Plant Natural Compounds with Antibacterial Activity towards Common Pathogens of Pond-Cultured Channel Catfish (Ictalurus punctatus)

Abstract
:1. Introduction
2. Results and Discussion


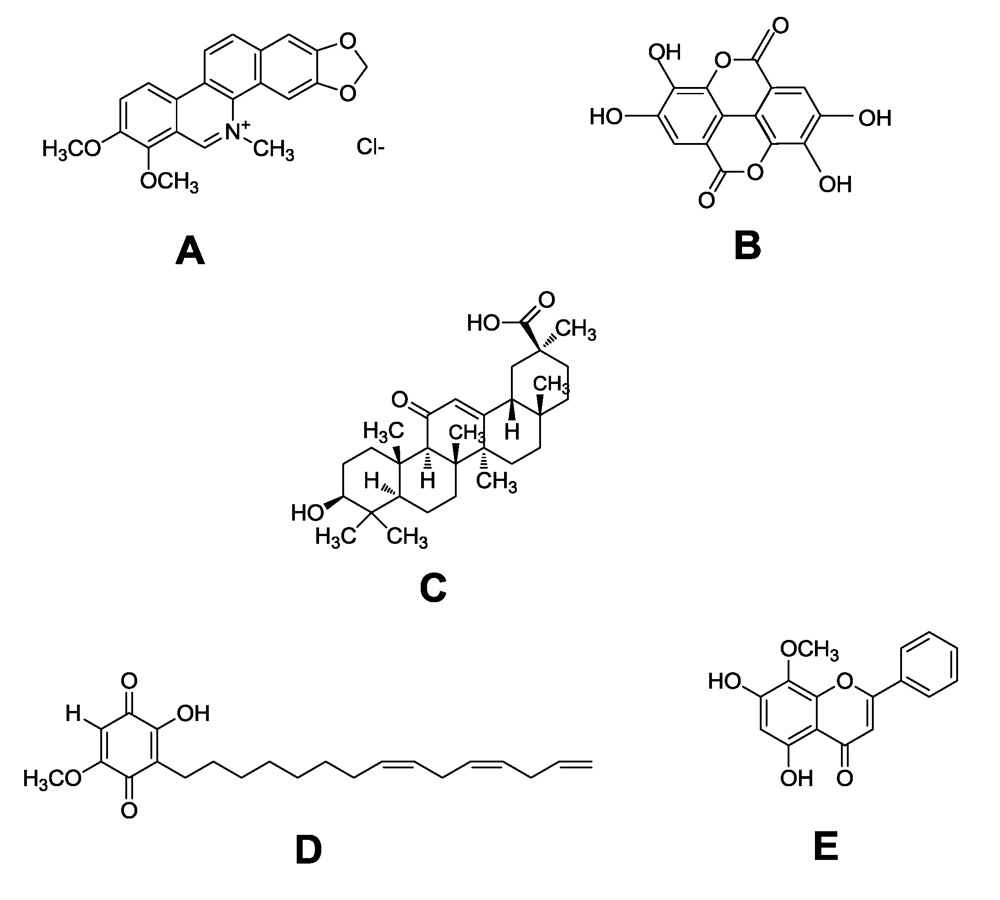{kind=link}
| Compound | 24-h IC50a | MICb | 24-h IC50 | MIC | ||
|---|---|---|---|---|---|---|
| (mg/L) | (mg/L) | RDCFc | RDCOd | RDCFc | RDCOd | |
| Acacetin | >284.3 | >284.3 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Allyl disulfide | >146.3 | 146.3 | >2040.8 | >3846.2 | 1000.0 | 1111.1 |
| Apigenin | >270.2 | >270.2 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Apigenin 7-glucoside | >432.4 | >432.4 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Baicalein | 243.2 | 27.0 | 1839.7 | 3461.5 | 100.0 | 111.1 |
| Baicalin | >446.4 | >446.4 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Camphor | >152.2 | >154.2 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Chelerythrine chloride | 7.3 | 2.1 | 38.8 | 73.1 | 10.0 | 11.1 |
| Cichoriin | 54.4 | 178.1 | 622.3 | 1173.1 | 1000.0 | 1111.1 |
| p-Coumaric acid | >164.2 | 164.2 | >2040.8 | >3846.2 | 1000.0 | 1111.1 |
| Cyanidin chloride | >287.2 | >287.2 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| p-Cymene | >134.2 | >134.2 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Daidzein | >254.2 | >254.2 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Delphinidin chloride | 101.6 | >338.7 | 612.2 | 1153.9 | >1000.0 | >1111.1 |
| Ellagic acid | 15.1 | 6.5 | 83.3 | 143.6 | 10.0 | 10.6 |
| Embelin | >294.4 | >294.4 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Emetine | >553.6 | >553.6 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Eucalyptol | >154.3 | >154.3 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Flavone | >222.2 | >222.4 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Gallocatechin | 260.3 | 306.3 | 1734.7 | 3269.2 | 1000.0 | 1111.1 |
| Genistein | >270.2 | >270.2 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Glycyrrhetinic acid (α) | >470.7 | >470.7 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Glycyrrhetinic acid (β) | >470.7 | >470.7 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Hyperforin | >536.8 | >536.8 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Inulin | >522.5 | >522.5 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Lavandulol | 50.9 | 154.3 | 673.5 | 1269.2 | 1000.0 | 1111.1 |
| (S)-Limonene | >136.2 | >136.2 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Myrcene | >136.2 | >136.2 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Myricetin | >318.2 | 318.2 | >2040.8 | >3846.2 | 1000.0 | 1111.1 |
| Naringenin | >272.3 | 27.2 | >2040.8 | >3846.2 | 100.0 | 111.1 |
| Pelargonidin chloride | 85.9 | >306.7 | 571.4 | 1076.9 | >1000.0 | >1111.1 |
| Piceatannol | >244.3 | >244.3 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| ß-Sitosterol | >414.7 | >414.7 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Sorgoleone | >358.0 | >358.0 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Stigmasterol | >412.7 | >412.7 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Vitexin | >432.4 | >432.4 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Wogonin | >284.2 | >284.2 | >2040.8 | >3846.2 | >1000.0 | >1111.1 |
| Zingiberone | >194.2 | 194.2 | >2040.8 | >3846.2 | 1000.0 | 1111.1 |
| Compound | 24-h IC50a | MICb | 24-h IC50 | MIC | ||
|---|---|---|---|---|---|---|
| (mg/L) | (mg/L) | RDCFc | RDCOd | RDCFc | RDCOd | |
| Acacetin | >284.3 | >284.3 | >518.1 | >492.6 | >1000.0 | >1075.3 |
| Allyl disulfide | >146.3 | 146.3 | >518.1 | >492.6 | 1000.0 | 1075.3 |
| Apigenin | 22.8 | 27.0 | 43.8 | 41.6 | 100.0 | 107.5 |
| Apigenin 7-glucoside | 216.2 | >432.4 | 259.1 | 246.3 | >1000.0 | >1075.3 |
| Baicalein | 8.2 | 14.9 | 15.6 | 14.8 | 55.0 | 59.2 |
| Baicalin | 169.6 | 446.4 | 196.9 | 187.2 | 1000.0 | 1075.3 |
| Camphor | >152.2 | >154.2 | >518.1 | >492.6 | 1000.0 | 1075.3 |
| Chelerythrine chloride | 5.0 | 2.1 | 6.8 | 6.4 | 5.5 | 6.0 |
| Cichoriin | 12.4 | 17.8 | 36.0 | 34.3 | 100.0 | 107.5 |
| p-Coumaric acid | >164.2 | 164.2 | >518.1 | >492.6 | 1000.0 | 1075.3 |
| Cyanidin chloride | >322.7 | >322.7 | >518.1 | >492.6 | >1000.0 | >1075.3 |
| p-Cymene | >134.2 | >134.2 | >518.1 | >492.6 | >1000.0 | >1075.3 |
| Daidzein | >254.2 | >254.2 | >518.1 | >492.6 | >1000.0 | >1075.3 |
| Delphinidin chloride | 54.2 | >338.7 | 82.9 | 78.8 | >1000.0 | >1075.3 |
| Ellagic acid | 9.7 | 3.0 | 19.1 | 15.2 | 10.0 | 10.4 |
| Embelin | 13.0 | 2.9 | 22.8 | 21.7 | 100.0 | 10.8 |
| Emetine | 83.0 | 55.4 | 77.7 | 73.9 | 100.0 | 10.8 |
| Eucalyptol | >154.3 | >154.3 | >518.1 | >492.6 | >1000.0 | >1075.3 |
| Flavone | 6.7 | 22.2 | 15.5 | 14.8 | 100.0 | 107.5 |
| Gallocatechin | 55.1 | 30.6 | 93.3 | 88.7 | 100.0 | 10.8 |
| Genistein | 16.8 | 27.0 | 32.1 | 30.6 | 100.0 | 107.5 |
| Glycyrrhetinic acid (α) | >470.7 | >470.7 | >518.1 | >492.6 | >1000.0 | >1075.3 |
| Glycyrrhetinic acid (β) | 10.2 | 2.6 | 11.2 | 10.6 | 5.5 | 6.0 |
| Hyperforin | >5.4 | >5.4 | >518.2 | >4.9 | >10.0 | >10.8 |
| Inulin | >522.5 | 522.5 | >518.1 | >492.6 | 1000.0 | 1075.3 |
| Lavandulol | 29.3 | 15.4 | 98.5 | 93.6 | 100.0 | 107.5 |
| (S)-Limonene | >136.2 | >136.2 | >518.1 | >492.6 | >1000.0 | >1075.3 |
| Myrcene | >136.2 | >136.2 | >518.1 | >492.6 | >1000.0 | >1075.3 |
| Myricetin | 28.6 | 31.8 | 46.6 | 44.3 | 100.0 | 107.5 |
| Naringenin | 46.3 | 27.2 | 88.1 | 83.7 | 100.0 | 107.5 |
| Pelargonidin chloride | 52.1 | 30.7 | 88.1 | 83.7 | 100.0 | 107.5 |
| Piceatannol | 9.7 | 24.4 | 20.5 | 19.5 | 100.0 | 107.5 |
| ß-Sitosterol | >414.7 | >414.7 | >518.1 | >492.6 | >1000.0 | >1075.3 |
| Sorgoleone | 9.0 | 3.6 | 13.0 | 12.3 | 10.0 | 10.8 |
| Stigmasterol | >412.7 | 412.7 | >518.1 | >492.6 | 1000.0 | 1075.3 |
| Vitexin | >432.4 | >432.4 | >518.1 | >492.6 | >1000.0 | >1075.3 |
| Wogonin | 28.4 | 0.3 | 51.8 | 49.3 | 1.0 | 1.1 |
| Zingiberone | >194.2 | >194.2 | >518.1 | >492.6 | >1000.0 | >1075.3 |
| Compound | 24-h IC50a | MICb | 24-h IC50 | MIC | ||
|---|---|---|---|---|---|---|
| (mg/L) | (mg/L) | RDCFc | RDCOd | RDCFc | RDCOd | |
| Acacetin | >284.3 | >284.3 | >423.7 | >588.2 | >1000.0 | >1075.3 |
| Allyl disulfide | >146.3 | 146.3 | >423.7 | >588.2 | 1000.0 | 1075.3 |
| Apigenin | 91.9 | 2.7 | 144.1 | 200.0 | 10.0 | 10.8 |
| Apigenin 7-glucoside | 255.1 | >432.4 | 250.0 | 347.0 | >1000.0 | >1075.3 |
| Baicalein | 8.2 | 15.0 | 12.7 | 17.7 | 55.0 | 59.2 |
| Baicalin | 116.1 | 446.4 | 110.2 | 152.9 | 1000.0 | 107.5 |
| Camphor | >152.2 | >152.2 | >423.7 | >588.2 | >1000.0 | >1075.3 |
| Chelerythrine chloride | 7.9 | 2.1 | 8.7 | 12.1 | 5.5 | 6.0 |
| Cichoriin | 7.5 | 17.8 | 17.8 | 24.8 | 100.0 | 107.5 |
| p-Coumaric acid | >164.2 | 164.2 | >423.7 | >588.2 | 1000.0 | 1075.3 |
| Cyanidin chloride | 242.0 | >322.7 | 317.8 | 441.2 | >1000.0 | >1075.3 |
| p-Cymene | >134.2 | >134.2 | >423.7 | >588.2 | >1000.0 | >1075.3 |
| Daidzein | >254.2 | >254.2 | >423.7 | >588.2 | >1000.0 | >1075.3 |
| Delphinidin chloride | 135.5 | 338.7 | 169.5 | 235.3 | 1000.0 | 1075.3 |
| Ellagic acid | 10.3 | 16.6 | 14.1 | 17.8 | 55.0 | 58.8 |
| Embelin | 53.0 | 29.4 | 76.3 | 105.9 | 100.0 | 107.5 |
| Emetine | 121.8 | 55.4 | 93.2 | 129.4 | 100.0 | 107.5 |
| Eucalyptol | >154.3 | >154.3 | >423.7 | >588.2 | >1000.0 | >1075.3 |
| Flavone | 7.8 | 22.2 | 14.8 | 20.6 | 100.0 | 107.5 |
| Gallocatechin | 147.0 | 306.3 | 203.4 | 282.4 | 1000.0 | 1075.3 |
| Genistein | 16.2 | 1.5 | 25.5 | 35.3 | 5.5 | 6.0 |
| Glycyrrhetinic acid (α) | >470.7 | >470.7 | >423.7 | >588.2 | >1000.0 | >1075.3 |
| Glycyrrhetinic acid (β) | 13.7 | 47.1 | 12.2 | 17.1 | 100.0 | 107.5 |
| Hyperforin | >5.4 | >5.4 | >4.2 | >5.9 | >10.0 | >10.8 |
| Inulin | >522.5 | >522.5 | >423.7 | >588.2 | >1000.0 | >1075.3 |
| Lavandulol | 29.3 | 15.4 | 80.5 | 111.8 | 100.0 | 107.5 |
| (S)-Limonene | >136.2 | >136.2 | >423.7 | >588.2 | >1000.0 | >1075.3 |
| Myrcene | >136.2 | >136.2 | >423.7 | >588.2 | >1000.0 | >1075.3 |
| Myricetin | >318.2 | 31.8 | >423.7 | >588.2 | 100.0 | 107.5 |
| Naringenin | 38.1 | 27.2 | 59.3 | 82.4 | 100.0 | 107.5 |
| Pelargonidin chloride | 116.5 | >306.7 | 161.0 | 223.5 | >1000.0 | >1075.3 |
| Piceatannol | 8.7 | 24.4 | 15.1 | 20.9 | 100.0 | 107.5 |
| ß-Sitosterol | >414.7 | >414.7 | >423.7 | >588.2 | >1000.0 | >1075.3 |
| Sorgoleone | 9.3 | 3.6 | 11.1 | 15.3 | 10.0 | 10.8 |
| Stigmasterol | >412.7 | 412.7 | >423.7 | >588.2 | 1000.0 | 1075.3 |
| Vitexin | >432.4 | >432.4 | >423.7 | >588.2 | >1000.0 | >1075.3 |
| Wogonin | 19.4 | 0.3 | 28.8 | 40.0 | 1.0 | 1.1 |
| Zingiberone | >194.2 | >194.2 | >423.7 | >588.2 | >1000.0 | >1075.3 |
3. Materials and Methods
| Compound | Sourcea | Solvent | Purity (%) |
|---|---|---|---|
| Acacetin | 1 | Ethanol | 97 |
| Allyl disulfide | 1 | Methanol | 80 |
| Apigenin | 1 | Ethanol | 95 |
| Apigenin 7-glucoside | 1 | Water | 97 |
| Baicalein | 1 | Ethanol | 98 |
| Baicalin | 1 | Acetone | 95 |
| Camphor | 1 | Methanol | 98 |
| Chelerythrine chloride | 1 | Water | 95 |
| Cichoriin | 1 | Methanol | 98 |
| p-Coumaric acid | 1 | Ethanol | 100 |
| Cyanidin chloride | 2 | Methanol | 100 |
| p-Cymene | 1 | Methanol | 99 |
| Daidzein | 1 | Methanol | 98 |
| Delphinidin chloride | 1 | Methanol | 95 |
| Embelin | 1 | Methanol | 98 |
| Emetine | 1 | Methanol | 97 |
| Eucalyptol | 3 | Methanol | 99 |
| Flavone | 1 | Acetone | 100 |
| (-)-Gallocatechin | 1 | Methanol | 98 |
| Genistein | 2 | Ethanol | 99 |
| α-Glycyrrhetinic acid | 1 | Methanol | 98 |
| β-Glycyrrhetinic acid | 1 | Methanol | 97 |
| Hyperforin | 1 | Methanol | 85 |
| Inulin | 1 | Water | 100 |
| Lavandulol | 3 | Methanol | 90 |
| (S)-(-)-Limonene | 1 | Methanol | 96 |
| Myrcene | 3 | Dichloromethane | 90 |
| Myricetin | 2 | Ethanol | 100 |
| Naringenin | 1 | Ethanol | 100 |
| Pelargonidin chloride | 1 | Ethanol | 100 |
| Piceatannol | 4 | Ethanol | 98 |
| ß-Sitosterol | 1 | Methanol | 99 |
| Sorgoleone | 5 | Ethanol | 95 |
| Stigmasterol | 1 | Methanol | 99 |
| Vitexin | 1 | Methanol | 96 |
| Wogonin | 1 | Ethanol | 98 |
| Zingiberone | 1 | Ethanol | 96 |
4. Conclusions
Acknowledgements
References
- Hawke, J.P.; Durborow, R.M.; Thune, R.L.; Camus, A.C. ESC: Enteric Septicemia of Catfish; Southern Regional Aquaculture Center: Stoneville, MS, USA, 1998. [Google Scholar]
- Durborow, R.M.; Thune, R.L.; Hawke, J.P.; Camus, A.C. Columnaris Disease: A Bacterial Infection Caused by Flavobacterium columnare; Southern Regional Aquaculture Center: Stoneville, MS, USA, 1998. [Google Scholar]
- Plumb, J.A. Health Maintenance and Principal Microbial Diseases of Cultured Fishes; Iowa State University Press: Ames, IA, USA, 1999. [Google Scholar]
- Darwish, A.M.; Mitchell, A.J.; Straus, D.L. Evaluation of potassium permanganate against an experimental subacute infection of Flavobacterium columnare in channel catfish, Ictalurus punctatus (Rafinesque). J. Fish Dis. 2009, 32, 193–199. [Google Scholar]
- Boyd, C.E.; Tucker, C.S. Pond Aquaculture Water Quality Management; Kluwer Academic Publishers: Boston, MA, USA, 1998. [Google Scholar]
- Thomas-Jinu, S.; Goodwin, A.E. Acute columnaris infection in channel catfish, Ictalurus punctatus (Rafinesque): Efficacy of practical treatments for warmwater aquaculture ponds. J. Fish Dis. 2004, 27, 23–28. [Google Scholar]
- Darwish, A.M.; Mitchell, A.J. Evaluation of diquat against an acute experimental infection of Flavobacterium columnare in channel catfish, Ictalurus punctatus (Rafinesque). J. Fish Dis. 2009, 32, 401–408. [Google Scholar]
- Hawke, J.P.; Khoo, L.H. Infectious diseases. In Biology and Culture of Channel Catfish; Tucker, C.S., Hargreaves, J.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 387–443. [Google Scholar]
- Nascimento, G.G.F.; Locatelli, J.; Freitas, P.C.; Silva, G.L. Antibacterial activity of plant extracts and phytochemicals on antibiotic-resistant bacteria. Braz. J. Microbiol. 2000, 31, 247–256. [Google Scholar]
- Pérez, C.; Anesini, C. Antibacterial activity of alimentary plants against Staphylococcus aureus growth. Am. J. Chin. Med. 1994, 22, 169–174. [Google Scholar]
- Mansouri, S. Inhibition of Staphylococcus aureus mediated by extracts of Iranian plants. Pharm. Biol. 1999, 37, 375–377. [Google Scholar]
- Schrader, K.K.; Harries, M.D. A rapid bioassay for bactericides against the catfish pathogens Edwardsiella ictaluri and Flavobacterium columnare. Aquacult. Res. 2006, 37, 928–937. [Google Scholar]
- Schrader, K.K. Compounds with inhibitory activity against the channel catfish pathogens Edwardsiella ictaluri and Flavobacterium columnare. N. Am. J. Aquacult. 2008, 70, 147–153. [Google Scholar] [CrossRef]
- Gibbons, S.; Leimkugel, J.; Oluwatuyi, M.; Heinrich, M. Activity of Zanthoxylum clava-herculis extracts against multi-drug resistant methicillin-resistant Staphylococcus aureus (mdr-MRSA). Phytother. Res. 2003, 17, 274–275. [Google Scholar]
- Zuo, G.Y.; Meng, F.Y.; Hao, X.Y.; Zhang, Y.L.; Wang, G.C.; Xu, G.L. Antibacterial alkaloids from Chelidonium majus Linn (Papaveraceae) against clinical isolates of methicillin-resistant Staphylococcus aureus. J. Pharm. Pharmaceut. Sci. 2008, 11, 90–94. [Google Scholar]
- Chung, K.-T.; Stevens, S.E.; Lin, W.-F.; Wei, C.I. Growth inhibition of selected food-borne bacteria by tannic acid, propyl gallate and related compounds. Lett. Appl. Microbiol. 1993, 17, 29–32. [Google Scholar]
- Cai, L.; Wu, C.D. Compounds from Syzygium aromaticum possessing growth inhibitory activity against oral pathogens. J. Nat. Prod. 1996, 59, 987–990. [Google Scholar]
- Thiem, B.; Goślińska, O. Antimicrobial activity of Rubus chamaemorus leaves. Fitoterapia 2004, 75, 93–95. [Google Scholar]
- Shoemaker, C.A.; Olivares-Fuster, O.; Arias, C.R.; Klesius, P.H. Flavobacterium columnare genomovar influences mortality in channel catfish (Ictalurus punctatus). Vet. Microbiol. 2008, 127, 353–359. [Google Scholar]
- Soto, E.; Mauel, M.J.; Karsi, A.; Lawerence, M.L. Genetic and virulence characterization of Flavobacterium columnare from channel catfish (Ictalurus punctatus). J. Appl. Microbiol. 2008, 104, 1302–1310. [Google Scholar]
- Wang, X.K.; Lee, M.J. The extraction and analysis of antimicrobial components of Scutellaria baicalensis. Arch. Sichuan Medical College 1959, 4, 97–98. [Google Scholar]
- Fujita, M.; Shiota, S.; Kuroda, T.; Hatano, T.; Yoshida, T.; Mizushima, T.; Tsuchiya, T. Remarkable synergies between baicalein and tetracycline, and baicalein and beta-lactams against methicillin-resistant Staphylococcus aureus. Microbiol. Immunol. 2005, 49, 391–396. [Google Scholar]
- Saric, M.R. Medicinal Plants of SR Serbia; Serbian Academy of Sciences and Arts: Belgrade, Yugoslavia, 1989. [Google Scholar]
- Petrovic, J.; Stanojkovic, A.; Comic, L.; Curcic, S. Antibacterial activity of Cichorium intybus. Fitoterapia 2004, 75, 737–739. [Google Scholar]
- Kaufman, P.B.; Duke, J.A.; Brielmann, H.; Boik, J.; Hoyt, J.E. A comparative survey of leguminous plants as sources of isoflavones, genistein and daidzein: Implications for human nutrition and health. J. Altern. Complement. Med. 1997, 3, 7–12. [Google Scholar]
- Hong, H.; Landauer, M.R.; Foriska, M.A.; Ledney, G.D. Antibacterial activity of the soy isoflavone genistein. J. Basic Microbiol. 2006, 46, 329–335. [Google Scholar]
- Ulanowska, K.; Tkaczyk, A.; Konopa, G.; Węgrzyn, G. Differential antibacterial activity of genistein arising from global inhibition of DNA, RNA and protein synthesis in some bacterial strains. Arch. Microbiol. 2006, 184, 271–278. [Google Scholar]
- Kim, H.K.; Park, Y.; Kim, H.N.; Choi, B.H.; Jeong, H.G.; Lee, D.G.; Hahm, K.S. Antimicrobial mechanism of β-glycyrrhetinic acid isolated from licorice, Glycyrrhiza glabra. Biotechnol. Lett. 2002, 24, 1899–1902. [Google Scholar]
- Lee, S.K.; Mbwambo, Z.H.; Chung, H.; Luyengi, L.; Gamez, E.J.; Mehta, R.G.; Kinghorn, A.D.; Pezzuto, J.M. Evaluation of the antioxidant potential of natural products. Comb. Chem. High Throughput Screen. 1998, 1, 35–46. [Google Scholar]
- Czarnota, M.A.; Paul, R.N.; Weston, L.A.; Duke, S.O. Anatomy of sorgoleone-secreting root hairs of Sorghum species. Int. J. Plant Sci. 2003, 164, 861–866. [Google Scholar]
- Gimsing, A.L.; Bælum, J.; Dayan, F.E.; Locke, M.; Sejerø, L.H.; Jacobsen, C.S. Mineralization of the allelochemical sorgoleone in soil. Chemosphere 2009, 76, 1041–1047. [Google Scholar]
- Hui, K.M.; Huen, M.S.Y.; Wang, H.Y.; Zheng, H.; Sigel, E.; Baur, R.; Ren, H.; Li, Z.W.; Wong, J.T.-F.; Xue, H. Anxiolytic effect of wogonin, a benzodiazepine receptor ligand isolated from Scutellaria baicalensis Georgi. Biochem. Pharmacol. 2002, 64, 1415–1424. [Google Scholar]
- Ueng, Y.-F.; Shyu, C.-C.; Liu, T.-Y.; Oda, Y.; Lin, Y.-L.; Liao, J.-F.; Chen, C.-F. Protective effects of baicalein and wogonin against benzo[a]pyrene- and alflatoxin B1-induced genotoxicities. Biochem. Pharmacol. 2001, 62, 1653–1660. [Google Scholar]
- Tai, M.C.; Tsang, S.Y.; Chang, L.Y.F.; Xue, H. Therapeutic potential of wogonin: A naturally occurring flavonoid. CNS Drug Rev. 2005, 11, 141–150. [Google Scholar]
- Decostere, A.; Ducatelle, R.; Haesebrouck, F. Flavobacterium columnare (Flexibacter columnaris) associated with severe gill necrosis in koi carp (Cyprinus carpio L). Vet. Res. 2002, 150, 694–705. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Schrader, K.K. Plant Natural Compounds with Antibacterial Activity towards Common Pathogens of Pond-Cultured Channel Catfish (Ictalurus punctatus). Toxins 2010, 2, 1676-1689. https://doi.org/10.3390/toxins2071676
Schrader KK. Plant Natural Compounds with Antibacterial Activity towards Common Pathogens of Pond-Cultured Channel Catfish (Ictalurus punctatus). Toxins. 2010; 2(7):1676-1689. https://doi.org/10.3390/toxins2071676
Chicago/Turabian StyleSchrader, Kevin K. 2010. "Plant Natural Compounds with Antibacterial Activity towards Common Pathogens of Pond-Cultured Channel Catfish (Ictalurus punctatus)" Toxins 2, no. 7: 1676-1689. https://doi.org/10.3390/toxins2071676
APA StyleSchrader, K. K. (2010). Plant Natural Compounds with Antibacterial Activity towards Common Pathogens of Pond-Cultured Channel Catfish (Ictalurus punctatus). Toxins, 2(7), 1676-1689. https://doi.org/10.3390/toxins2071676