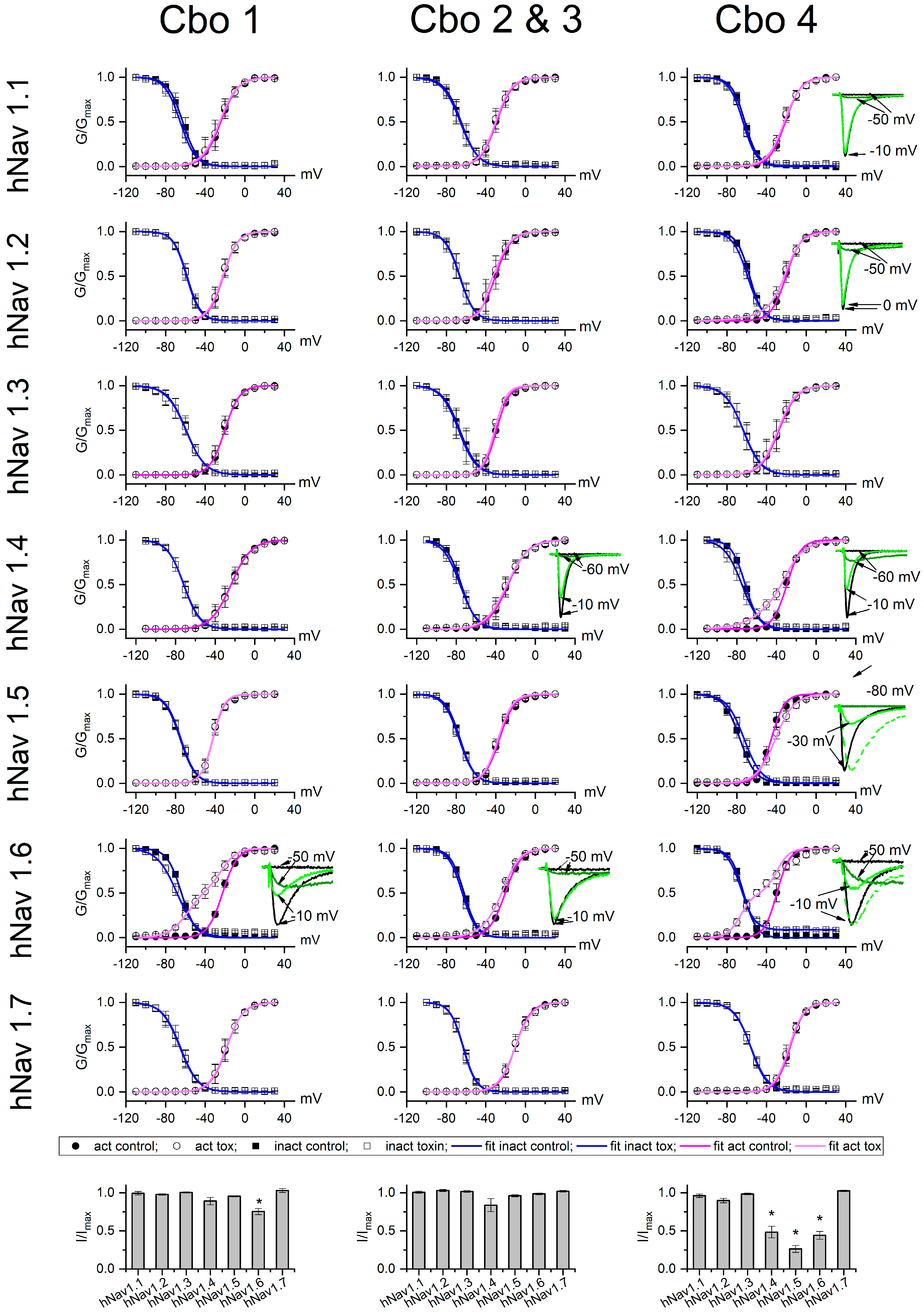
2.4. Physiological Characterization
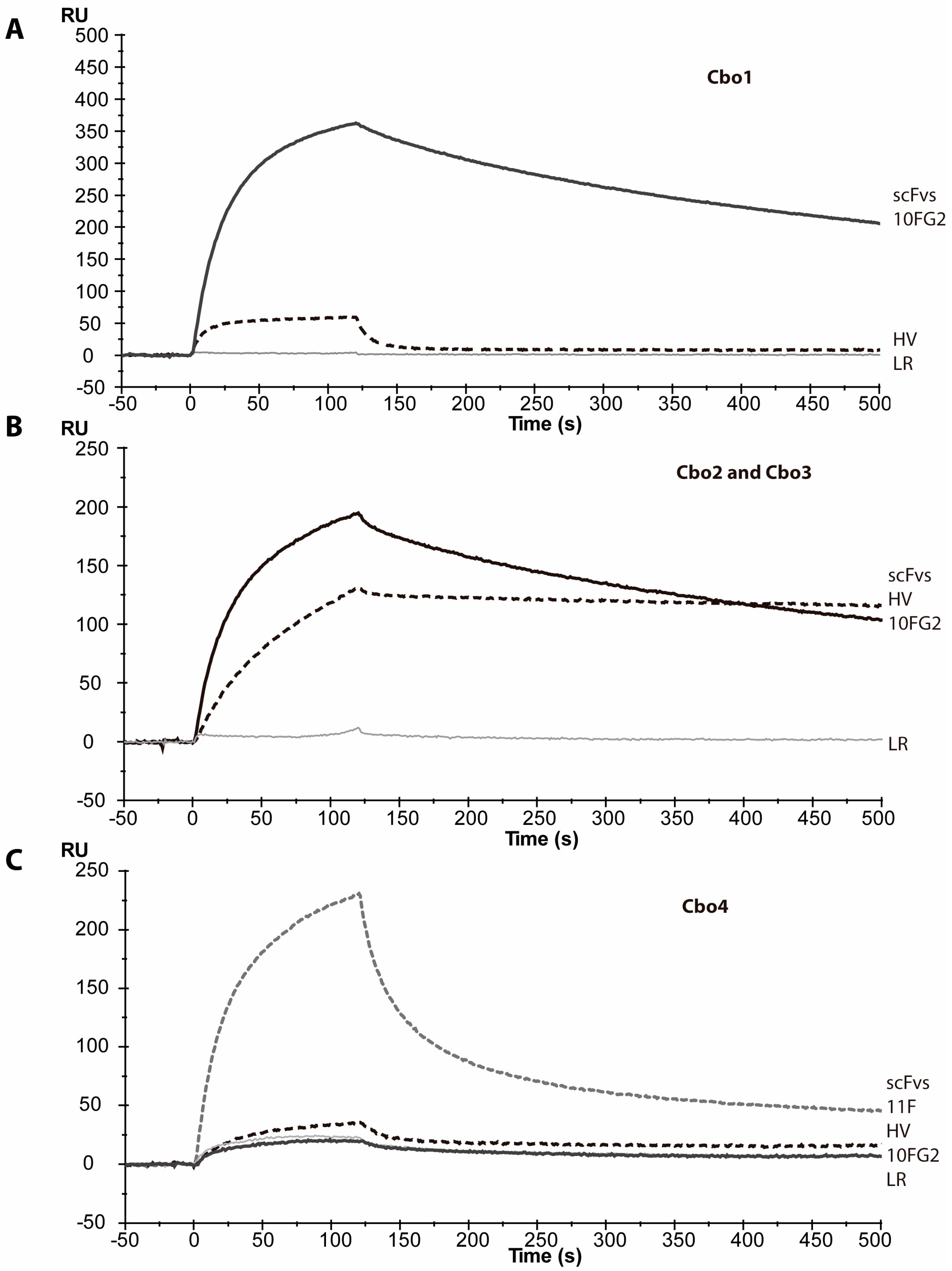
2.5. Neutralization with Single-Chains
3. Discussion
4. Materials and Methods
4.1. Venom Extraction, LD50 Determination, and Purification
4.2. Mass Spectrometry and Sequence Determination
4.3. Phylogenetic Analysis
4.4. Cell Culture and Electrophysiology Characterization
4.5. Evaluation of the Interaction of scFvs and C. bonito Toxins by Surface Plasmon Resonance
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rein, J.O. The Scorpion Files; NTNU: Trondheim, Norway, 2023. [Google Scholar]
- Chippaux, J.P.; Goyffon, M. Epidemiology of scorpionism: A global appraisal. Acta Trop. 2008, 107, 71–79. [Google Scholar] [CrossRef]
- Abroug, F.; Ouanes-Besbes, L.; Tilouche, N.; Elatrous, S. Scorpion envenomation: State of the art. Intensive Care Med. 2020, 46, 401–410. [Google Scholar] [CrossRef]
- Balozet, L. Scorpionism in the old world. In Venomous Animal and Their Venoms; Bücherl, W., Buckley, E.E., Deulofeu, V., Eds.; Academic Press: New York, NY, USA, 1971; Volume 3, pp. 349–371. [Google Scholar]
- Debont, T.; Swerts, A.; Van der Walt, J.J.; Muller, G.J.; Verdonck, F.; Daenens, P.; Tytgat, J. Comparison and characterization of the venoms of three Parabuthus scorpion species occurring in southern Africa. Toxicon Off. J. Int. Soc. Toxinol. 1998, 36, 341–352. [Google Scholar] [CrossRef]
- Mazzotti, L.; Bravo-Becherelle, M.A. Scorpionism in the Mexican Republic. In Venomous and Poisonous Animals and Noxious Plants of the Pacific Region: A Collection of Papers Based on a Symposium in the Public Health and Medical Science Division at the Tenth Pacific Science Congress; Keegan, H.L., Macfarlane, W.V., Eds.; Macmillan: London, UK, 1963; pp. 119–131. [Google Scholar]
- Bücherl, W.; Diniz, C.R. Venoms of Tityinae. In Arthropod Venoms. Handbuch der Experimentellen Pharmakologie, 1st ed.; Bettini, S., Ed.; Springer: Berlin/Heidelberg, Germany, 1978; Volume 48. [Google Scholar]
- Watt, D.D.; Simard, J.M.; Babin, D.R.; Mlejnek, R.V. Physiological characterization of toxins isolated from scorpion venom. In Toxins: Animal, Plant and Microbial; Rosenber, P., Ed.; Pergamon Press: Oxford, UK, 1978; pp. 647–660. [Google Scholar]
- Al-Asmari, A.K.; Kunnathodi, F.; Al Saadon, K.; Idris, M.M. Elemental analysis of scorpion venoms. J. Venom Res. 2016, 7, 16–20. [Google Scholar]
- Zlotkin, E.; Miranda, F.; Rochat, H. Chemistry and pharmacology of Buthinae scorpion venoms. In Handbook of Experimental Physiology; Bettini, S., Ed.; Springer: Berlin/Heidelberg, Germany, 1978; Volume 48. [Google Scholar]
- Meves, H.; Rubly, N.; Watt, D.D. Effect of toxins isolated from the venom of the scorpion Centruroides sculpturatus on the Na currents of the node of Ranvier. Pflug. Arch. Eur. J. Physiol. 1982, 393, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Neurotoxins that act on voltage-sensitive sodium channels in excitable membranes. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 15–43. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Purification of a toxic protein from scorpion venom which activates the action potential Na+ ionophore. J. Biol. Chem. 1976, 251, 5528–5536. [Google Scholar] [CrossRef] [PubMed]
- Mozhayeva, G.N.; Naumov, A.P.; Nosyreva, E.D.; Grishin, E.V. Potential-dependent interaction of toxin from venom of the scorpion Buthus eupeus with sodium channels in myelinated fibre: Voltage clamp experiments. Biochim. Biophys. Acta 1980, 597, 587–602. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.; Martin-Eauclaire, M.F.; Cestele, S.; Kopeyan, C.; Carlier, E.; Khalifa, R.B.; Pelhate, M.; Rochat, H. Scorpion toxins affecting sodium current inactivation bind to distinct homologous receptor sites on rat brain and insect sodium channels. J. Biol. Chem. 1996, 271, 8034–8045. [Google Scholar] [CrossRef] [PubMed]
- Barhanin, J.; Giglio, J.R.; Leopold, P.; Schmid, A.; Sampaio, S.V.; Lazdunski, M. Tityus serrulatus venom contains two classes of toxins. Tityus gamma toxin is a new tool with a very high affinity for studying the Na+ channel. J. Biol. Chem. 1982, 257, 12553–12558. [Google Scholar] [CrossRef] [PubMed]
- Mendes, L.C.; Viana, G.M.M.; Nencioni, A.L.A.; Pimenta, D.C.; Beraldo-Neto, E. Scorpion Peptides and Ion Channels: An Insightful Review of Mechanisms and Drug Development. Toxins 2023, 15, 238. [Google Scholar] [CrossRef] [PubMed]
- Santibanez-Lopez, C.E.; Aharon, S.; Ballesteros, J.A.; Gainett, G.; Baker, C.M.; Gonzalez-Santillan, E.; Harvey, M.S.; Hassan, M.K.; Abu Almaaty, A.H.; Aldeyarbi, S.M.; et al. Phylogenomics of Scorpions Reveal Contemporaneous Diversification of Scorpion Mammalian Predators and Mammal-Active Sodium Channel Toxins. Syst. Biol. 2022, 71, 1281–1289. [Google Scholar] [CrossRef]
- Santibanez-Lopez, C.E.; Francke, O.F.; Ureta, C.; Possani, L.D. Scorpions from Mexico: From Species Diversity to Venom Complexity. Toxins 2015, 8, 2. [Google Scholar] [CrossRef]
- Quijano-Ravell, A.F.; Teruel, R.; Ponce-Saavedra, J. A new Centruroides Marx, 1890 (Scorpiones, Buthidae) from Southern Guerrero State, Mexico. Rev. Ibérica Aracnol. 2016, 28, 25–34. [Google Scholar]
- Gonzalez-Santillan, E.; Possani, L.D. North American scorpion species of public health importance with a reappraisal of historical epidemiology. Acta Trop. 2018, 187, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Alagon, A.C.; Guzman, H.S.; Martin, B.M.; Ramirez, A.N.; Carbone, E.; Possani, L.D. Isolation and characterization of two toxins from the Mexican scorpion Centruroides limpidus limpidus Karsch. Comp. Biochem. Physiol. B Comp. Biochem. 1988, 89, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Dehesa-Davila, M.; Ramirez, A.N.; Zamudio, F.Z.; Gurrola-Briones, G.; Lievano, A.; Darszon, A.; Possani, L.D. Structural and functional comparison of toxins from the venom of the scorpions Centruroides infamatus infamatus, Centruroides limpidus limpidus and Centruroides noxius. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1996, 113, 331–339. [Google Scholar] [CrossRef]
- Garcia-Villalvazo, P.E.; Jimenez-Vargas, J.M.; Lino-Lopez, G.J.; Meneses, E.P.; Bermudez-Guzman, M.J.; Barajas-Saucedo, C.E.; Delgado Enciso, I.; Possani, L.D.; Valdez-Velazquez, L.L. Unveiling the Protein Components of the Secretory-Venom Gland and Venom of the Scorpion Centruroides possanii (Buthidae) through Omic Technologies. Toxins 2023, 15, 498. [Google Scholar] [CrossRef]
- Garcia-Guerrero, I.A.; Carcamo-Noriega, E.; Gomez-Lagunas, F.; Gonzalez-Santillan, E.; Zamudio, F.Z.; Gurrola, G.B.; Possani, L.D. Biochemical characterization of the venom from the Mexican scorpion Centruroides ornatus, a dangerous species to humans. Toxicon 2020, 173, 27–38. [Google Scholar] [CrossRef]
- Gomez-Ramirez, I.V.; Riano-Umbarila, L.; Olamendi-Portugal, T.; Restano-Cassulini, R.; Possani, L.D.; Becerril, B. Biochemical, electrophysiological and immunological characterization of the venom from Centruroides baergi, a new scorpion species of medical importance in Mexico. Toxicon Off. J. Int. Soc. Toxinol. 2020, 184, 10–18. [Google Scholar] [CrossRef]
- Valencia-Martinez, H.; Olamendi-Portugal, T.; Restano-Cassulini, R.; Serrano-Posada, H.; Zamudio, F.; Possani, L.D.; Riano-Umbarila, L.; Becerril, B. Characterization of Four Medically Important Toxins from Centruroides huichol Scorpion Venom and Its Neutralization by a Single Recombinant Antibody Fragment. Toxins 2022, 14, 369. [Google Scholar] [CrossRef]
- Lopez-Giraldo, A.E.; Olamendi-Portugal, T.; Riano-Umbarila, L.; Becerril, B.; Possani, L.D.; Delepierre, M.; Del Rio-Portilla, F. The three-dimensional structure of the toxic peptide Cl13 from the scorpion Centruroides limpidus. Toxicon Off. J. Int. Soc. Toxinol. 2020, 184, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Babin, D.R.; Watt, D.D.; Goos, S.M.; Mlejnek, R.V. Amino acid sequence of neurotoxin I from Centruroides sculpturatus Ewing. Arch. Biochem. Biophys. 1975, 166, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Cestele, S.; Qu, Y.; Rogers, J.C.; Rochat, H.; Scheuer, T.; Catterall, W.A. Voltage sensor-trapping: Enhanced activation of sodium channels by beta-scorpion toxin bound to the S3-S4 loop in domain II. Neuron 1998, 21, 919–931. [Google Scholar] [CrossRef]
- Fernandez-Taboada, G.; Riano-Umbarila, L.; Olvera-Rodriguez, A.; Gomez-Ramirez, I.V.; Losoya-Uribe, L.F.; Becerril, B. The venom of the scorpion Centruroides limpidus, which causes the highest number of stings in Mexico, is neutralized by two recombinant antibody fragments. Mol. Immunol. 2021, 137, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Riano-Umbarila, L.; Contreras-Ferrat, G.; Olamendi-Portugal, T.; Morelos-Juarez, C.; Corzo, G.; Possani, L.D.; Becerril, B. Exploiting cross-reactivity to neutralize two different scorpion venoms with one single chain antibody fragment. J. Biol. Chem. 2011, 286, 6143–6151. [Google Scholar] [CrossRef]
- Riano-Umbarila, L.; Gomez-Ramirez, I.V.; Ledezma-Candanoza, L.M.; Olamendi-Portugal, T.; Rodriguez-Rodriguez, E.R.; Fernandez-Taboada, G.; Possani, L.D.; Becerril, B. Generation of a Broadly Cross-Neutralizing Antibody Fragment against Several Mexican Scorpion Venoms. Toxins 2019, 11, 32. [Google Scholar] [CrossRef]
- Rendon-Anaya, M.; Delaye, L.; Possani, L.D.; Herrera-Estrella, A. Global transcriptome analysis of the scorpion Centruroides noxius: New toxin families and evolutionary insights from an ancestral scorpion species. PLoS ONE 2012, 7, e43331. [Google Scholar] [CrossRef]
- Vazquez, A.; Tapia, J.V.; Eliason, W.K.; Martin, B.M.; Lebreton, F.; Delepierre, M.; Possani, L.D.; Becerril, B. Cloning and characterization of the cDNAs encoding Na+ channel-specific toxins 1 and 2 of the scorpion Centruroides noxius Hoffmann. Toxicon Off. J. Int. Soc. Toxinol. 1995, 33, 1161–1170. [Google Scholar] [CrossRef]
- Valdez-Velazquez, L.L.; Romero-Gutierrez, M.T.; Delgado-Enciso, I.; Dobrovinskaya, O.; Melnikov, V.; Quintero-Hernandez, V.; Ceballos-Magana, S.G.; Gaitan-Hinojosa, M.A.; Coronas, F.I.; Puebla-Perez, A.M.; et al. Comprehensive analysis of venom from the scorpion Centruroides tecomanus reveals compounds with antimicrobial, cytotoxic, and insecticidal activities. Toxicon 2016, 118, 95–103. [Google Scholar] [CrossRef]
- Herzig, V.; Ikonomopoulou, M.; Smith, J.J.; Dziemborowicz, S.; Gilchrist, J.; Kuhn-Nentwig, L.; Rezende, F.O.; Moreira, L.A.; Nicholson, G.M.; Bosmans, F.; et al. Molecular basis of the remarkable species selectivity of an insecticidal sodium channel toxin from the African spider Augacephalus ezendami. Sci. Rep. 2016, 6, 29538. [Google Scholar] [CrossRef]
- Kasheverov, I.E.; Oparin, P.B.; Zhmak, M.N.; Egorova, N.S.; Ivanov, I.A.; Gigolaev, A.M.; Nekrasova, O.V.; Serebryakova, M.V.; Kudryavtsev, D.S.; Prokopev, N.A.; et al. Scorpion toxins interact with nicotinic acetylcholine receptors. FEBS Lett. 2019, 593, 2779–2789. [Google Scholar] [CrossRef]
- Olamendi-Portugal, T.; Restano-Cassulini, R.; Riano-Umbarila, L.; Becerril, B.; Possani, L.D. Functional and immuno-reactive characterization of a previously undescribed peptide from the venom of the scorpion Centruroides limpidus. Peptides 2017, 87, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Pucca, M.B.; Cerni, F.A.; Janke, R.; Bermudez-Mendez, E.; Ledsgaard, L.; Barbosa, J.E.; Laustsen, A.H. History of Envenoming Therapy and Current Perspectives. Front. Immunol. 2019, 10, 1598. [Google Scholar] [CrossRef] [PubMed]
- Roncolato, E.C.; Campos, L.B.; Pessenda, G.; Costa e Silva, L.; Furtado, G.P.; Barbosa, J.E. Phage display as a novel promising antivenom therapy: A review. Toxicon Off. J. Int. Soc. Toxinol. 2015, 93, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Alonso Villela, S.M.; Kraiem-Ghezal, H.; Bouhaouala-Zahar, B.; Bideaux, C.; Aceves Lara, C.A.; Fillaudeau, L. Production of recombinant scorpion antivenoms in E. coli: Current state and perspectives. Appl. Microbiol. Biotechnol. 2023, 107, 4133–4152. [Google Scholar] [CrossRef]
- von Reumont, B.M.; Anderluh, G.; Antunes, A.; Ayvazyan, N.; Beis, D.; Caliskan, F.; Crnkovic, A.; Damm, M.; Dutertre, S.; Ellgaard, L.; et al. Modern venomics-Current insights, novel methods, and future perspectives in biological and applied animal venom research. Gigascience 2022, 11, giac048. [Google Scholar] [PubMed]
- Dixon, W.J.; Mood, A.M. A Method for Obtaining and Analyzing Sensitivity Data. J. Am. Stat. Assoc. 1948, 43, 109–126. [Google Scholar] [CrossRef]
- Shakeel, K.; Olamendi-Portugal, T.; Naseem, M.U.; Becerril, B.; Zamudio, F.Z.; Delgado-Prudencio, G.; Possani, L.D.; Panyi, G. Of Seven New K(+) Channel Inhibitor Peptides of Centruroides bonito, alpha-KTx 2.24 Has a Picomolar Affinity for Kv1.2. Toxins 2023, 15, 506. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Whelan, S.; Goldman, N. A general empirical model of protein evolution derived from multiple protein families using a maximum-likelihood approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Stothard, P. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. Biotechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef]
- Schiavon, E.; Sacco, T.; Cassulini, R.R.; Gurrola, G.; Tempia, F.; Possani, L.D.; Wanke, E. Resurgent current and voltage sensor trapping enhanced activation by a beta-scorpion toxin solely in Nav1.6 channel. Significance in mice Purkinje neurons. J. Biol. Chem. 2006, 281, 20326–20337. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Restano-Cassulini, R.; Olamendi-Portugal, T.; Riaño-Umbarila, L.; Zamudio, F.Z.; Delgado-Prudencio, G.; Becerril, B.; Possani, L.D. Characterization of Sodium Channel Peptides Obtained from the Venom of the Scorpion Centruroides bonito. Toxins 2024, 16, 125. https://doi.org/10.3390/toxins16030125
Restano-Cassulini R, Olamendi-Portugal T, Riaño-Umbarila L, Zamudio FZ, Delgado-Prudencio G, Becerril B, Possani LD. Characterization of Sodium Channel Peptides Obtained from the Venom of the Scorpion Centruroides bonito. Toxins. 2024; 16(3):125. https://doi.org/10.3390/toxins16030125
Chicago/Turabian StyleRestano-Cassulini, Rita, Timoteo Olamendi-Portugal, Lidia Riaño-Umbarila, Fernando Z. Zamudio, Gustavo Delgado-Prudencio, Baltazar Becerril, and Lourival D. Possani. 2024. "Characterization of Sodium Channel Peptides Obtained from the Venom of the Scorpion Centruroides bonito" Toxins 16, no. 3: 125. https://doi.org/10.3390/toxins16030125
APA StyleRestano-Cassulini, R., Olamendi-Portugal, T., Riaño-Umbarila, L., Zamudio, F. Z., Delgado-Prudencio, G., Becerril, B., & Possani, L. D. (2024). Characterization of Sodium Channel Peptides Obtained from the Venom of the Scorpion Centruroides bonito. Toxins, 16(3), 125. https://doi.org/10.3390/toxins16030125