ADAM-15 Disintegrin-Like Domain Structure and Function

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
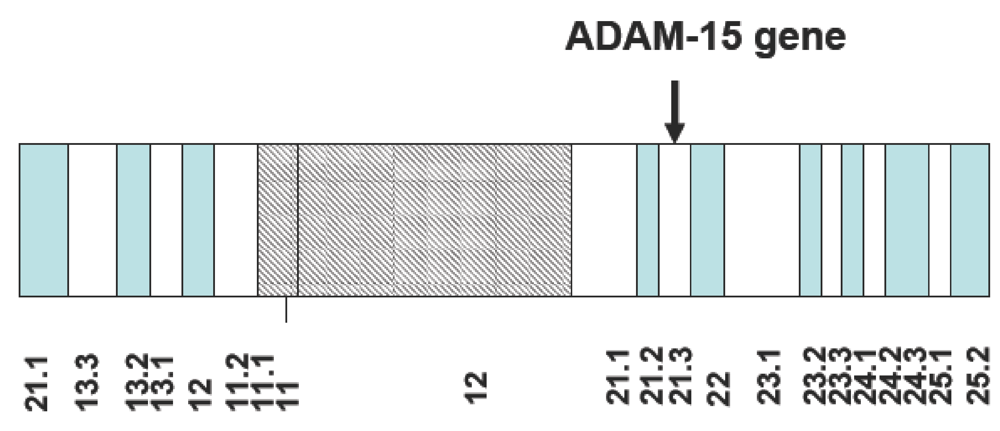
2. Overview of the ADAM-15 Gene Structure


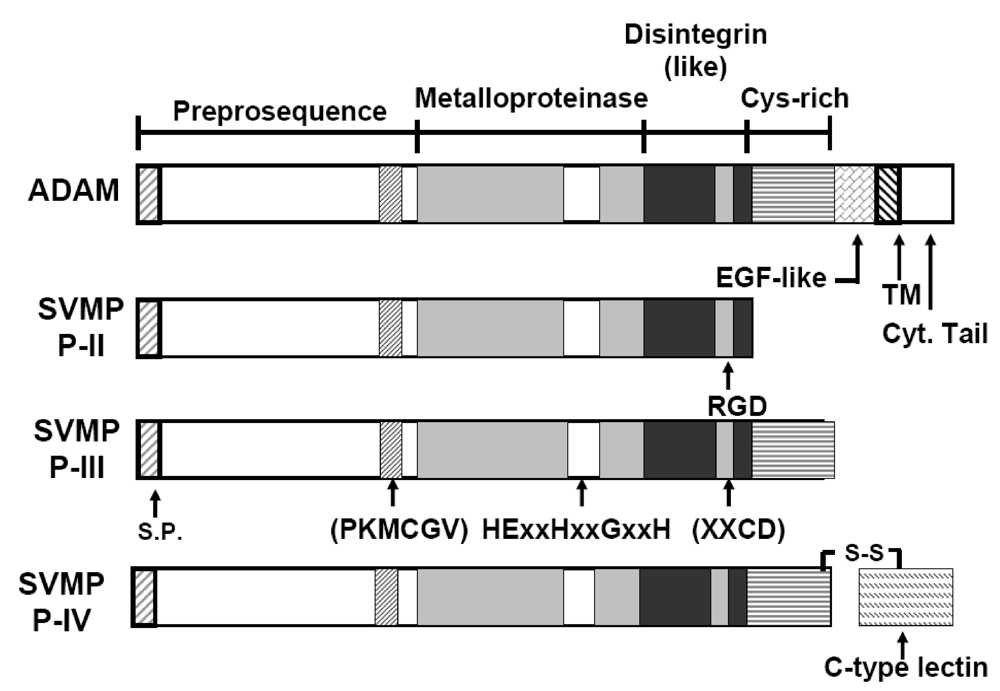
3. Overview of the Structural Domains of ADAM 15
4. Integrin Interactions of the Disintegrin-Like Domain of ADAM-15
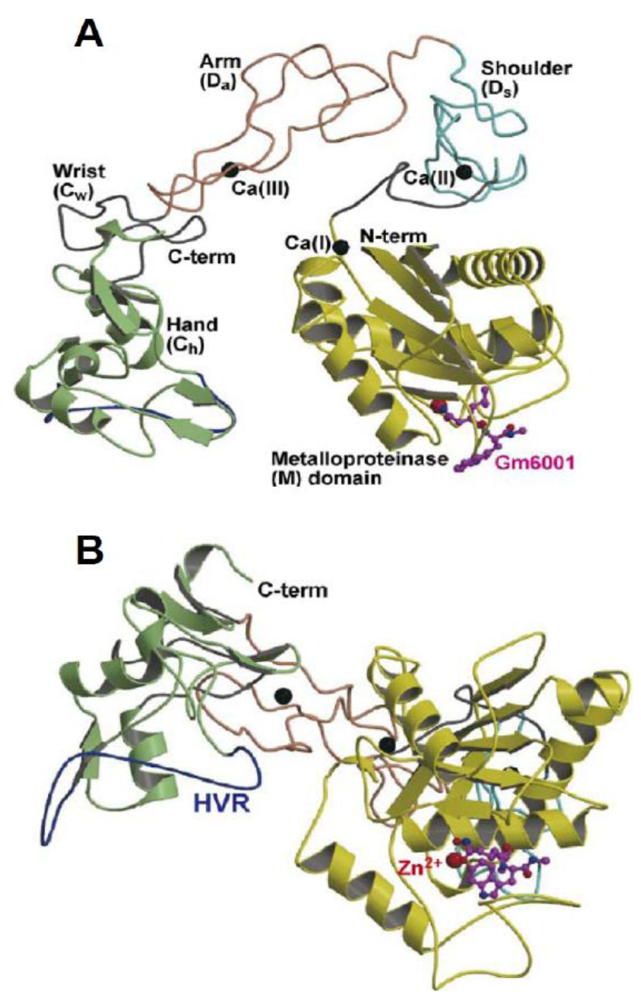
5. Structural Model of the Disintegrin-Like Domains of ADAMs Proteins

6. Disintegrin-Like Domain of ADAM-15: Structure and Function
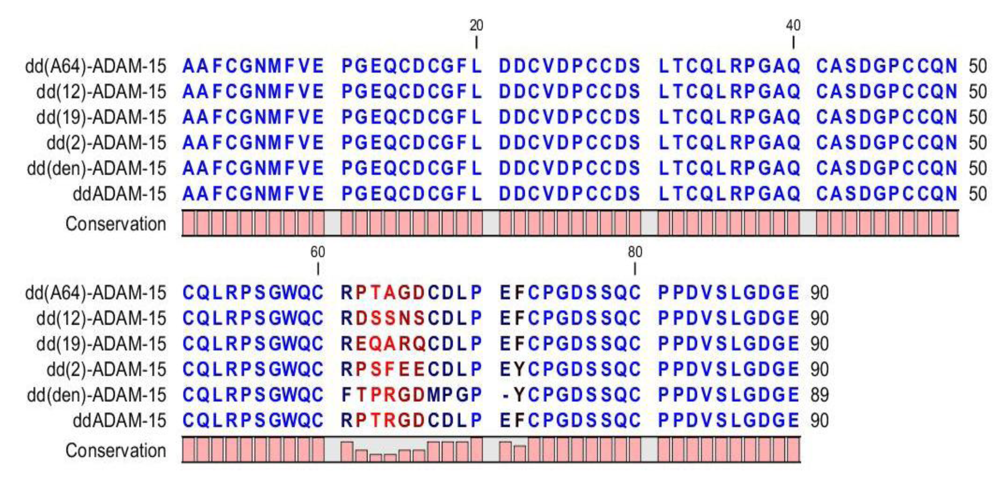
7. Investigation of the Integrin Interaction Using a Recombinant Disintegrin Domain from ADAM‑15 (ddADAM-15) and Various Mutants

8. ADAM-15 Is Associated with Diseases
9. Concluding Remarks
Acknowledgements
References
- Gould, R.J.; Polokoff, M.A.; Friedman, P.A.; Huang, T.F.; Holt, J.C.; Cook, J.J.; Niewiarowski, S. Disintegrins: a family of integrin inhibitory proteins from viper venoms. Proc. Soc. Exp. Biol. Med. 1990, 195, 168–171. [Google Scholar]
- Niewiarowski, S.; McLane, M.A.; Kloczewiak, M.; Stewart, G.J. Disintegrins and other naturally occurring antagonists of platelet fibrinogen receptors. Semin. Hematol. 1994, 31, 289–300. [Google Scholar]
- McLane, M.A.; Sanchez, E.E.; Wong, A.; Paquette-Straub, C.; Perez, J.C. Disintegrins. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2004, 4, 327–355. [Google Scholar] [CrossRef] [PubMed]
- Scarborough, R.M.; Rose, J.W.; Hsu, M.A.; Phillips, D.R.; Fried, V.A.; Campbell, A.M.; Nannizzi, L.; Charo, I.F. Barbourin. A GPIIb-IIIa-specific integrin antagonist from the venom of Sistrurus m. barbouri. J. Biol. Chem. 1991, 266, 9359–9362. [Google Scholar] [PubMed]
- Oshikawa, K.; Terada, S. Ussuristatin 2, a novel KGD-bearing disintegrin from Agkistrodon ussuriensis venom. J. Biochem. 1999, 125, 31–35. [Google Scholar]
- Nikai, T.; Taniguchi, K.; Komori, Y.; Masuda, K.; Fox, J.W.; Sugihara, H. Primary structure and functional characterization of bilitoxin-1, a novel dimeric P-II snake venom metalloproteinase from Agkistrodon bilineatus venom. Arch. Biochem. Biophys. 2000, 378, 6–15. [Google Scholar]
- Calvete, J.J.; Fox, J.W.; Agelan, A.; Niewiarowski, S.; Marcinkiewicz, C. The presence of the WGD motif in CC8 heterodimeric disintegrin increases its inhibitory effect on alphaII(b)beta 3, alpha (v)beta3, and alpha5beta1 integrins. Biochemistry 2002, 41, 2014–2021. [Google Scholar]
- Siigur, E.; Aaspollu, A.; Tu, A.T.; Siigur, J. cDNA cloning and deduced amino acid sequence of fibrinolytic enzyme (lebetase) from Vipera lebetina snake venom. Biochem. Biophys. Res. Commun. 1996, 224, 229–236. [Google Scholar]
- Marcinkiewicz, C.; Calvete, J.J.; Marcinkiewicz, M.M.; Raida, M.; Vijay-Kumar, S.; Huang, Z.; Lobb, R.R.; Niewiarowski, S. EC3, a novel heterodimeric disintegrin from Echis carinatus venom, inhibits alpha4 and alpha5 integrins in an RGD-independent manner. J. Biol. Chem. 1999, 274, 12468–12473. [Google Scholar]
- Seymour, J.L.; Henzel, W.J.; Nevins, B.; Stults, J.T.; Lazarus, R.A. Decorsin. A potent glycoprotein IIb-IIIa antagonist and platelet aggregation inhibitor from the leech Macrobdella decora. J. Biol. Chem. 1990, 265, 10143–10147. [Google Scholar] [PubMed]
- Mazur, P.; Henzel, W.J.; Seymour, J.L.; Lazarus, R.A. Ornatins: potent glycoprotein IIb-IIIa antagonists and platelet aggregation inhibitors from the leech Placobdella ornata. Eur. J. Biochem. 1991, 202, 1073–1082. [Google Scholar]
- Wang, X.; Coons, L.B.; Taylor, D.B.; Stevens, S.E., Jr.; Gartner, T.K. Variabilin, a novel RGD-containing antagonist of glycoprotein IIb-IIIa and platelet aggregation inhibitor from the hard tick Dermacentor variabilis. J. Biol. Chem. 1996, 271, 17785–17790. [Google Scholar]
- Bjarnason, J.B.; Fox, J.W. Hemorrhagic metalloproteinases from snake venoms. Pharmacol. Ther. 1994, 62, 325–372. [Google Scholar]
- Lu, X.; Lu, D.; Scully, M.F.; Kakkar, V.V. Snake venom metalloproteinase containing a disintegrin-like domain, its structure-activity relationships at interacting with integrins. Curr. Med. Chem. Cardiovasc. Hematol. Agents 2005, 3, 249–260. [Google Scholar]
- Weskamp, G.; Blobel, C.P. A family of cellular proteins related to snake venom disintegrins. Proc. Natl. Acad. Sci. USA 1994, 91, 2748–2751. [Google Scholar]
- Wolfsberg, T.G.; Primakoff, P.; Myles, D.G.; White, J.M. ADAM, a novel family of membrane proteins containing A Disintegrin and Metalloproteinase domain: multipotential functions in cell-cell and cell-matrix interactions. J. Cell Biol. 1995, 131, 275–278. [Google Scholar]
- Wolfsberg, T.G.; Straight, P.D.; Gerena, R.L.; Huovila, A.P.; Primakoff, P.; Myles, D.G.; White, J.M. ADAM, a widely distributed and developmentally regulated gene family encoding membrane proteins with a disintegrin and metalloproteinase domain. Dev. Biol. 1995, 169, 378–383. [Google Scholar]
- Wolfsberg, T.G.; White, J.M. ADAMs in fertilization and development. Dev. Biol. 1996, 180, 389–401. [Google Scholar]
- Kuno, K.; Kanada, N.; Nakashima, E.; Fujiki, F.; Ichimura, F.; Matsushima, K. Molecular cloning of a gene encoding a new type of metalloproteinase-disintegrin family protein with thrombospondin motifs as an inflammation associated gene. J. Biol. Chem. 1997, 272, 556–562. [Google Scholar]
- Tang, B.L.; Hong, W. ADAMTS: a novel family of proteases with an ADAM protease domain and thrombospondin 1 repeats. FEBS Lett. 1999, 445, 223–225. [Google Scholar]
- Hirohata, S.; Wang, L.W.; Miyagi, M.; Yan, L.; Seldin, M.F.; Keene, D.R.; Crabb, J.W.; Apte, S.S. Punctin, a novel ADAMTS-like molecule, ADAMTSL-1, in extracellular matrix. J. Biol. Chem. 2002, 277, 12182–12189. [Google Scholar]
- Perry, A.C.; Jones, R.; Hall, L. Analysis of transcripts encoding novel members of the mammalian metalloproteinase-like, disintegrin-like, cysteine-rich (MDC) protein family and their expression in reproductive and non-reproductive monkey tissues. Biochem. J. 1995, 312, 239–244. [Google Scholar]
- Selistre de Araujo, H.S.; de Souza, D.H.; Ownby, C.L. Analysis of a cDNA sequence encoding a novel member of the snake venom metalloproteinase, disintegrin-like, cysteine-rich (MDC) protein family from Agkistrodon contortrix laticinctus. Biochim. Biophys. Acta 1997, 1342, 109–115. [Google Scholar]
- Lum, L.; Reid, M.S.; Blobel, C.P. Intracellular maturation of the mouse metalloproteinase disintegrin MDC15. J. Biol. Chem. 1998, 273, 26236–26247. [Google Scholar]
- Herren, B.; Raines, E.; Ross, R. Expression of a disintegrin-like protein in cultured human vascular cells and in vivo. FASEB J. 1997, 11, 173–180. [Google Scholar]
- Kratzschmar, J.; Lum, L.; Blobel, C.P. Metargidin, a membrane-anchored metalloproteinase-disintegrin protein with an RGD integrin binding sequence. J. Biol. Chem. 1996, 271, 4593–4596. [Google Scholar]
- Kärkkäinen, I.; Karhu, R.; Huovila, A.P. Assignment of the ADAM15 gene to human chromosome band 1q21.3 by in situ hybridization. Cytogenet. Cell Genet. 2000, 88, 206–207. [Google Scholar] [CrossRef] [PubMed]
- Kleino, I.; Ortiz, R.M.; Huovila, A.P. ADAM15 gene structure and differential alternative exon use in human tissues. BMC Mol. Biol. 2007, 8, 90–119. [Google Scholar]
- Kleino, I.; Ortiz, R.M.; Yritys, M.; Huovila, A.P.; Saksela, K. Alternative splicing of ADAM15 regulates its interactions with cellular SH3 proteins. Cell Biochem. 2009, 108, 877–885. [Google Scholar]
- Hurskainen, T.L.; Hirohata, S.; Seldin, M.F.; Apte, S.S. ADAM-TS5, ADAM-TS6, and ADAM-TS7, novel members of a new family of zinc metalloproteinases. General features and genomic distribution of the ADAM-TS family. J. Biol. Chem. 1999, 274, 25555–25563. [Google Scholar] [PubMed]
- Becker, J.W.; Marcy, A.I.; Rokosz, L.L.; Axel, M.G.; Burbaum, J.J.; Fitzgerald, P.M.; Cameron, P.M.; Esser, C.K.; Hagmann, W.K.; Hermes, J.D.; Springer, J.P. Stromelysin-1: three-dimensional structure of the inhibited catalytic domain and of the C-truncated proenzyme. Protein Sci. 1995, 4, 1966–1976. [Google Scholar]
- Kang, T.; Zhao, Y.G.; Pei, D.; Sucic, J.F.; Sang, Q.X. Intracellular activation of human adamalysin 19/disintegrin and metalloproteinase 19 by furin occurs via one of the two consecutive recognition sites. J. Biol. Chem. 2002, 277, 25583–25592. [Google Scholar]
- Anders, A.; Gilbert, S.; Garten, W.; Postina, R.; Fahrenholz, F. Regulation of the alpha-secretase ADAM10 by its prodomain and proprotein convertases. FASEB J. 2001, 15, 1837–1879. [Google Scholar]
- Fox, J.W.; Bjarnason, J.B. Zinc Metalloproteinases in Health and Disease; Hooper, N.M., Ed.; Taylor & Francis: Bristol, UK, 1996; pp. 47–81. [Google Scholar]
- Marcinkiewicz, C. Functional characteristic of snake venom disintegrins: potential therapeutic implication. Curr. Pharm. Des. 2005, 11, 815–827. [Google Scholar]
- Loechel, F.; Gilpin, B.J.; Engvall, E.; Albrechtsen, R.; Wewer, U.M. Human ADAM-12 (meltrin α) is an active metalloproteinase. J. Biol. Chem. 1998, 273, 16993–16997. [Google Scholar]
- Wolfsberg, T.G.; Bazan, J.F.; Blobel, C.P.; Myles, D.G.; Primakoff, P.; White, J.M. The precursor region of a protein active in sperm-egg fusion contains a metalloproteinase and a disintegrin domain: structural, functional, and evolutionary implications. Proc. Natl. Acad. Sci. USA 1993, 90, 10783–10787. [Google Scholar]
- Smith, K.M.; Gaultier, A.; Cousin, H.; Alfandari, D.; White, J.M.; DeSimone, D.W. The cysteine-rich domain regulates ADAM protease function in vivo. J. Cell Biol. 2002, 159, 893–902. [Google Scholar]
- Zhong, J.L.; Poghosyan, Z.; Pennington, C.J.; Scott, X.; Handsley, M.M.; Warn, A.; Gavrilovic, J.; Honert, K.; Krüger, A.; Span, P.N.; et al. Distinct Functions of Natural ADAM-15 Cytoplasmic Domain Variants in Human Mammary Carcinoma. Mol. Cancer Res. 2008, 6, 383–394. [Google Scholar] [CrossRef] [PubMed]
- White, J.M.; Bridges, L.C.; DeSimone, D.W.; Tomczuk, M.; Wolfsberg, T.G. Introduction to the ADAM family. In The ADAM Family of Proteases: Proteases in Biology and Disease; Hooper, N.M., Lendeckel, V.D., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 4, pp. 1–28. [Google Scholar]
- Langer, H.; May, A.E.; Bültmann, A.; Gawaz, M. ADAM 15 is an adhesion receptor for platelet GPIIb-IIIa and induces platelet activation. Thromb. Haemost. 2005, 94, 555–561. [Google Scholar]
- Eto, K.; Huet, C.; Tarui, T.; Kupriyanov, S.; Liu, H.Z.; Puzon-McLaughlin, W.; Zhang, X.P.; Sheppard, D.; Engvall, E.; Takada, Y. Functional classification of ADAMs based on a conserved motif for binding to integrin alpha 9beta 1: implications for sperm-egg binding and other cell interactions. J. Biol. Chem. 2002, 277, 17804–17810. [Google Scholar]
- Tomczuk, M.; Takahashi, Y.; Huang, J.; Murase, S.; Mistretta, M.; Klaffky, E.; Sutherland, A.; Bolling, L.; Coonrod, S.; Marcinkiewicz, C.; et al. Role of multiple beta1 integrins in cell adhesion to the disintegrin domains of ADAMs 2 and 3. Exp. Cell Res. 2003, 290, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Lu, D.; Scully, M.; Kakkar, V. ADAM proteins - therapeutic potential in cancer. Curr. Cancer Drug Targets 2008, 8, 720–732. [Google Scholar]
- Adler, M.; Lazarus, R.A.; Dennis, M.S.; Wagner, G. Solution structure of kistrin, a potent platelet aggregation inhibitor and GP IIb-IIIa antagonist. Science 1991, 253, 445–448. [Google Scholar]
- Adler, M.; Wagner, G. Sequential 1H NMR assignments of kistrin, a potent platelet aggregation inhibitor and glycoprotein IIb-IIIa antagonist. Biochemistry 1992, 31, 1031–1039. [Google Scholar]
- Adler, M.; Carter, P.; Lazarus, R.A.; Wagner, G. Cysteine pairing in the glycoprotein IIb-IIIa antagonist kistrin using NMR, chemical analysis, and structure calculations. Biochemistry 1993, 32, 282–289. [Google Scholar]
- Senn, H.; Klaus, W. The nuclear magnetic resonance solution structure of flavoridin, an antagonist of the platelet GP IIb-IIIa receptor. J. Mol. Biol. 1993, 232, 907–925. [Google Scholar] [CrossRef] [PubMed]
- Saudek, V.; Atkinson, R.A.; Lepage, P.; Pelton, J.T. The secondary structure of echistatin from 1H-NMR, circular-dichroism and Raman spectroscopy. Eur. J. Biochem. 1991, 202, 329–338. [Google Scholar]
- Cooke, R.M.; Carter, B.G.; Martin, D.M.A.; Murray-Rust, P.; Weir, M.P. Nuclear magnetic resonance studies of the snake toxin echistatin. 1H resonance assignments and secondary structure. Eur. J. Biochem. 1991, 202, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Cooke, R.M.; Carter, B.G.; Murray-Rust, P.; Hartshorn, M.J.; Herzyk, P.; Hubbard, R.E. The solution structure of echistatin: evidence for disulphide bond rearrangement in homologous snake toxins. Protein Eng. 1992, 5, 473–477. [Google Scholar]
- Jaseja, M.; Smith, K.J.; Lu, X.; Williams, J.A.; Trayer, H.; Trayer, I.P.; Hyde, E.I. 1H-NMR studies and secondary structure of the RGD-containing snake toxin, albolabrin. Eur. J. Biochem. 1993, 218, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Jaseja, M.; Lu, X.; Williams, J.A.; Sutcliffe, M.J.; Kakkar, V.V.; Parslow, R.A.; Hyde, E.I. 1H-NMR assignments and secondary structure of dendroaspin, an RGD-containing glycoprotein IIb-IIIa (alpha IIb-beta 3) antagonist with a neurotoxin fold. Eur. J. Biochem. 1994, 226, 861–868. [Google Scholar]
- Fujii, Y.; Okuda, D.; Fujimoto, Z.; Horii, K.; Morita, T.; Mizuno, H. Crystal structure of trimestatin, a disintegrin containing a cell adhesion recognition motif RGD. J. Mol. Biol. 2003, 332, 1115–1122. [Google Scholar]
- McLane, M.A.; Marcinkiewicz, C.; Vijay-Kumar, S.; Wierzbicka-Patynowski, I.; Niewiarowski, S. Viper venom disintegrins and related molecules. Proc. Soc. Exp. Biol. Med. 1998, 219, 109–119. [Google Scholar]
- Lu, X.; Lu, D.; Scully, M.F.; Kakkar, V.V. Structure-activity relationship studies on ADAM protein-integrin interactions. Cardiovasc. Hematol. Agents Med. Chem. 2007, 5, 29–42. [Google Scholar]
- Gomis-Ruth, F.X. Structural aspects of the metzincin clan of metalloendopeptidases. Mol. Biotechnol. 2003, 24, 157–202. [Google Scholar]
- White, J.M. ADAMs: modulators of cell–cell and cel–matrix interactions. Curr. Opin. Cell Biol. 2003, 15, 598–606. [Google Scholar]
- Terra, R.M.; Pinto, A.F.; Guimarães, J.A.; Fox, J.W. Proteomic profiling of snake venom metalloproteinases (SVMPs): insights into venom induced pathology. Toxicon 2009, 54, 836–844. [Google Scholar]
- Baldo, C.; Tanjoni, I.; León, I.R.; Batista, I.F.; Della-Casa, M.S.; Clissa, P.B.; Weinlich, R.; Lopes-Ferreira, M.; Lebrun, I.; Amarante-Mendes, G.P.; et al. BnP1, a novel P-I metalloproteinase from Bothrops neuwiedi venom: biological effects benchmarking relatively to jararhagin, a P-III SVMP. Toxicon 2008, 51, 54–65. [Google Scholar] [PubMed]
- Guan, H.H.; Goh, K.S.; Davamani, F.; Wu, P.L.; Huang, Y.W.; Jeyakanthan, J.; Wu, W.G.; Chen, C.J. Structures of two elapid snake venom metalloproteases with distinct activities highlight the disulfide patterns in the D domain of ADAMalysin family proteins. J. Struct. Biol. 2010, 169, 294–303. [Google Scholar]
- Liu, H.; Shim, A.H.; He, X. Structural characterization of the ectodomain of a disintegrin and metalloproteinase-22 (ADAM22), a neural adhesion receptor instead of metalloproteinase: insights on ADAM function. J. Biol. Chem. 2009, 284, 29077–29086. [Google Scholar]
- Takeda, S.; Igarashi, T.; Mori, H.; Araki, S. Crystal structures of VAP1 reveal ADAMs’ MDC domain architecture and its unique C-shaped scaffold. EMBO J. 2006, 25, 2388–2396. [Google Scholar]
- Igarashi, T.; Araki, S.; Mori, H.; Takeda, S. Crystal structures of catrocollastatin/VAP2B reveal a dynamic, modular architecture of ADAM/adamalysin/reprolysin family proteins. FEBS Lett. 2007, 581, 2416–2422. [Google Scholar]
- Takeda, S. VAP1: snake venom homolog of mammalian ADAMs. In Handbook of Metalloproteins; Messerschmidt, A., Ed.; John Wiley & Sons, Inc.: Somerset, NJ, USA, 2008. [Google Scholar]
- Takeda, S. Three-dimensional domain architecture of the ADAM family proteinases. Semin. Cell Dev. Biol. 2009, 20, 146–152. [Google Scholar]
- Nath, D.; Slocombe, P.M.; Stephens, P.E.; Warn, A.; Hutchinson, G.R.; Yamada, K.M.; Docherty, A.J.; Murphy, G. Interaction of metargidin (ADAM-15) with alphavbeta3 and alpha5beta1 integrins on different haemopoietic cells. J. Cell Sci. 1999, 112, 579–587. [Google Scholar]
- Eto, K.; Puzon-McLaughlin, W.; Sheppard, D.; Sehara-Fujisawa, A.; Zhang, X.P.; Takada, Y. RGD-independent binding of integrin alpha9beta1 to the ADAM-12 and -15 disintegrin domains mediates cell-cell interaction. J. Biol. Chem. 2000, 275, 34922–34930. [Google Scholar]
- Fourie, A.M.; Coles, F.; Moreno, V.; Karlsson, L. Catalytic activity of ADAM8, ADAM15, and MDC-L (ADAM28) on synthetic peptide substrates and in ectodomain cleavage of CD23. J. Biol. Chem. 2003, 278, 30469–30477. [Google Scholar]
- Seals, D.F.; Courtneidge, S.A. The ADAMs family of metalloproteinases: multidomain proteins with multiple functions. Genes Dev. 2003, 17, 7–30. [Google Scholar]
- Taooka, Y.; Chen, J.; Yednock, T.; Sheppard, D. The integrin alpha9beta1 mediates adhesion to activated endothelial cells and transendothelial neutrophil migration through interaction with vascular cell adhesion molecule-1. J. Cell Biol. 1999, 145, 413–420. [Google Scholar]
- Kanayama, M.; Kurotaki, D.; Morimoto, J.; Asano, T.; Matsui, Y.; Nakayama, Y.; Saito, Y.; Ito, K.; Kimura, C.; Iwasaki, N.; et al. Alpha9 integrin and Its Ligands Constitute Critical Joint Microenvironments for Development of Autoimmune Arthritis. J. Immunol. 2009, 182, 8015–8025. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Chung, K.F.; Xia, M.; Lu, X.; Scully, M.; Kakkar, V. Integrin binding characteristics of the disintegrin-like domain of ADAM-15. Thromb. Haemost. 2006, 96, 642–651. [Google Scholar]
- Charrier, L.; Yan, Y.; Nguyen, H.T.; Dalmasso, G.; Laboisse, C.L.; Gewirtz, A.T.; Sitaraman, S.V.; Merlin, D. ADAM-15/metargidin mediates homotypic aggregation of human T lymphocytes and heterotypic interactions of T lymphocytes with intestinal epithelial cells. J. Biol. Chem. 2007, 282, 16948–16958. [Google Scholar]
- Zhang, X.P.; Kamata, T.; Yokoyama, K.; Puzon-McLaughlin, W.; Takada, Y. Specific interaction of the recombinant disintegrin-like domain of MDC-15 (metargidin, ADAM-15) with integrin alphavbeta3. J. Biol. Chem. 1998, 273, 7345–7350. [Google Scholar]
- Mosnier, J.F.; Jarry, A.; Bou-Hanna, C.; Denis, M.G.; Merlin, D.; Laboisse, C.L. ADAM15 upregulation and interaction with multiple binding partners in inflammatory bowel disease. Lab. Invest. 2006, 86, 1064–1073. [Google Scholar]
- Eliceiri, B.P.; Cheresh, D.A. Role of alpha v integrins during angiogenesis. Cancer J. 2000, 3, S245–S249. [Google Scholar]
- Kim, J.B.; Islam, S.; Kim, Y.J.; Prudoff, R.S.; Sass, K.M.; Wheelock, M.J.; Johnson, K.R. N-Cadherin extracellular repeat 4 mediates epithelial to mesenchymal transition and increased motility. J. Cell Biol. 2000, 151, 1193–1206. [Google Scholar]
- Jeon, O.H.; Kim, D.; Choi, Y.J.; Kim, S.H.; Choi, W.S.; Kim, D.S. Novel function of human ADAM15 disintegrin-like domain and its derivatives in platelet aggregation. Thromb. Res. 2007, 119, 609–619. [Google Scholar]
- Lu, D.; Xie, S.; Sukkar, M.B.; Lu, X.; Scully, M.F.; Chung, K.F. Inhibition of airway smooth muscle adhesion and migration by the disintegrin domain of ADAM-15. Am. J. Respir. Cell Mol. Biol. 2007, 37, 494–500. [Google Scholar]
- Pierschbacher, M.D.; Ruoslahti, E. Variants of the cell recognition site of fibronectin that retain attachment-promoting activity. Proc. Natl. Acad. Sci. USA 1984, 81, 5985–5988. [Google Scholar]
- Mould, A.P.; Burrows, L.; Humphries, M.J. Identification of amino acid residues that form part of the ligand-binding pocket of integrin alpha5 beta1. J. Biol. Chem. 1998, 273, 25664–25672. [Google Scholar]
- Murphy, G. The ADAMs: Signalling scissors in the tumour microenvironment. Nat. Rev. Cancer 2008, 8, 929–941. [Google Scholar]
- Horiuchi, K,; Weskamp, G.; Lum, L.; Hammes, H.P.; Cai, H.; Brodie, T.A.; Ludwig, T.; Chiusaroli, R.; Baron, R.; Preissner, K.T.; et al. Potential role for ADAM15 in pathological neovascularization in mice. Mol. Cell Biol. 2003, 23, 5614–5624. [Google Scholar] [CrossRef] [PubMed]
- Yamada, D.; Ohuchida, K.; Mizumoto, K.; Ohhashi, S.; Yu, J.; Egami, T.; Fujita, H.; Nagai, E.; Tanaka, M. Increased expression of ADAM 9 and ADAM 15 mRNA in pancreatic cancer. Anticancer Res. 2007, 27, 793–799. [Google Scholar]
- Najy, A.J.; Day, K.C.; Day, M.L. ADAM15 supports prostate cancer metastasis by modulating tumor cell-endothelial cell interaction. Cancer Res. 2008, 68, 1092–1099. [Google Scholar]
- Najy, A.J.; Day, K.C.; Day, M.L. The ectodomain shedding of E-cadherin by ADAM15 supports ErbB receptor activation. J. Biol. Chem. 2008, 283, 18393–18401. [Google Scholar]
- Ross, R. Atherosclerosis-an inflammatory disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar]
- Herren, B.; Raines, E.W.; Ross, R. Expression of a disintegrin-like protein in cultured human vascular cells and in vivo. FASEB J. 1997, 11, 173–180. [Google Scholar]
- Yuan, S.Y.; Ustinova, E.E.; Wu, M.H.; Tinsley, J.H.; Xu, W.; Korompai, F.L.; Taulman, A.C. Protein kinase C activation contributes to microvascular barrier dysfunction in the heart at early stages of diabetes. Circ. Res. 2000, 87, 412–417. [Google Scholar]
- Matsuki, K.; Sasho, T.; Nakagawa, K.; Tahara, M.; Sugioka, K.; Ochiai, N.; Ogino, S.; Wada, Y.; Moriya, H. RGD peptide-induced cell death of chondrocytes and synovial cells. J. Orthop. Sci. 2008, 13, 524–532. [Google Scholar]
- Zhou, Q.; Nakada, M.T.; Arnold, C.; Shieh, K.Y.; Markland, F.S., Jr. Contortrostatin, a dimeric disintegrin from Agkistrodon contortrix contortrix, inhibits angiogenesis. Angiogenesis 1999, 3, 259–269. [Google Scholar]
- Wu, W.B.; Peng, H.C.; Huang, T.F. Disintegrin causes proteolysis of beta-catenin and apoptosis of endothelial cells. Involvement of cellcell and cell-ECM interactions in regulating cell viability. Exp. Cell Res. 2003, 286, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.H.; Kim, S.H.; Han, K.Y.; Sohn, Y.D.; Chang, S.I.; Baek, K.H.; Jang, Y.; Kim, D.S.; Kang, I.C. Inhibitory effect of salmosin, a Korean snake venom-derived disintegrin, on the integrin alphav-mediated proliferation of SK-Mel-2 human melanoma cells. J. Pharm. Pharmacol. 2003, 55, 1577–1582. [Google Scholar]
- Kerr, J.S.; Slee, A.M.; Mousa, S.A. The alpha v integrin antagonists as novel anticancer agents: an update. Expert Opin. Investig. Drugs 2002, 11, 1765–1774. [Google Scholar]
- Hallahan, D.E.; Qu, S.; Geng, L.; Cmelak, A.; Chakravarthy, A.; Martin, W.; Scarfone, C.; Giorgio, T. Radiation-mediated control of drug delivery. Am. J. Clin. Oncol. 2001, 24, 473–480. [Google Scholar]
- Coller, B.S. Anti-GPIIb/IIIa drugs: current strategies and future directions. Thromb. Haemost. 2001, 86, 427–443. [Google Scholar]
- Xiao, T.; Takagi, J.; Coller, B.S.; Wang, J.H.; Springer, T.A. Structural basis for allostery in integrins and binding to fibrinogen-mimetic therapeutics. Nature 2004, 432, 59–67. [Google Scholar]
- Pinto, A.F.; Terra, R.M.; Guimaraes, J.A.; Fox, J.W. Mapping von Willebrand factor A domain binding sites on a snake venom metalloproteinase cysteinerich domain. Arch. Biochem. Biophys. 2007, 457, 41–46. [Google Scholar]
- Cal, S.; Freije, J.M.; López, J.M.; Takada, Y.; López-Otín, C. ADAM 23/MDC3, a human disintegrin that promotes cell adhesion via interaction with the alphavbeta3 integrin through an RGD-independent mechanism. Mol. Biol. Cell 2000, 11, 1457–1469. [Google Scholar]
- Bridges, L.C.; Tani, P.H.; Hanson, K.R.; Roberts, C.M.; Judkins, M.B.; Bowditch, R.D. The lymphocyte metalloprotease MDC-L (ADAM 28) is a ligand for the integrin α4β1. J. Biol. Chem. 2002, 277, 3784–3792. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lu, D.; Scully, M.; Kakkar, V.; Lu, X. ADAM-15 Disintegrin-Like Domain Structure and Function. Toxins 2010, 2, 2411-2427. https://doi.org/10.3390/toxins2102411
Lu D, Scully M, Kakkar V, Lu X. ADAM-15 Disintegrin-Like Domain Structure and Function. Toxins. 2010; 2(10):2411-2427. https://doi.org/10.3390/toxins2102411
Chicago/Turabian StyleLu, Dong, Mike Scully, Vijay Kakkar, and Xinjie Lu. 2010. "ADAM-15 Disintegrin-Like Domain Structure and Function" Toxins 2, no. 10: 2411-2427. https://doi.org/10.3390/toxins2102411
APA StyleLu, D., Scully, M., Kakkar, V., & Lu, X. (2010). ADAM-15 Disintegrin-Like Domain Structure and Function. Toxins, 2(10), 2411-2427. https://doi.org/10.3390/toxins2102411