Biological Profile of Erucin: A New Promising Anticancer Agent from Cruciferous Vegetables

Abstract
:1. Introduction
2. Pharmacokinetic and Bioavailability of ER
3. Bioactivity of Rocket salad Species


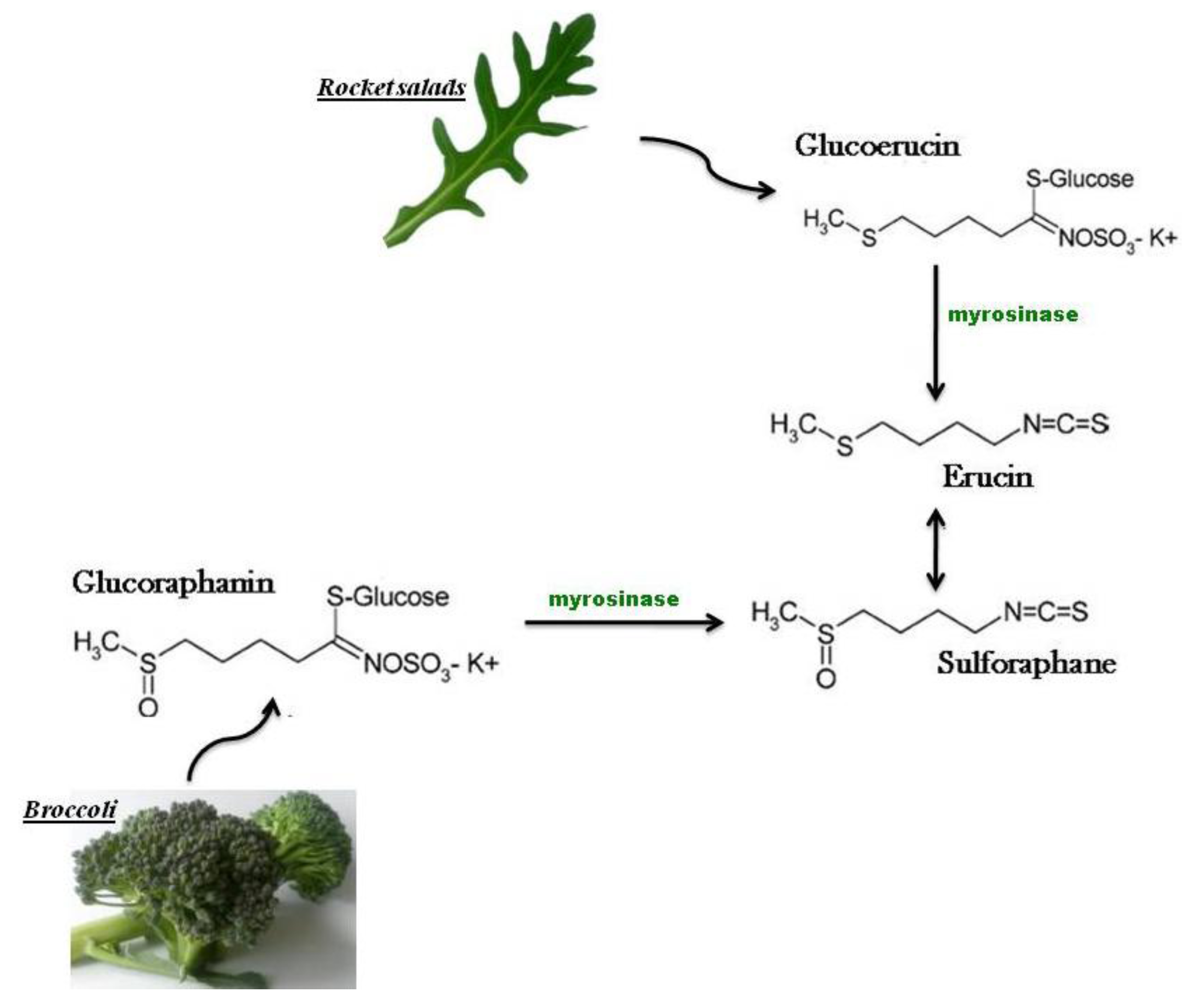{kind=link}
{kind=link}
{kind=link}
| Rocket species | Health promoting activity | Experimental Model | Reference |
|---|---|---|---|
| Eruca sativa (seeds) | antidiabetic activity | rats | [48] |
| Eruca sativa (seeds) | protective effect against HgCl2-induced nephrotoxicity | rats | [49] |
| Eruca sativa (seeds, sprouts) | antioxidant activity | in vitro assays | [50] |
| Eruca sativa (leaves) | [51] | ||
| Eruca sativa (leaves) | [52] | ||
| Eruca sativa (leaves) | anti- ulcer activity | albino rats | [53] |
| Eruca sativa | antigenotoxic activity | human hepatocellular carcinoma HepG2 cell line | [77] |
| Eruca sativa (leaves) | chemopreventive activity | human colonic cancer HT-29 cell line | [55] |
| Diplotaxis tenuifolia (leaves) |
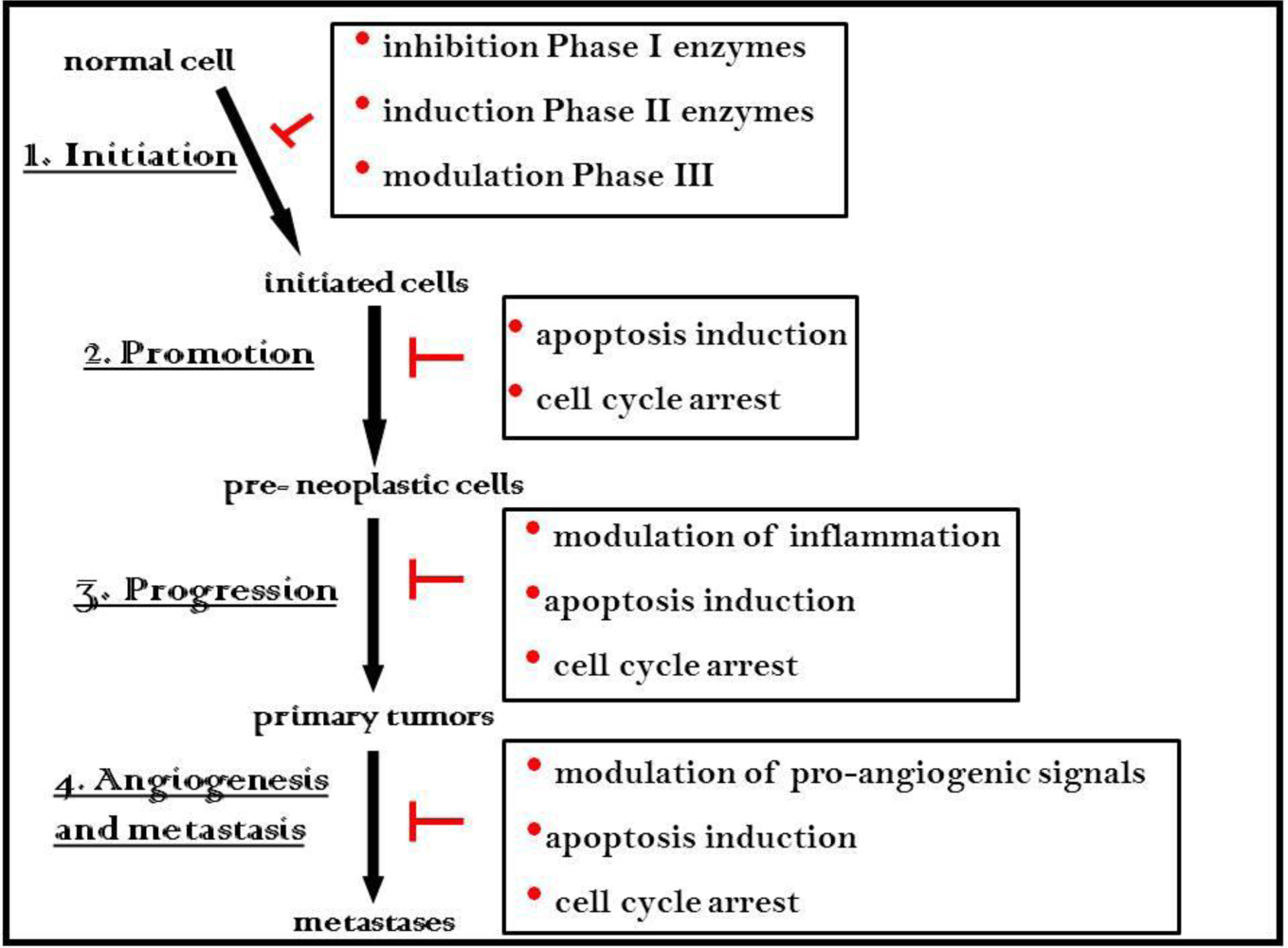
4. Cancer Chemoprevention and Treatment with ER: Evidence from Cell and Animal Research
5. Molecular Mechanisms of Cancer Chemoprevention by ER

| Biological activity | Molecular targets | Experimental model | Reference |
|---|---|---|---|
| Modulation of Phase I enzymes | CYP540 isoforms | human hepatocellular carcinoma HepG2 cell line | [77] |
| ex vivo rat and human tissues | [57,58] | ||
| Induction of Phase II enzymes | quinone reductase (QR) glutatione transferase (GST) | mouse tissues | [56] |
| human tissues | [61] | ||
| human colonic cancer CACO-2 cells | [62] | ||
| rat and human tissues | [59] | ||
| Up- regulation of Phase III detoxification system | multidrug resistance proteins (MRP- 1 and 2) | human colonic cancer CACO-2 cells | [61] |
| human cancer cell lines (HepG2, CACO-2, A549) | [91] | ||
| Modulation of cell proliferation | tumour suppressor proteins (p53, p21) | human lung adenocarcinoma A549 cells | [31] |
| human hepatocellular carcinoma HepG2 cells | [54] | ||
| Cell cycle checkpoints | human colonic cancer CACO-2 cells | [61] | |
| human leukemia cells | [64] | ||
| [32] | |||
| pro- apoptotic signals | human colonic cancer CACO-2 cells | [61] | |
| human leukemia cells | [64] | ||
| [32] | |||
| human hepatocellular carcinoma HepG2 cells | [54] | ||
| androgen receptor (AR) | human prostate cancer LNCaP cell line | [63] | |
| reactive oxygen species (ROS) | human acute myeloid leukaemia HL60 and erythroblastic chronic myelogenous leukemia K562 cell lines | [105] |
5.1. Effects of ER on xenobiotic metabolism processes
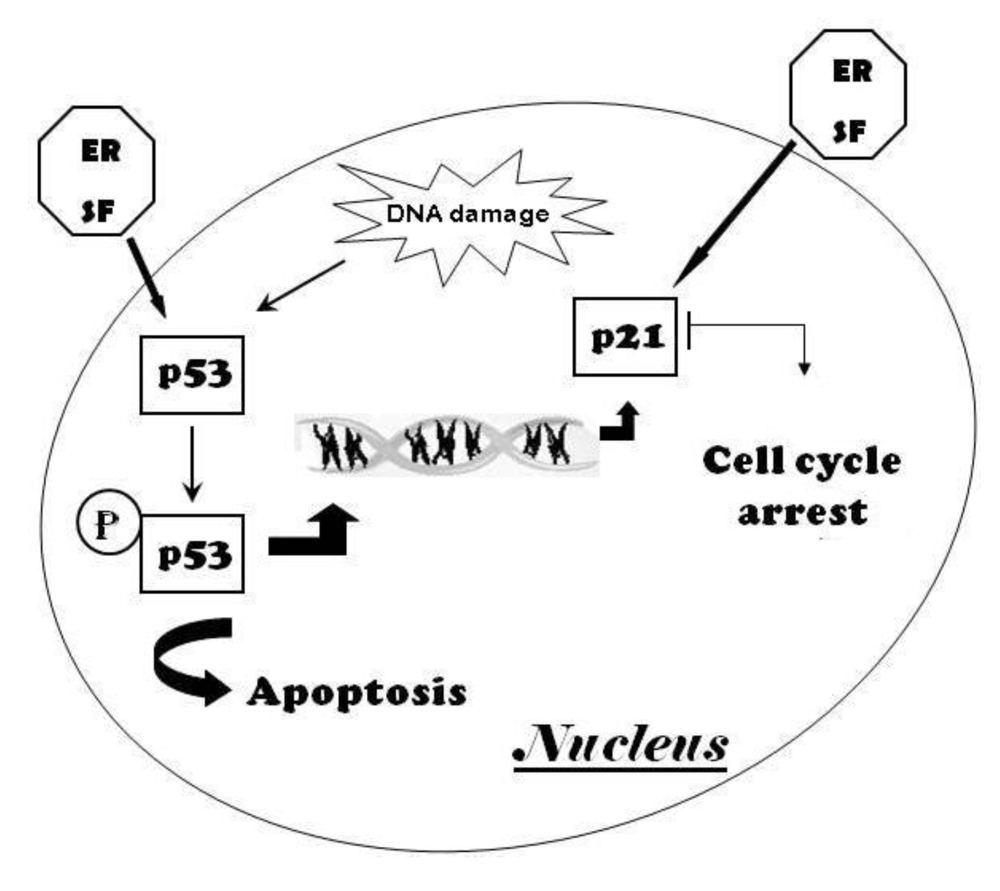
5.2. Effects of ER on the physiological control of cell proliferation

5.3. Effects of ER trough ROS-mediated mechanisms
5.4. Down-regulation of androgen receptor (AR) signaling pathway as novel mechanism of chemoprevention by ER
6. Conclusions and Remarks
References
- AICR. Food, Nutrition, Physical Activity, and the Prevention of Cancer: A Global Perspective; World Cancer Research Fund/American Institute for Cancer Research: Washington, DC, USA, 2007. [Google Scholar]
- Ambrosone, C.B.; Tang, L. Cruciferous vegetable intake and cancer prevention: Role of nutrigenetics. Cancer Prev. Res. (Phila Pa) 2009, 2, 298–300. [Google Scholar] [CrossRef]
- Traka, M.; Gasper, A.V.; Melchini, A.; Bacon, J.R.; Needs, P.W.; Frost, V.; Chantry, A.; Jones, A.M.; Ortori, C.A.; Barrett, D.A.; Ball, R.Y.; Mills, R.D.; Mithen, R.F. Broccoli consumption interacts with GSTM1 to perturb oncogenic signalling pathways in the prostate. PLoS One 2008, 3, e2568. [Google Scholar]
- Giovannucci, E.; Rimm, E.B.; Liu, Y.; Stampfer, M.J.; Willett, W.C. A prospective study of cruciferous vegetables and prostate cancer. Cancer Epidemiol. Biomarkers Prev. 2003, 12, 1403–1409. [Google Scholar]
- Kristal, A.R.; Lampe, J.W. Brassica vegetables and prostate cancer risk: a review of the epidemiological evidence. Nutr. Cancer 2002, 42, 1–9. [Google Scholar]
- Lampe, J.W.; Peterson, S. Brassica, biotransformation and cancer risk: Genetic polymorphisms alter the preventive effects of cruciferous vegetables. J. Nutr. 2002, 132, 2991–2994. [Google Scholar]
- Higdon, J.V.; Delage, B.; Williams, D.E.; Dashwood, R.H. Cruciferous vegetables and human cancer risk: Epidemiologic evidence and mechanistic basis. Pharmacol. Res. 2007, 55, 224–236. [Google Scholar]
- Lynn, A.; Collins, A.; Fuller, Z.; Hillman, K.; Ratcliffe, B. Cruciferous vegetables and colo-rectal cancer. Proc. Nutr. Soc. 2006, 65, 135–144. [Google Scholar]
- Munday, R.; Mhawech-Fauceglia, P.; Munday, C.M.; Paonessa, J.D.; Tang, L.; Munday, J.S.; Lister, C.; Wilson, P.; Fahey, J.W.; Davis, W.; Zhang, Y. Inhibition of urinary bladder carcinogenesis by broccoli sprouts. Cancer Res. 2008, 68, 1593–1600. [Google Scholar]
- Tang, L.; Zirpoli, G.R.; Guru, K.; Moysich, K.B.; Zhang, Y.; Ambrosone, C.B.; McCann, S.E. Consumption of raw cruciferous vegetables is inversely associated with bladder cancer risk. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 938–944. [Google Scholar]
- Thomson, C.A.; Rock, C.L.; Caan, B.J.; Flatt, S.W.; Al-Delaimy, W.A.; Newman, V.A.; Hajek, R.A.; Chilton, J.A.; Pierce, J.P. Increase in cruciferous vegetable intake in women previously treated for breast cancer participating in a dietary intervention trial. Nutr. Cancer 2007, 57, 11–19. [Google Scholar]
- Bonnesen, C.; Eggleston, I.M.; Hayes, J.D. Dietary indoles and isothiocyanates that are generated from cruciferous vegetables can both stimulate apoptosis and confer protection against DNA damage in human colon cell lines. Cancer Res. 2001, 61, 6120–6130. [Google Scholar]
- Hecht, S.S. Inhibition of carcinogenesis by isothiocyanates. Drug Metab. Rev. 2000, 32, 395–411. [Google Scholar]
- Zhang, Y.; Munday, R.; Jobson, H.E.; Munday, C.M.; Lister, C.; Wilson, P.; Fahey, J.W.; Mhawech-Fauceglia, P. Induction of GST and NQO1 in Cultured bladder cells and in the Urinary bladders of rats by an Extract of broccoli (Brassica oleracea italica) sprouts. J. Agric. Food Chem. 2006, 54, 9370–9376. [Google Scholar]
- Zhao, H.; Lin, J.; Grossman, H.B.; Hernandez, L.M.; Dinney, C.P.; Wu, X. Dietary isothiocyanates, GSTM1, GSTT1, NAT2 polymorphisms and bladder cancer risk. Int. J. Cancer 2007, 120, 2208–2213. [Google Scholar]
- Bianchini, F.; Vainio, H. Isothiocyanates in cancer prevention. Drug Metab. Rev. 2004, 36, 655–667. [Google Scholar]
- Hayes, J.D.; Kelleher, M.O.; Eggleston, I.M. The cancer chemopreventive actions of phytochemicals derived from glucosinolates. Eur. J. Nutr. 2008, 47 (Suppl. 2), 73–88. [Google Scholar] [PubMed]
- Keum, Y.S.; Jeong, W.S.; Kong, A.N. Chemopreventive functions of isothiocyanates. Drug News Perspect. 2005, 18, 445–451. [Google Scholar]
- Munday, R.; Zhang, Y.; Fahey, J.W.; Jobson, H.E.; Munday, C.M.; Li, J.; Stephenson, K.K. Evaluation of isothiocyanates as potent inducers of carcinogen-detoxifying enzymes in the urinary bladder: Critical nature of in vivo bioassay. Nutr. Cancer 2006, 54, 223–231. [Google Scholar]
- Nakamura, Y. Chemoprevention by isothiocyanates: Molecular basis of apoptosis induction. Forum Nutr. 2009, 61, 170–181. [Google Scholar]
- Wu, X.; Zhou, Q.H.; Xu, K. Are isothiocyanates potential anti-cancer drugs? Acta Pharmacol. Sin. 2009, 30, 501–512. [Google Scholar] [PubMed]
- Yang, G.; Gao, Y.T.; Shu, X.O.; Cai, Q.; Li, G.L.; Li, H.L.; Ji, B.T.; Rothman, N.; Dyba, M.; Xiang, Y.B.; Chung, F.L.; Chow, W.H.; Zheng, W. Isothiocyanate exposure, glutathione S-transferase polymorphisms, and colorectal cancer risk. Am. J. Clin. Nutr. 2009, 91. [Google Scholar]
- Bogani, P.; Visioli, F. Antioxidants in the Mediterranean diets: An update. World Rev. Nutr. Diet 2007, 97, 162–179. [Google Scholar]
- Schaffer, S.; Schmitt-Schillig, S.; Muller, W.E.; Eckert, G.P. Antioxidant properties of Mediterranean food plant extracts: geographical differences. J. Physiol. Pharmacol. 2005, 56 (Suppl. 1), 115–124. [Google Scholar] [PubMed]
- Bennett, R.N.; Rosa, E.A.; Mellon, F.A.; Kroon, P.A. Ontogenic profiling of glucosinolates, flavonoids, and other secondary metabolites in Eruca sativa (salad rocket), Diplotaxis erucoides (wall rocket), Diplotaxis tenuifolia (wild rocket), and Bunias orientalis (Turkish rocket). J. Agric. Food Chem. 2006, 54, 4005–4015. [Google Scholar] [PubMed]
- Vermeulen, M.; van den Berg, R.; Freidig, A.P.; van Bladeren, P.J.; Vaes, W.H. Association between consumption of cruciferous vegetables and condiments and excretion in urine of isothiocyanate mercapturic acids. J. Agric. Food Chem. 2006, 54, 5350–5358. [Google Scholar]
- Martinez-Sanchez, A.; Gil-Izquierdo, A.; Gil, M.I.; Ferreres, F. A comparative study of flavonoid compounds, vitamin C, and antioxidant properties of baby leaf Brassicaceae species. J. Agric. Food Chem. 2008, 56, 2330–2340. [Google Scholar]
- D'Antuono, L.F.; Elementi, S.; Neri, R. Glucosinolates in Diplotaxis and Eruca leaves: Diversity, taxonomic relations and applied aspects. Phytochemistry 2008, 69, 187–199. [Google Scholar]
- Cerny, M.S.; Taube, E.; Battaglia, R. Identification of bis(4-isothiocyanatobutyl) disulfide and its precursor from Rocket salad (Eruca sativa). J. Agric. Food Chem. 1996, 44, 3835–3839. [Google Scholar]
- Iori, R.; Bernardi, R.; Gueyrard, D.; Rollin, P.; Palmieri, S. Formation of glucoraphanin by chemoselective oxidation of natural glucoerucin: a chemoenzymatic route to sulforaphane. Bioorg. Med. Chem. Lett. 1999, 9, 1047–1048. [Google Scholar]
- Melchini, A.; Costa, C.; Traka, M.; Miceli, N.; Mithen, R.; De Pasquale, R.; Trovato, A. Erucin, a new promising cancer chemopreventive agent from rocket salads, shows anti-proliferative activity on human lung carcinoma A549 cells. Food Chem. Toxicol. 2009, 47, 1430–1436. [Google Scholar]
- Fimognari, C.; Nusse, M.; Iori, R.; Cantelli-Forti, G.; Hrelia, P. The new isothiocyanate 4-(methylthio)butylisothiocyanate selectively affects cell-cycle progression and apoptosis induction of human leukemia cells. Invest. New Drugs 2004, 22, 119–129. [Google Scholar]
- Gmelin, R.; Schluter, M. Isolation of 4-methylthiobutylglucosinolate (glucoerucin) from seeds of Eruca sativa Mill. Arch. Pharm. Ber. Dtsch. Pharm. Ges. 1970, 303, 330–334. [Google Scholar]
- Leoni, O.; Iori, R.; Palmieri, S.; Esposito, E.; Menegatti, E.; Cortesi, R.; Nastruzzi, C. Myrosinase-generated isothiocyanate from glucosinolates: Isolation, characterization and in vitro antiproliferative studies. Bioorg. Med. Chem. 1997, 5, 1799–1806. [Google Scholar]
- Kassahun, K.; Davis, M.; Hu, P.; Martin, B.; Baillie, T. Biotransformation of the naturally occurring isothiocyanate sulforaphane in the rat: Identification of phase I metabolites and glutathione conjugates. Chem. Res. Toxicol. 1997, 10, 1228–1233. [Google Scholar]
- Juge, N.; Mithen, R.F.; Traka, M. Molecular basis for chemoprevention by sulforaphane: A comprehensive review. Cell Mol. Life Sci. 2007, 64, 1105–1127. [Google Scholar]
- Kolm, R.H.; Danielson, U.H.; Zhang, Y.; Talalay, P.; Mannervik, B. Isothiocyanates as substrates for human glutathione transferases: structure-activity studies. Biochem. J. 1995, 311 (Part 2), 453–459. [Google Scholar]
- Jiao, D.; Ho, C.T.; Foiles, P.; Chung, F.L. Identification and quantification of the N-acetylcysteine conjugate of allyl isothiocyanate in human urine after ingestion of mustard. Cancer Epidemiol. Biomarkers Prev. 1994, 3, 487–492. [Google Scholar]
- Bheemreddy, R.M.; Jeffery, E.H. The metabolic fate of purified glucoraphanin in F344 rats. J. Agric. Food Chem. 2007, 55, 2861–2866. [Google Scholar]
- Conaway, C.C.; Krzeminski, J.; Amin, S.; Chung, F.L. Decomposition rates of isothiocyanate conjugates determine their activity as inhibitors of cytochrome p450 enzymes. Chem. Res. Toxicol. 2001, 14, 1170–1176. [Google Scholar]
- Al Janobi, A.A.; Mithen, R.F.; Gasper, A.V.; Shaw, P.N.; Middleton, R.J.; Ortori, C.A.; Barrett, D.A. Quantitative measurement of sulforaphane, iberin and their mercapturic acid pathway metabolites in human plasma and urine using liquid chromatography-tandem electrospray ionisation mass spectrometry. J. Chromatogr. B 2006, 844, 223–234. [Google Scholar]
- Gasper, A.V.; Al-Janobi, A.; Smith, J.A.; Bacon, J.R.; Fortun, P.; Atherton, C.; Taylor, M.A.; Hawkey, C.J.; Barrett, D.A.; Mithen, R.F. Glutathione S-transferase M1 polymorphism and metabolism of sulforaphane from standard and high-glucosinolate broccoli. Am. J. Clin. Nutr. 2005, 82, 1283–1291. [Google Scholar]
- Conaway, C.C.; Wang, C.X.; Pittman, B.; Yang, Y.M.; Schwartz, J.E.; Tian, D.; McIntee, E.J.; Hecht, S.S.; Chung, F.L. Phenethyl isothiocyanate and sulforaphane and their N-acetylcysteine conjugates inhibit malignant progression of lung adenomas induced by tobacco carcinogens in A/J mice. Cancer Res. 2005, 65, 8548–8557. [Google Scholar]
- Uphof, J.C.T. Dictionary of Economic Plants; Verlag Von J. Cramer Publ.: New York, NY, USA, 1968. [Google Scholar]
- Balme, F. Plantas Medicinais; Hemus Livraria Editora Limitada: São Paulo, Brazil, 1978. [Google Scholar]
- Yaniv, Z.; Schafferman, D.; Amar, Z. Tradition, uses and biodiversity of rocket (Eruca sativa, Brassicaceae) in Israel. Econ. Bot. 1998, 52, 394–400. [Google Scholar] [CrossRef]
- Perry, L.M.; Metzger, J. Medicinal Plants of SE Asia: Attributed Properties and Uses; The MIT Press: Cambridge, London, UK; Massachusetts, USA, 1978; pp. 23–24. [Google Scholar]
- El-Missiry, M.A.; El Gindy, A.M. Amelioration of alloxan induced diabetes mellitus and oxidative stress in rats by oil of Eruca sativa seeds. Ann. Nutr. Metab. 2000, 44, 97–100. [Google Scholar]
- Sarwar Alam, M.; Kaur, G.; Jabbar, Z.; Javed, K.; Athar, M. Eruca sativa seeds possess antioxidant activity and exert a protective effect on mercuric chloride induced renal toxicity. Food Chem. Toxicol. 2007, 45, 910–920. [Google Scholar]
- Barillari, J.; Canistro, D.; Paolini, M.; Ferroni, F.; Pedulli, G.F.; Iori, R.; Valgimigli, L. Direct antioxidant activity of purified glucoerucin, the dietary secondary metabolite contained in rocket (Eruca sativa Mill.) seeds and sprouts. J. Agric. Food Chem. 2005, 53, 2475–2482. [Google Scholar] [PubMed]
- Heimler, D.; Isolani, L.; Vignolini, P.; Tombelli, S.; Romani, A. Polyphenol content and antioxidative activity in some species of freshly consumed salads. J. Agric. Food Chem. 2007, 55, 1724–1729. [Google Scholar]
- Kim, S.J.; Jin, S.; Ishii, G. Isolation and structural elucidation of 4-(beta-D-glucopyranosyldisulfanyl)butyl glucosinolate from leaves of rocket salad (Eruca sativa L.) and its antioxidative activity. Biosci. Biotechnol. Biochem. 2004, 68, 2444–2450. [Google Scholar] [CrossRef] [PubMed]
- Alqasoumi, S.; Al-Sohaibani, M.; Al-Howiriny, T.; Al-Yahya, M.; Rafatullah, S. Rocket "Eruca sativa": A salad herb with potential gastric anti-ulcer activity. World J. Gastroenterol. 2009, 15, 1958–1965. [Google Scholar]
- Lamy, E.; Mersch-Sundermann, V. MTBITC mediates cell cycle arrest and apoptosis induction in human HepG2 cells despite its rapid degradation kinetics in the in vitro model. Environ. Mol. Mutagen. 2009, 50, 190–200. [Google Scholar]
- Jin, J.; Koroleva, O.A.; Gibson, T.; Swanston, J.; Magan, J.; Zhang, Y.; Rowland, I.R.; Wagstaff, C. Analysis of phytochemical composition and chemoprotective capacity of rocket (Eruca sativa and Diplotaxis tenuifolia) leafy salad following cultivation in different environments. J. Agric. Food Chem. 2009, 57, 5227–5234. [Google Scholar]
- Zhang, Y.; Talalay, P.; Cho, C.G.; Posner, G.H. A major inducer of anticarcinogenic protective enzymes from broccoli: Isolation and elucidation of structure. Proc. Natl. Acad. Sci. USA 1992, 89, 2399–2403. [Google Scholar]
- Hanlon, N.; Coldham, N.; Sauer, M.J.; Ioannides, C. Up-regulation of the CYP1 family in rat and human liver by the aliphatic isothiocyanates erucin and sulforaphane. Toxicology 2008, 252, 92–98. [Google Scholar] [Green Version]
- Hanlon, N.; Coldham, N.; Sauer, M.J.; Ioannides, C. Modulation of rat pulmonary carcinogen-metabolising enzyme systems by the isothiocyanates erucin and sulforaphane. Chem. Biol. Interact 2009, 177, 115–120. [Google Scholar] [Green Version]
- Hanlon, N.; Poynton, C.L.; Coldham, N.; Sauer, M.J.; Ioannides, C. The aliphatic isothiocyanates erucin and sulforaphane do not effectively up-regulate NAD(P)H:quinone oxidoreductase (NQO1) in human liver compared with rat. Mol. Nutr. Food Res. 2009, 53, 836–844. [Google Scholar]
- Wang, W.; Wang, S.; Howie, A.F.; Beckett, G.J.; Mithen, R.; Bao, Y. Sulforaphane, erucin, and iberin up-regulate thioredoxin reductase 1 expression in human MCF-7 cells. J. Agric. Food Chem. 2005, 53, 1417–1421. [Google Scholar]
- Munday, R.; Munday, C.M. Induction of phase II detoxification enzymes in rats by plant-derived isothiocyanates: Comparison of allyl isothiocyanate with sulforaphane and related compounds. J. Agric. Food Chem. 2004, 52, 1867–1871. [Google Scholar]
- Jakubikova, J.; Sedlak, J.; Mithen, R.; Bao, Y. Role of PI3K/Akt and MEK/ERK signaling pathways in sulforaphane-and erucin-induced phase II enzymes and MRP2 transcription, G2/M arrest and cell death in Caco-2 cells. Biochem. Pharmacol. 2005, 69, 1543–1552. [Google Scholar]
- Kim, S.H.; Singh, S.V. D,L-Sulforaphane causes transcriptional repression of androgen receptor in human prostate cancer cells. Mol. Cancer Ther. 2009, 8, 1946–1954. [Google Scholar]
- Jakubikova, J.; Bao, Y.; Sedlak, J. Isothiocyanates induce cell cycle arrest, apoptosis and mitochondrial potential depolarization in HL-60 and multidrug-resistant cell lines. Anticancer Res. 2005, 25, 3375–3386. [Google Scholar]
- Fimognari, C.; Nusse, M.; Berti, F.; Iori, R.; Cantelli-Forti, G.; Hrelia, P. Sulforaphane modulates cell cycle and apoptosis in transformed and non-transformed human T lymphocytes. Ann. N.Y. Acad. Sci. 2003, 1010, 393–398. [Google Scholar]
- Trachootham, D.; Zhou, Y.; Zhang, H.; Demizu, Y.; Chen, Z.; Pelicano, H.; Chiao, P.J.; Achanta, G.; Arlinghaus, R.B.; Liu, J.; Huang, P. Selective killing of oncogenically transformed cells through a ROS-mediated mechanism by beta-phenylethyl isothiocyanate. Cancer Cell. 2006, 10, 241–252. [Google Scholar]
- Singh, M.S.; Michael, M. Role of xenobiotic metabolic enzymes in cancer epidemiology. Methods Mol. Biol. 2009, 472, 243–264. [Google Scholar]
- Yu, S.; Kong, A.N. Targeting carcinogen metabolism by dietary cancer preventive compounds. Curr. Cancer Drug Targets 2007, 7, 416–424. [Google Scholar]
- Zhou, C.; Poulton, E.J.; Grun, F.; Bammler, T.K.; Blumberg, B.; Thummel, K.E.; Eaton, D.L. The dietary isothiocyanate sulforaphane is an antagonist of the human steroid and xenobiotic nuclear receptor. Mol. Pharmacol. 2007, 71, 220–229. [Google Scholar]
- Yoxall, V.; Kentish, P.; Coldham, N.; Kuhnert, N.; Sauer, M.J.; Ioannides, C. Modulation of hepatic cytochromes P450 and phase II enzymes by dietary doses of sulforaphane in rats: Implications for its chemopreventive activity. Int. J. Cancer 2005, 117, 356–362. [Google Scholar]
- Gross-Steinmeyer, K.; Stapleton, P.L.; Tracy, J.H.; Bammler, T.K.; Lehman, T.; Strom, S.C.; Eaton, D.L. Influence of Matrigel-overlay on constitutive and inducible expression of nine genes encoding drug-metabolizing enzymes in primary human hepatocytes. Xenobiotica 2005, 35, 419–438. [Google Scholar]
- Barcelo, S.; Mace, K.; Pfeifer, A.M.; Chipman, J.K. Production of DNA strand breaks by N-nitrosodimethylamine and 2-amino-3-methylimidazo[4,5-f]quinoline in THLE cells expressing human CYP isoenzymes and inhibition by sulforaphane. Mutat. Res. 1998, 402, 111–120. [Google Scholar]
- Maheo, K.; Morel, F.; Langouet, S.; Kramer, H.; Le Ferrec, E.; Ketterer, B.; Guillouzo, A. Inhibition of cytochromes P-450 and induction of glutathione S-transferases by sulforaphane in primary human and rat hepatocytes. Cancer Res. 1997, 57, 3649–3652. [Google Scholar]
- Barcelo, S.; Gardiner, J.M.; Gescher, A.; Chipman, J.K. CYP2E1-mediated mechanism of anti-genotoxicity of the broccoli constituent sulforaphane. Carcinogenesis 1996, 17, 277–282. [Google Scholar]
- Yang, C.S.; Smith, T.J.; Hong, J.Y. Cytochrome P-450 enzymes as targets for chemoprevention against chemical carcinogenesis and toxicity: opportunities and limitations. Cancer Res. 1994, 54, 1982s–1986s. [Google Scholar]
- Zhang, Y.; Talalay, P. Anticarcinogenic activities of organic isothiocyanates: Chemistry and mechanisms. Cancer Res. 1994, 54, 1976s–1981s. [Google Scholar]
- Lamy, E.; Schroder, J.; Paulus, S.; Brenk, P.; Stahl, T.; Mersch-Sundermann, V. Antigenotoxic properties of Eruca sativa (rocket plant), erucin and erysolin in human hepatoma (HepG2) cells towards benzo(a)pyrene and their mode of action. Food Chem. Toxicol. 2008, 46, 2415–2421. [Google Scholar]
- Bollard, M.; Stribbling, S.; Mitchell, S.; Caldwell, J. The disposition of allyl isothiocyanate in the rat and mouse. Food Chem. Toxicol. 1997, 35, 933–943. [Google Scholar]
- van Bladeren, P.J.; den Besten, C.; Bruggeman, I.M.; Mertens, J.J.W.M.; van Ommen, B.; Spenkelink, B.; Rutten, A.L.M.; Temmink, J.H.M.; Vos, R.M.E. Glutathione conjugation as a toxication reaction. In Metabolism of Xenobiotics; Gorrod, J.W, Oelschlager, H., Caldwell, J., Eds.; Taylor and Francis: London, UK, 1988; pp. 267–274. [Google Scholar]
- Rushmore, T.H.; Morton, M.R.; Pickett, C.B. The antioxidant responsive element. Activation by oxidative stress and identification of the DNA consensus sequence required for functional activity. J. Biol. Chem. 1991, 266, 11632–11639. [Google Scholar] [PubMed]
- Cole, S.P.; Bhardwaj, G.; Gerlach, J.H.; Mackie, J.E.; Grant, C.E.; Almquist, K.C.; Stewart, A.J.; Kurz, E.U.; Duncan, A.M.; Deeley, R.G. Overexpression of a transporter gene in a multidrug-resistant human lung cancer cell line. Science 1992, 258, 1650–1654. [Google Scholar]
- Bates, S.E.; Robey, R.; Miyake, K.; Rao, K.; Ross, D.D.; Litman, T. The role of half-transporters in multidrug resistance. J. Bioenerg. Biomembr. 2001, 33, 503–511. [Google Scholar]
- Awasthi, Y.C.; Sharma, R.; Yadav, S.; Dwivedi, S.; Sharma, A.; Awasthi, S. The non-ABC drug transporter RLIP76 (RALBP-1) plays a major role in the mechanisms of drug resistance. Curr. Drug Metab. 2007, 8, 315–323. [Google Scholar]
- Borst, P.; Elferink, R.O. Mammalian ABC transporters in health and disease. Ann. Rev. Biochem. 2002, 71, 537–592. [Google Scholar]
- Bandler, P.E.; Westlake, C.J.; Grant, C.E.; Cole, S.P.; Deeley, R.G. Identification of regions required for apical membrane localization of human multidrug resistance protein 2. Mol. Pharmacol. 2008, 74, 9–19. [Google Scholar]
- Adachi, T.; Nakagawa, H.; Chung, I.; Hagiya, Y.; Hoshijima, K.; Noguchi, N.; Kuo, M.T.; Ishikawa, T. Nrf2-dependent and -independent induction of ABC transporters ABCC1, ABCC2, and ABCG2 in HepG2 cells under oxidative stress. J. Exp. Ther. Oncol. 2007, 6, 335–348. [Google Scholar]
- Barancik, M.; Bohacova, V.; Sedlak, J.; Sulova, Z.; Breier, A. LY294,002, a specific inhibitor of PI3K/Akt kinase pathway, antagonizes P-glycoprotein-mediated multidrug resistance. Eur. J. Pharm. Sci. 2006, 29, 426–434. [Google Scholar]
- Brugge, J.; Hung, M.C.; Mills, G.B. A new mutational AKTivation in the PI3K pathway. Cancer Cell 2007, 12, 104–107. [Google Scholar]
- Payen, L.; Courtois, A.; Loewert, M.; Guillouzo, A.; Fardel, O. Reactive oxygen species-related induction of multidrug resistance-associated protein 2 expression in primary hepatocytes exposed to sulforaphane. Biochem. Biophys. Res. Commun. 2001, 282, 257–263. [Google Scholar]
- Traka, M.; Gasper, A.V.; Smith, J.A.; Hawkey, C.J.; Bao, Y.; Mithen, R.F. Transcriptome analysis of human colon Caco-2 cells exposed to sulforaphane. J. Nutr. 2005, 135, 1865–1872. [Google Scholar]
- Harris, K.E.; Jeffery, E.H. Sulforaphane and erucin increase MRP1 and MRP2 in human carcinoma cell lines. J. Nutr. Biochem. 2008, 19, 246–254. [Google Scholar]
- Mi, L.; Xiao, Z.; Hood, B.L.; Dakshanamurthy, S.; Wang, X.; Govind, S.; Conrads, T.P.; Veenstra, T.D.; Chung, F.L. Covalent binding to tubulin by isothiocyanates. A mechanism of cell growth arrest and apoptosis. J. Biol. Chem. 2008, 283, 22136–22146. [Google Scholar] [PubMed]
- Chuang, L.T.; Moqattash, S.T.; Gretz, H.F.; Nezhat, F.; Rahaman, J.; Chiao, J.W. Sulforaphane induces growth arrest and apoptosis in human ovarian cancer cells. Acta Obstet. Gynecol. Scand. 2007, 1–6. [Google Scholar]
- Cuddihy, S.L.; Brown, K.K.; Thomson, S.J.; Hampton, M.B. Induction of apoptosis by phenethyl isothiocyanate in cells overexpressing Bcl-XL. Cancer Lett. 2008, 271, 215–221. [Google Scholar]
- Jadhav, U.; Ezhilarasan, R.; Vaughn, S.F.; Berhow, M.A.; Mohanam, S. Iberin induces cell cycle arrest and apoptosis in human neuroblastoma cells. Int. J. Mol. Med. 2007, 19, 353–361. [Google Scholar]
- Kuang, Y.F.; Chen, Y.H. Induction of apoptosis in a non-small cell human lung cancer cell line by isothiocyanates is associated with P53 and P21. Food Chem. Toxicol. 2004, 42, 1711–1718. [Google Scholar]
- Mas, S.; Crescenti, A.; Gasso, P.; Deulofeu, R.; Molina, R.; Ballesta, A.; Kensler, T.W.; Lafuente, A. Induction of apoptosis in HT-29 cells by extracts from isothiocyanates-rich varieties of Brassica oleracea. Nutr. Cancer 2007, 58, 107–114. [Google Scholar]
- Prives, C.; Hall, P.A. The p53 pathway. J. Pathol. 1999, 187, 112–126. [Google Scholar]
- Tibbetts, R.S.; Brumbaugh, K.M.; Williams, J.M.; Sarkaria, J.N.; Cliby, W.A.; Shieh, S.Y.; Taya, Y.; Prives, C.; Abraham, R.T. A role for ATR in the DNA damage-induced phosphorylation of p53. Genes Dev. 1999, 13, 152–157. [Google Scholar]
- Mi, L.; Wang, X.; Govind, S.; Hood, B.L.; Veenstra, T.D.; Conrads, T.P.; Saha, D.T.; Goldman, R.; Chung, F.L. The role of protein binding in induction of apoptosis by phenethyl isothiocyanate and sulforaphane in human non-small lung cancer cells. Cancer Res. 2007, 67, 6409–6416. [Google Scholar]
- Barillari, J.; Iori, R.; Papi, A.; Orlandi, M.; Bartolini, G.; Gabbanini, S.; Pedulli, G.F.; Valgimigli, L. Kaiware Daikon (Raphanus sativus L.) extract: A naturally multipotent chemopreventive agent. J. Agric. Food Chem. 2008, 56, 7823–7830. [Google Scholar] [PubMed]
- Papi, A.; Orlandi, M.; Bartolini, G.; Barillari, J.; Iori, R.; Paolini, M.; Ferroni, F.; Grazia Fumo, M.; Pedulli, G.F.; Valgimigli, L. Cytotoxic and antioxidant activity of 4-methylthio-3-butenyl isothiocyanate from Raphanus sativus L. (Kaiware Daikon) sprouts. J. Agric. Food Chem. 2008, 56, 875–883. [Google Scholar] [PubMed]
- Rahman, K. Studies on free radicals, antioxidants, and co-factors. Clin. Interv. Aging. 2007, 2, 219–236. [Google Scholar]
- Wu, X.J.; Hua, X. Targeting ROS: Selective killing of cancer cells by a cruciferous vegetable derived pro-oxidant compound. Cancer Biol. Ther. 2007, 6, 646–647. [Google Scholar]
- Doudican, N.A.; Bowling, B.; Orlow, S.J. Enhancement of arsenic trioxide cytotoxicity by dietary isothiocyanates in human leukemic cells via a reactive oxygen species-dependent mechanism. Leuk Res. 2009, 34, 229–234. [Google Scholar]
- Nair, B.; Wilt, T.; MacDonald, R.; Rutks, I. Early versus deferred androgen suppression in the treatment of advanced prostatic cancer. Cochrane Database Syst. Rev. 2002. [Google Scholar]
- Scher, H.I.; Sawyers, C.L. Biology of progressive, castration-resistant prostate cancer: Directed therapies targeting the androgen-receptor signaling axis. J. Clin. Oncol. 2005, 23, 8253–8261. [Google Scholar]
- Taplin, M.E. Drug insight: Role of the androgen receptor in the development and progression of prostate cancer. Nat. Clin. Pract. Oncol. 2007, 4, 236–244. [Google Scholar]
- Cohen, J.H.; Kristal, A.R.; Stanford, J.L. Fruit and vegetable intakes and prostate cancer risk. J. Natl. Cancer Inst. 2000, 92, 61–68. [Google Scholar]
- Kolonel, L.N.; Hankin, J.H.; Whittemore, A.S.; Wu, A.H.; Gallagher, R.P.; Wilkens, L.R.; John, E.M.; Howe, G.R.; Dreon, D.M.; West, D.W.; Paffenbarger, R.S., Jr. Vegetables, fruits, legumes and prostate cancer: A multiethnic case-control study. Cancer Epidemiol. Biomarkers Prev. 2000, 9, 795–804. [Google Scholar] [PubMed]
- Wang, L.G.; Liu, X.M.; Chiao, J.W. Repression of androgen receptor in prostate cancer cells by phenethyl isothiocyanate. Carcinogenesis 2006, 27, 2124–2132. [Google Scholar]
- Gong, A.; He, M.; Krishna Vanaja, D.; Yin, P.; Karnes, R.J.; Young, C.Y. Phenethyl isothiocyanate inhibits STAT3 activation in prostate cancer cells. Mol. Nutr. Food Res. 2009, 53, 878–886. [Google Scholar]
- Gibbs, A.; Schwartzman, J.; Deng, V.; Alumkal, J. Sulforaphane destabilizes the androgen receptor in prostate cancer cells by inactivating histone deacetylase 6. Proc. Natl. Acad. Sci. USA 2009, 106, 16663–16668. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Melchini, A.; Traka, M.H. Biological Profile of Erucin: A New Promising Anticancer Agent from Cruciferous Vegetables. Toxins 2010, 2, 593-612. https://doi.org/10.3390/toxins2040593
Melchini A, Traka MH. Biological Profile of Erucin: A New Promising Anticancer Agent from Cruciferous Vegetables. Toxins. 2010; 2(4):593-612. https://doi.org/10.3390/toxins2040593
Chicago/Turabian StyleMelchini, Antonietta, and Maria H. Traka. 2010. "Biological Profile of Erucin: A New Promising Anticancer Agent from Cruciferous Vegetables" Toxins 2, no. 4: 593-612. https://doi.org/10.3390/toxins2040593
APA StyleMelchini, A., & Traka, M. H. (2010). Biological Profile of Erucin: A New Promising Anticancer Agent from Cruciferous Vegetables. Toxins, 2(4), 593-612. https://doi.org/10.3390/toxins2040593