Structure and Function of Snake Venom Proteins Affecting Platelet Plug Formation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
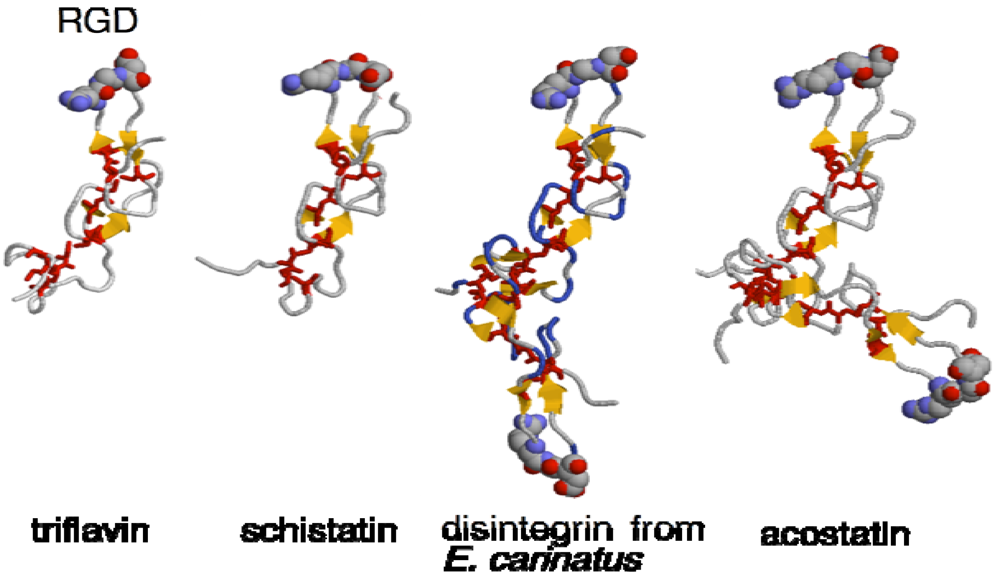
2. Structure and Function of Disintegrins

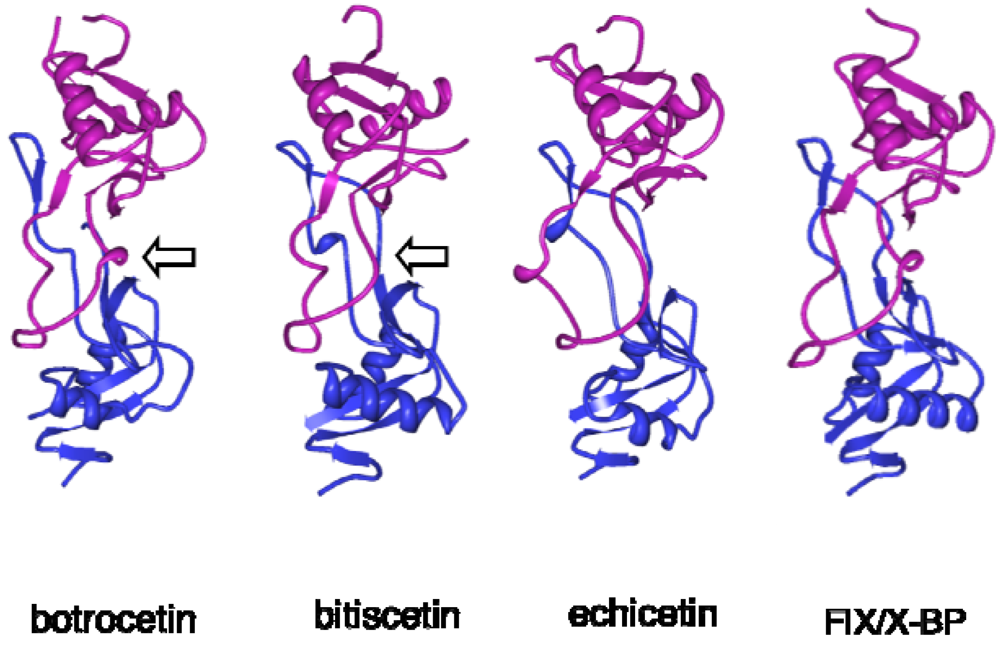
3. Structure and Function of VWF-Modulating Snake Venom Proteins


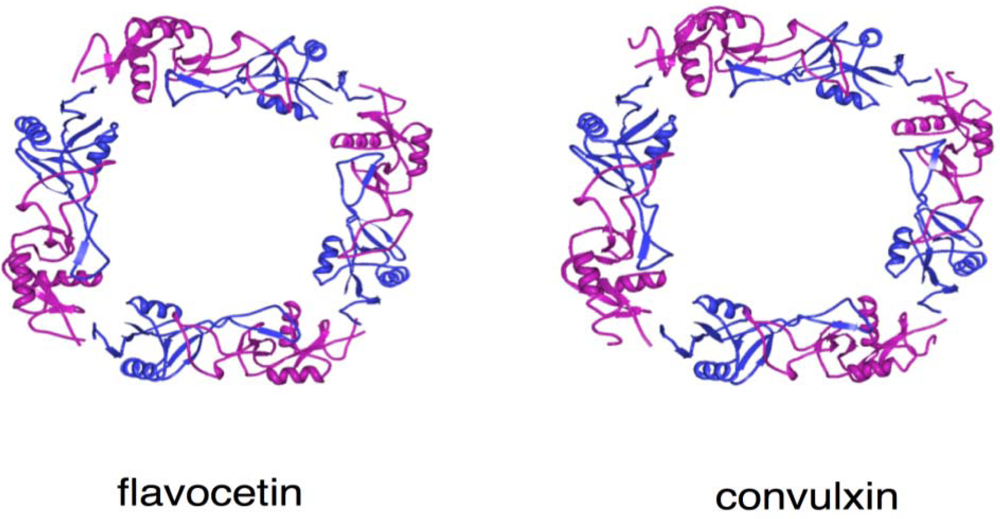
4. Structure and Function of GPIb-binding Snake Venom Proteins

5. Perspectives
References
- Sadler, J.E. von Willebrand factor: Two sides of a coin. J. Thromb. Haemost. 2005, 3, 1702–1709. [Google Scholar] [PubMed]
- Ruggeri, Z.M. The role of von Willebrand factor in thrombus formation. Thromb. Res. 2007, 120 (Suppl 1), S5–S9. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, C.; Teng, C.M.; Huang, T.F. Characterization of snake venom components acting on blood coagulation and platelet function. Toxicon 1992, 30, 945–966. [Google Scholar] [PubMed]
- Matsui, T.; Fujimura, Y.; Titani, K. Snake venom proteases affecting hemostasis and thrombosis. Biochim. Biophys. Acta 2000, 1477, 146–156. [Google Scholar] [PubMed]
- Matsui, T.; Hamako, J. Structure and function of snake venom toxins interacting with human von Willebrand factor. Toxicon 2005, 45, 1075–1087. [Google Scholar] [PubMed]
- Andrews, R.K.; Gardiner, E.E.; Berndt, M.C. Snake venom toxins affecting platelet function. Methods Mol. Biol. 2004, 273, 335–348. [Google Scholar] [PubMed]
- Marsh, N.A. Diagnostic uses of snake venom. Haemostasis 2001, 31, 211–217. [Google Scholar] [PubMed]
- Marsh, N.; Williams, V. Practical applications of snake venom toxins in haemostasis. Toxicon 2005, 45, 1171–1181. [Google Scholar] [PubMed]
- Clemetson, K.J.; Clemetson, J.M. Platelet GPIb complex as a target for anti-thrombotic drug development. Thromb. Haemost. 2008, 99, 473–479. [Google Scholar] [PubMed]
- Huang, T.F.; Holt, J.C.; Lukasiewicz, H.; Niewiarowski, S. Trigramin. A low molecular weight peptide inhibiting fibrinogen interaction with platelet receptors expressed on glycoprotein IIb-IIIa complex. J. Biol. Chem. 1987, 262, 16157–16163. [Google Scholar] [PubMed]
- Huang, T.F.; Holt, J.C.; Kirby, E.P.; Niewiarowski, S. Trigramin: Primary structure and its inhibition of von Willebrand factor binding to glycoprotein IIb/IIIa complex on human platelets. Biochemistry 1989, 28, 661–666. [Google Scholar] [PubMed]
- McLane, M.A.; Paquette-Straub, C. Scientific and standardization committee communications: Classification and nomenclature of disintegrins isolated from snake venoms. J. Thromb. Haemost. 2007, 5, 1971. [Google Scholar] [PubMed]
- McLane, M.A.; Sanchez, E.E.; Wong, A.; Paquette-Straub, C.; Perez, J.C. Disintegrins. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2004, 4, 327–355. [Google Scholar] [PubMed]
- Moiseeva, N.; Bau, R.; Swenson, S.D.; Markland, F.S., Jr.; Choe, J.Y.; Liu, Z.J.; Allaire, M. Structure of acostatin, a dimeric disintegrin from Southern copperhead (Agkistrodon contortrix contortrix), a.7 A resolution. Acta Crystallogr. D. Biol. Crystallogr. 2008, 64, 466–470. [Google Scholar] [PubMed]
- Adler, M.; Lazarus, R.A.; Dennis, M.S.; Wagner, G. Solution structure of kistrin, a potent platelet aggregation inhibitor and GP IIb-IIIa antagonis. Science 1991, 253, 445–448. [Google Scholar] [PubMed]
- Monleon, D.; Esteve, V.; Kovacs, H.; Calvete, J.J.; Celda, B. Conformation and concerted dynamics of the integrin-binding site and the C-terminal region of echistatin revealed by homonuclear NMR. Biochem. J. 2005, 387, 57–66. [Google Scholar] [PubMed]
- Fujii, Y.; Okuda, D.; Fujimoto, Z.; Horii, K.; Morita, T.; Mizuno, H. Crystal structure of trimestatin, a disintegrin containing a cell adhesion recognition motif RG. J. Mol. Biol. 2003, 332, 1115–1122. [Google Scholar] [PubMed]
- Shin, J.; Hong, S.Y.; Chung, K.; Kang, I.; Jang, Y.; Kim, D.S.; Lee, W. Solution structure of a novel disintegrin, salmosin, from Agkistrondon halys ven. Biochemistry 2003, 42, 14408–14415. [Google Scholar] [PubMed]
- Bilgrami, S.; Tomar, S.; Yadav, S.; Kaur, P.; Kumar, J.; Jabeen, T.; Sharma, S.; Singh, T.P. Crystal structure of schistatin, a disintegrin homodimer from saw-scaled viper (Echis carinatus) at .5 A resolution. J. Mol. Biol. 2004, 341, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Bilgrami, S.; Yadav, S.; Kaur, P.; Sharma, S.; Perbandt, M.; Betzel, C.; Singh, T.P. Crystal structure of the disintegrin heterodimer from saw-scaled viper (Echis carinatus) at 1.9 A resolution. Biochemistry 2005, 44, 11058–11066. [Google Scholar] [PubMed]
- Sayle, R.A.; Milner-White, E.J. RASMOL: Biomolecular graphics for all. Trends. Biochem. Sci. 1995, 20, 374. [Google Scholar] [PubMed]
- Kim, J.; Hong, S.Y.; Park, H.S.; Kim, D.S.; Lee, W. Structure and function of RGD peptides derived from disintegrin proteins. Mol. Cells 2005, 19, 205–211. [Google Scholar] [PubMed]
- Scarborough, R.M.; Rose, J.W.; Hsu, M.A.; Phillips, D.R.; Fried, V.A.; Campbell, A.M.; Nannizzi, L.; Charo, I.F. Barbourin. A GPIIb-IIIa-specific integrin antagonist from the venom of Sistrurus m. barbouri. J. Biol. Chem. 1991, 266, 9359–9362. [Google Scholar] [PubMed]
- Scarborough, R.M.; Naughton, M.A.; Teng, W.; Rose, J.W.; Phillips, D.R.; Nannizzi, L.; Arfsten, A.; Campbell, A.M.; Charo, I.F. Design of potent and specific integrin antagonists. Peptide antagonists with high specificity for glycoprotein IIb-IIIa. J. Biol. Chem 1993, 268, 1066–1073. [Google Scholar] [PubMed]
- Scarborough, R.M. Development of eptifibatide. Am. Heart. J. 1999, 138, 1093–1104. [Google Scholar] [PubMed]
- Phillips, D.R.; Scarborough, R.M. Clinical pharmacology of eptifibatide. Am. J. Cardiol. 80, 11–20. [PubMed]
- Plosker, G.L.; Ibbotson, T. Eptifibatide: A pharmacoeconomic review of its use in percutaneous coronary intervention and acute coronary syndromes. Pharmacoeconomics 2003, 21, 885–912. [Google Scholar] [PubMed]
- Zeymer, U.; Wienbergen, H. A review of clinical trials with eptifibatide in cardiology. Cardiovasc. Drug Rev. 2007, 25, 301–315. [Google Scholar] [PubMed]
- Sheffield, W.P.; Gataiance, S.; Eltringham-Smith, L.J. Combined administration of barbourin-albumin and hirudin-albumin fusion proteins limits fibrin(ogen) deposition on the rabbit balloon-injured aorta. Thromb. Res. 2007, 119, 195–207. [Google Scholar] [PubMed]
- Lu, X.; Lu, D.; Scully, M.F.; Kakkar, V.V. Snake venom metalloproteinase containing a disintegrin-like domain, its structure-activity relationships at interacting with integrin. Curr. Med. Chem. Cardiovasc. Hematol. Agents 2005, 3, 249–260. [Google Scholar] [PubMed]
- McLane, M.A.; Marcinkiewicz, C.; Vijay-Kumar, S.; Wierzbicka-Patynowski, I.; Niewiarowski, S. Viper venom disintegrins and related molecules. Proc. Soc. Exp. Biol. Med. 1998, 219, 109–119. [Google Scholar] [PubMed]
- Myles, D.G.; Kimmel, L.H.; Blobel, C.P.; White, J.M.; Primakoff, P. Identification of a binding site in the disintegrin domain of fertilin required for sperm-egg fusion. Proc. Natl. Acad. Sci. USA 1994, 91, 4195–4198. [Google Scholar]
- Yagami-Hiromasa, T.; Sato, T.; Kurisaki, T.; Kamijo, K.; Nabeshima, Y.; Fujisawa-Sehara, A. A metalloprotease-disintegrin participating in myoblast fusion. Nature 1995, 377, 652–656. [Google Scholar] [PubMed]
- Blobel, C.P. Metalloprotease-disintegrins: Links to cell adhesion and cleavage of TNF alpha and Notch. Cell 1997, 90, 589–592. [Google Scholar] [PubMed]
- Ito, M.; Hamako, J.; Sakurai, Y.; Matsumoto, M.; Fujimura, Y.; Suzuki, M.; Hashimoto, K.; Titani, K.; Matsui, T. Complete amino acid sequence of kaouthiagin, a novel cobra venom metalloproteinase with two disintegrin-like sequence. Biochemistry 2001, 40, 4503–4511. [Google Scholar] [PubMed]
- Hamako, J.; Matsui, T.; Nishida, S.; Nomura, S.; Fujimura, Y.; Ito, M.; Ozeki, Y.; Titani, K. Purification and characterization of kaouthiagin, a von Willebrand factor-binding and -cleaving metalloproteinase from Naha kaouthia cobra veno. Thromb. Haemost. 1998, 80, 499–505. [Google Scholar] [PubMed]
- Brinkhous, K.M.; Fricke, W.A.; Read, M.S. Determinants of von Willebrand factor activity elicited by ristocetin and botrocetin: Studies on a human von Willebrand factor-binding antibody. Semin. Thromb. Hemost. 1985, 11, 337–341. [Google Scholar] [PubMed]
- Scott, J.P.; Montgomery, R.R.; Retzinger, G.S. Dimeric ristocetin flocculates proteins, binds to platelets, and mediates von Willebrand factor-dependent agglutination of platele. J. Biol. Chem. 1991, 266, 8149–8155. [Google Scholar] [PubMed]
- Read, M.S.; Shermer, R.W.; Brinkhous, K.M. Venom coagglutinin: An activator of platelet aggregation dependent on von Willebrand factor. Proc. Natl. Acad. Sci. USA 1978, 75, 4514–4518. [Google Scholar]
- Read, M.S.; Smith, S.V.; Lamb, M.A.; Brinkhous, K.M. Role of botrocetin in platelet agglutination: Formation of an activated complex of botrocetin and von Willebrand factor. Blood 1989, 74, 1031–1035. [Google Scholar] [PubMed]
- Usami, Y.; Fujimura, Y.; Suzuki, M.; Ozeki, Y.; Nishio, K.; Fukui, H.; Titani, K. Primary structure of two-chain botrocetin, a von Willebrand factor modulator purified from the venom of Bothrops jararac. Proc. Natl. Acad. Sci. USA 1993, 90, 928–932. [Google Scholar]
- Matsui, T.; Hamako, J.; Suzuki, M.; Hayashi, N.; Ito, M.; Makita, K.; Fujimura, Y.; Ozeki, Y.; Titani, K. Complete amino acid sequence of bitiscetin, a novel von Willebrand factor modulator protein, purified from snake venom of Bitis arieta. Res. Commun. Biochem. Cell Mol. Biol. 1997, 1, 271–284. [Google Scholar]
- Kawasaki, T.; Fujimura, Y.; Usami, Y.; Suzuki, M.; Miura, S.; Sakurai, Y.; Makita, K.; Taniuchi, Y.; Hirano, K.; Titani, K. Complete amino acid sequence and identification of the platelet glycoprotein Ib-binding site of jararaca GPIb-BP, a snake venom protein isolated from Bothrops jararac. J. Biol. Chem. 1996, 271, 10635–10639. [Google Scholar] [PubMed]
- Fukuda, K.; Doggett, T.; Laurenzi, I.J.; Liddington, R.C.; Diacovo, T.G. The snake venom protein botrocetin acts as a biological brace to promote dysfunctional platelet aggregation. Nat. Struct. Mol. Biol. 2005, 12, 152–159. [Google Scholar] [PubMed]
- Sen, U.; Vasudevan, S.; Subbarao, G.; McClintock, R.A.; Celikel, R.; Ruggeri, Z.M.; Varughese, K.I. Crystal structure of the von Willebrand factor modulator botrocetin. Biochemistry 2001, 40, 345–352. [Google Scholar] [PubMed]
- Hirotsu, S.; Mizuno, H.; Fukuda, K.; Qi, M.C.; Matsui, T.; Hamako, J.; Morita, T.; Titani, K. Crystal structure of bitiscetin, a von Willebrand factor-dependent platelet aggregation induce. Biochemistry 2001, 40, 13592–13597. [Google Scholar] [PubMed]
- Matsushita, T.; Meyer, D.; Sadler, J.E. Localization of von willebrand factor-binding sites for platelet glycoprotein Ib and botrocetin by charged-to-alanine scanning mutagenesis. J. Biol. Chem. 2000, 275, 11044–11049. [Google Scholar] [PubMed]
- Maita, N.; Nishio, K.; Nishimoto, E.; Matsui, T.; Shikamoto, Y.; Morita, T.; Sadler, J.E.; Mizuno, H. Crystal structure of von Willebrand factor A1 domain complexed with snake venom, bitiscetin: Insight into glycoprotein Ibalpha binding mechanism induced by snake venom protein. J. Biol. Chem. 2003, 278, 37777–37781. [Google Scholar] [PubMed]
- Matsui, T.; Hamako, J.; Matsushita, T.; Nakayama, T.; Fujimura, Y.; Titani, K. Binding site on human von Willebrand factor of bitiscetin, a snake venom-derived platelet aggregation induce. Biochemistry 2002, 41, 7939–7946. [Google Scholar] [PubMed]
- Obert, B.; Romijn, R.A.; Houllier, A.; Huizinga, E.G.; Girma, J.P. Characterization of bitiscetin-2, a second form of bitiscetin from the venom of Bitis arietans : Comparison of its binding site with the collagen-binding site on the von Willebrand factor A3-domai. J. Thromb. Haemost. 2006, 4, 1596–1601. [Google Scholar] [PubMed]
- Mizuno, H.; Fujimoto, Z.; Koizumi, M.; Kano, H.; Atoda, H.; Morita, T. Structure of coagulation factors IX/X-binding protein, a heterodimer of C-type lectin domain. Nat. Struct. Biol. 1997, 4, 438–441. [Google Scholar] [PubMed]
- Atoda, H.; Ishikawa, M.; Mizuno, H.; Morita, T. Coagulation factor X-binding protein from Deinagkistrodon acutus venom is a Gla domain-binding protein. Biochemistry 1998, 37, 17361–17370. [Google Scholar] [PubMed]
- Fukuda, K.; Mizuno, H.; Atoda, H.; Morita, T. Crystallization and preliminary x-ray studies of flavocetin-A, a platelet glycoprotein Ib-binding protein from the habu snake veno. Acta Crystallogr. D. Biol. Crystallogr. 1999, 55, 1911–1913. [Google Scholar] [PubMed]
- Murakami, M.T.; Zela, S.P.; Gava, L.M.; Michelan-Duarte, S.; Cintra, A.C.; Arni, R.K. Crystal structure of the platelet activator convulxin, a disulfide-linked alpha4beta4 cyclic tetramer from the venom of Crotalus durissus terrificu. Biochem. Biophys. Res. Commun. 2003, 310, 478–482. [Google Scholar] [PubMed]
- Moreland, J.L.; Gramada, A.; Buzko, O.V.; Zhang, Q.; Bourne, P.E. The Molecular Biology Toolkit (MBT): A modular platform for developing molecular visualization applications. BMC Bioinf. 2005, 6, 21. [Google Scholar]
- Miura, S.; Li, C.Q.; Cao, Z.; Wang, H.; Wardell, M.R.; Sadler, J.E. Interaction of von Willebrand factor domain A1 with platelet glycoprotein Ibalpha-(1-289). Slow intrinsic binding kinetics mediate rapid platelet adhesion. J. Biol. Chem. 2000, 275, 7539–7546. [Google Scholar] [PubMed]
- Fukuda, K.; Doggett, T.A.; Bankston, L.A.; Cruz, M.A.; Diacovo, T.G.; Liddington, R.C. Structural basis of von Willebrand factor activation by the snake toxin botrocetin. Structure 2002, 10, 943–950. [Google Scholar] [PubMed]
- Marchese, P.; Murata, M.; Mazzucato, M.; Pradella, P.; De Marco, L.; Ware, J.; Ruggeri, Z.M. Identification of three tyrosine residues of glycoprotein Ib alpha with distinct roles in von Willebrand factor and alpha-thrombin binding. J. Biol. Chem. 1995, 270, 9571–9578. [Google Scholar] [PubMed]
- Dong, J.; Ye, P.; Schade, A.J.; Gao, S.; Romo, G.M.; Turner, N.T.; McIntire, L.V.; Lopez, J.A. Tyrosine sulfation of glycoprotein I(b)alpha. Role of electrostatic interactions in von Willebrand factor binding. J. Biol. Chem. 2001, 276, 16690–16694. [Google Scholar] [PubMed]
- Peng, M.; Lu, W.; Kirby, E.P. Alboaggregin-B: A new platelet agonist that binds to platelet membrane glycoprotein Ib. Biochemistry 1991, 30, 11529–11536. [Google Scholar] [PubMed]
- Peng, M.; Lu, W.; Kirby, E.P. Characterization of three alboaggregins purified from Trimeresurus albolabris venom. Thromb. Haemost. 1992, 67, 702–707. [Google Scholar] [PubMed]
- Sakurai, Y.; Fujimura, Y.; Kokubo, T.; Imamura, K.; Kawasaki, T.; Handa, M.; Suzuki, M.; Matsui, T.; Titani, K.; Yoshioka, A. The cDNA cloning and molecular characterization of a snake venom platelet glycoprotein Ib-binding protein, mamushigin, from Agkistrodon halys blomhoffii ven. Thromb. Haemost. 1998, 79, 1199–1207. [Google Scholar] [PubMed]
- Du, X.Y.; Navdaev, A.; Clemetson, J.M.; Magnenat, E.; Wells, T.N.; Clemetson, K.J. Bilinexin, a snake C-type lectin from Agkistrodon bilineatus venom agglutinates platelets via GPIb and alpha2beta. Thromb. Haemost. 2001, 86, 1277–1283. [Google Scholar] [PubMed]
- Wang, W.J.; Huang, T.F. A novel tetrameric venom protein, agglucetin from Agkistrodon acutus, acts as a glycoprotein Ib agoni. Thromb. Haemost. 2001, 86, 1077–1086. [Google Scholar] [PubMed]
- Xu, G.; Ulrichts, H.; Vauterin, S.; De Meyer, S.F.; Deckmyn, H.; Teng, M.; Niu, L. How does agkicetin-C bind on platelet glycoprotein Ibalpha and achieve its platelet effects? Toxicon 2005, 45, 561–570. [Google Scholar] [PubMed]
- Lu, Q.; Navdaev, A.; Clemetson, J.M.; Clemetson, K.J. GPIb is involved in platelet aggregation induced by mucetin, a snake C-type lectin protein from Chinese habu (Trimeresurus mucrosquamatus) veno. Thromb. Haemost. 2004, 91, 1168–1176. [Google Scholar] [PubMed]
- Huang, K.F.; Ko, T.P.; Hung, C.C.; Chu, J.; Wang, A.H.; Chiou, S.H. Crystal structure of a platelet-agglutinating factor isolated from the venom of Taiwan habu (Trimeresurus mucrosquamatus). Biochem. J. 2004, 378, 399–407. [Google Scholar] [PubMed]
- Wang, W.J.; Ling, Q.D.; Liau, M.Y.; Huang, T.F. A tetrameric glycoprotein Ib-binding protein, agglucetin, from Formosan pit viper: Structure and interaction with human platele. Thromb. Haemost. 2003, 90, 465–475. [Google Scholar] [PubMed]
- Lu, Q.; Clemetson, J.M.; Clemetson, K.J. Translocation of GPIb and Fc receptor gamma-chain to cytoskeleton in mucetin-activated platelets. J. Thromb. Haemost. 2005, 3, 2065–2076. [Google Scholar] [PubMed]
- Peng, M.; Lu, W.; Beviglia, L.; Niewiarowski, S.; Kirby, E.P. Echicetin: A snake venom protein that inhibits binding of von Willebrand factor and alboaggregins to platelet glycoprotein Ib. Blood 1993, 81, 2321–2328. [Google Scholar] [PubMed]
- Taniuchi, Y.; Kawasaki, T.; Fujimura, Y.; Suzuki, M.; Titani, K.; Sakai, Y.; Kaku, S.; Hisamichi, N.; Satoh, N.; Takenaka, T. Flavocetin-A and -B, two high molecular mass glycoprotein Ib binding proteins with high affinity purified from Trimeresurus flavoviridis venom, inhibit platelet aggregation at high shear stres. Biochim. Biophys. Acta 1995, 1244, 331–338. [Google Scholar]
- Kawasaki, T.; Taniuchi, Y.; Hisamichi, N.; Fujimura, Y.; Suzuki, M.; Titani, K.; Sakai, Y.; Kaku, S.; Satoh, N.; Takenaka, T. Tokaracetin, a new platelet antagonist that binds to platelet glycoprotein ib and inhibits von Willebrand factor-dependent shear-induced platelet aggregation. Biochem. J. 1995, 308 (Pt 3), 947–953. [Google Scholar] [PubMed]
- Fujimura, Y.; Ikeda, Y.; Miura, S.; Yoshida, E.; Shima, H.; Nishida, S.; Suzuki, M.; Titani, K.; Taniuchi, Y.; Kawasaki, T. Isolation and characterization of jararaca GPIb-BP, a snake venom antagonist specific to platelet glycoprotein I. Thromb. Haemost. 1995, 74, 743–750. [Google Scholar] [PubMed]
- Zhong, S.R.; Jin, Y.; Wu, J.B.; Chen, R.Q.; Jia, Y.H.; Wang, W.Y.; Xiong, Y.L.; Zhang, Y. Characterization and molecular cloning of dabocetin, a potent antiplatelet C-type lectin-like protein from Daboia russellii siamensis veno. Toxicon 2006, 47, 104–112. [Google Scholar] [PubMed]
- Fukuda, K.; Mizuno, H.; Atoda, H.; Morita, T. Crystal structure of flavocetin-A, a platelet glycoprotein Ib-binding protein, reveals a novel cyclic tetramer of C-type lectin-like heterodime. Biochemistry 2000, 39, 1915–1923. [Google Scholar] [PubMed]
- Jasti, J.; Paramasivam, M.; Srinivasan, A.; Singh, T.P. Crystal structure of echicetin from Echis carinatus (Indian saw-scaled viper) at 2.4A resolution. J. Mol. Biol. 2004, 335, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Huang, Q.; Teng, M.; Liu, P.; Dong, Y.; Niu, L. Crystallization and preliminary X-ray crystallographic analysis of agkicetin-C from Deinagkistrodon acutus venom. Acta Crystallogr. Sect. F. Struct. Biol. Cryst. Commun. 2005, 61, 75–78. [Google Scholar] [PubMed]
- Taniuchi, Y.; Kawasaki, T.; Fujimura, Y. The high molecular mass, glycoprotein Ib-binding protein flavocetin-A induces only small platelet aggregates in vitr. Thromb. Res. 2000, 97, 69–75. [Google Scholar] [PubMed]
- Peng, M.; Holt, J.C.; Niewiarowski, S. Isolation, characterization and amino acid sequence of echicetin beta subunit, a specific inhibitor of von Willebrand factor and thrombin interaction with glycoprotein . Biochem. Biophys. Res. Commun. 1994, 205, 68–72. [Google Scholar] [PubMed]
- Williams, W. Endangered species. CITES puts off plan to hasten shipments. Science 2000, 288, 592. [Google Scholar] [CrossRef] [PubMed]
- Zeymer, U. The role of eptifibatide in patients undergoing percutaneous coronary intervention. Expert Opin. Pharmacother. 2007, 8, 1147–1154. [Google Scholar] [PubMed]
- Hoppe-Seyler, F.; Crnkovic-Mertens, I.; Tomai, E.; Butz, K. Peptide aptamers: Specific inhibitors of protein function. Curr. Mol. Med. 2004, 4, 529–538. [Google Scholar] [PubMed]
- Liu, J.; Joglekar, M.; Ware, J.; Fitzgerald, M.E.; Lowell, C.A.; Berndt, M.C.; Gartner, T.K. Evaluation of the physiological significance of botrocetin/von Willebrand factor in vitro signaling. J. Thromb. Haemost. 2008, 6, 1915–1922. [Google Scholar] [PubMed]
- Liu, J.; Fitzgerald, M.E.; Berndt, M.C.; Jackson, C.W.; Gartner, T.K. Bruton tyrosine kinase is essential for botrocetin/VWF-induced signaling and GPIb-dependent thrombus formation in vivo. Blood 2006, 108, 2596–2603. [Google Scholar] [PubMed]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Matsui, T.; Hamako, J.; Titani, K. Structure and Function of Snake Venom Proteins Affecting Platelet Plug Formation. Toxins 2010, 2, 10-23. https://doi.org/10.3390/toxins2010010
Matsui T, Hamako J, Titani K. Structure and Function of Snake Venom Proteins Affecting Platelet Plug Formation. Toxins. 2010; 2(1):10-23. https://doi.org/10.3390/toxins2010010
Chicago/Turabian StyleMatsui, Taei, Jiharu Hamako, and Koiti Titani. 2010. "Structure and Function of Snake Venom Proteins Affecting Platelet Plug Formation" Toxins 2, no. 1: 10-23. https://doi.org/10.3390/toxins2010010
APA StyleMatsui, T., Hamako, J., & Titani, K. (2010). Structure and Function of Snake Venom Proteins Affecting Platelet Plug Formation. Toxins, 2(1), 10-23. https://doi.org/10.3390/toxins2010010