Chemical, Physical and Biological Approaches to Prevent Ochratoxin Induced Toxicoses in Humans and Animals


Abstract
:1. Introduction
- (i) prevention of mycotoxin contamination;
- (ii) decontamination or detoxification of foods contaminated with mycotoxins;
- (iii) inhibition of the absorption of consumed mycotoxin in the gastrointestinal tract.
2. Prevention of Mycotoxin Contamination
2.1. Preharvest management
2.2. Postharvest management
2.3. HACCP approaches
3. Decontamination/Detoxification Approaches
- - They must destroy, inactivate, or remove mycotoxins;
- - They must not produce or leave toxic, carcinogenic, or mutagenic residues in the final products or in food products obtained from animals fed by decontaminated feed;
- - They should not adversely affect the desirable physical and sensory properties of the product;
- - They must be capable of destroying fungal spores and the mycelium in order to avoiding mycotoxin formation under favorable conditions;
- - They have to be technically and economically feasible.
3.1. Physical methods

{kind=link}
{kind=link}
| Origin of OTA | Roasting conditions | Reduction in OTA content (%) | References |
|---|---|---|---|
| Inoculation | 200 °C, 10–20 min | 0–12 | [86] |
| Inoculation | 180 °C, 10 min | 31.1 | [94] |
| Natural | 200 ± 5 °C, 20 min | 77–87 | [83] |
| Natural | 200 ± 5 °C, 20 min | 80–90 | [84] |
| Natural | 200 °C, 3 min | 65–100 | [95] |
| Inoculation | 180–240 °C, 5–12 min | 8–98 | [93] |
| Inoculation | 5–6 min, dark roasting | 48–87 | [85] |
| Natural | 5–6 min, dark roasting | 90–100 | [85] |
| Natural | 250 °C, 150 sec | 14–62 | [87] |
| Inoculation | 250 °C, 150 sec | 2–28 | [87] |
| Natural | 223 °C, 4 min | 84 | [88] |
| Natural | Light to dark | 69–96 | [89] |
| Natural | 175–204 °C, 7–9 min | >90 | [90] |
| Inoculation | 200–220 °C, 10–15 min | 22.5–93.9 | [92] |
| Natural | Industrial roasting | 66.5 | [91] |
3.2. Physicochemical methods
3.3. Chemical approaches
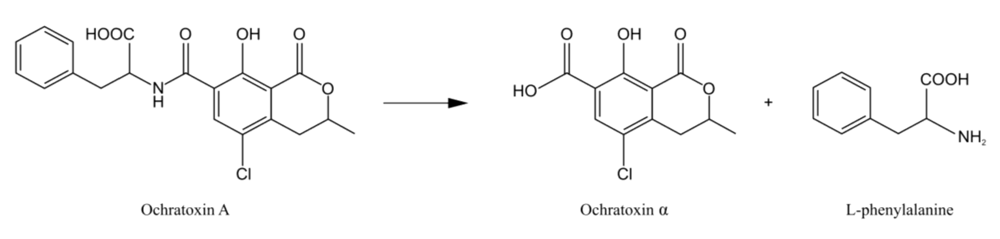
3.4. Microbiological methods

| Microbes or enzymes | Reference |
|---|---|
| Bacteria | |
| Rumen microbes | [149, 175] |
| Butyrivibrio fibrisolvens | [187] |
| Lactobacillus, Streptococcus, Bifidobacterium sp. | [174] |
| Bacillus subtilis, B. licheniformis | [171,188] |
| Acinetobacter calcoaceticus | [164] |
| Phenylobacterium immobile | [189] |
| Nocardia corynebacterioides, Rhodococcus erythropolis, Mycobacterium sp. | [190] |
| Lactobacillus sp. | [191,192] |
| Eubacterium callenderi, E. ramulus, Streptococcus pleomorphus, Lactobacillus vitullinus, Sphingomonas paucimobilis, S. saccharolytica, Stenotrophomonas nitritreducens, Ralstonia eutropha, R. basilensis, Ochrobactrum sp., Agrobacterium sp. | [147,172] |
| Pseudomonas cepacia, P. putida, Rhodococcus erythropolis, Agrobacterium tumefaciens, Comomonas acidovorans | [173] |
| Protozoa | [155,157] |
| Fungi | |
| Aspergillus niger, A. fumigatus | [175] |
| Aspergillus niger, A. versicolor, A. wentii, A. ochraceus | [177] |
| Aspergillus niger, A. japonicus | [178] |
| Pleurotus ostreatus | [193] |
| Saccharomyces cerevisiae | [191,194] |
| Saccharomyces cerevisiae, S. bayanus | [114] |
| Rhizopus stolonifer, R. microsporus, R. homothallicus, R. oryzae | [168] |
| Trichosporon mycotoxinivorans | [195] |
| Phaffia rhodozyma, Xanthophyllomyces dendrorhous | [180] |
| Saccharomyces cerevisiae, Kloeckera apiculata | [169] |
| Aureobasidium pullulans | [39] |
| Cryptococcus flavus, C. laurentii, C. curvatus, C. humicolus, Trichosporon ovoides, T. dulcitum, T. guehoae, T. mucoides, T. coremiiforme, T. cutaneum, T. laibachii, T. monilifotrme, Rhodotorula mucilaginosa, R. fujisanensis | [173,196] |
| Enzymes | |
| Carboxypeptidase A | [158,161] |
| Commercial proteases (Pancreatin from porcine pancreas, Protease A and Prolyve PAC from A. niger) | [163] |
| Commercial hydrolases (Amano A, crude lipase preparation from A. niger) | [162] |
| A. niger hydrolytic metalloenzyme | [197] |

4. Prevention of Toxic Effects of OTA
5. Conclusions
References
- Varga, J.; Rigó, K.; Téren, J.; Mesterházy, Á. Recent advances in ochratoxin research II. Biosynthesis, mode of action and control of ochratoxins. Cereal Res. Commun. 2001, 29, 93–100. [Google Scholar]
- van der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a toxic metabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar]
- Varga, J.; Kocsubé, S.; Péteri, Z.; Samson, R.A. An overview of ochratoxin research. In Applied Mycology; Rai, M., Bridge, P., Eds.; CABI Publishers: London, UK, 2009; pp. 38–55. [Google Scholar]
- Pfohl-Leszkowicz, A.; Manderville, R.A. Ochratoxin A: an overview on toxicity and carcinogenicity in animals and humans. Mol. Nutr. Food Res. 2007, 51, 61–99. [Google Scholar]
- Smith, J.E.; Moss, M.O. Mycotoxins. Formation, Analysis and Significance; John Wiley and Sons: Chichester, UK, 1985; pp. 1–154. [Google Scholar]
- Krogh, P.; Hald, B.; Plestina, R.; Ceovic, S. Balkan (endemic) nephropathy and food–borne ochratoxin A: preliminary results of a survey of foodstuff. Acta Pathol. Microbiol. Scand. B Microbiol. Immunol. 1977, 85, 238–240. [Google Scholar]
- Pfohl-Leszkowicz, A. Ochratoxin A and aristolochic acid involvement in nephropathies and associated urothelial tract tumours. Arh. Hig. Rada. Toksikol. 2009, 60, 465–483. [Google Scholar]
- International Agency for Research on Cancer (IARC). Ochratoxin A. In IARC Monographs on the Evaluation of Carcinogenic Risk of Chemicals to Humans; IARC Press: Lyon, France, 2003; Volume 56, pp. 489–521. [Google Scholar]
- Maaroufi, K.; Zakhama, A.; Baudrimont, I.; Achour, A.; Abid, S.; Ellouz, F.; Dhouib, S.; Creppy, E.E.; Bacha, H. Karyomegaly of tubular cells as early stage marker of the nephrotoxicity induced by ochratoxin A in rats. Hum. Exp. Toxicol. 1999, 18, 410–415. [Google Scholar]
- Wafa, E.W.; Yahya, R.S.; Sobh, M.A.; Eraky, I.; El Baz, H.; El Gayar, H.A.M.; Betbeder, A.M.; Creppy, E.E. Human ochratoxicosis and nephropathy in Egypt: a preliminary study. Hum. Exp. Toxicol. 1998, 17, 124–129. [Google Scholar]
- Schwartz, G.G. Hypothesis: does ochratoxin A cause testicular cancer? Cancer Causes Control 2002, 13, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Kabak, B.; Dobson, A.D.W.; Var, I. Strategies to prevent mycotoxin contamination of food and animal feed: a review. Crit. Rev. Food Sci. Nutr. 2006, 46, 593–619. [Google Scholar]
- Aldred, D.; Magan, N. The use of HACCP in the control of mycotoxins: the case of cereals. In Mycotoxins in Food: Detection and Control; Magan, N., Olsen, M., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 139–173. [Google Scholar]
- Rousseau, J.; Blateyron, L. Ochratoxin A in wines: No curative solution in wine, priority in the vineyard sanitary management. Rev. Oenol. France 2002, 29, 14–16. [Google Scholar]
- Wu, M.T.; Ayres, J.C. Effects of dichlorvos on ochratoxin production by Aspergillus ochraceus. J. Agric. Food Chem. 1974, 22, 536–537. [Google Scholar] [CrossRef] [PubMed]
- Varga, J. University of Szeged, Hungary, 2004; Unpublished work.
- Munimbazi, C.; Saxena, J.; Tsai, W.J.; Bullerman, L.B. Inhibition of production of cyclopiazonic acid and ochratoxin A by the fungicide iprodione. J. Food Prot. 1997, 60, 849–852. [Google Scholar]
- Karadimcheva, B. Effect of some fungicides on the epiphytic microflora in grapes. Gradinar. Lozar. Nauka 1978, 15, 159–166. [Google Scholar]
- Tandon, M.P.; Jamaluddin; Bhargava, V. Chemical control of decay of fruit of Vitis vinifera caused by Aspergillus niger and Penicillium sp. Curr. Sci. 1975, 44, 478. [Google Scholar]
- Lo Curto, R.; Pellicano, T.; Vilasi, F.; Munafo, P.; Dugo, G. Ochratoxin A occurrence in experimental wines in relationship with different pesticide treatments of grapes. Food Chem. 2004, 84, 71–75. [Google Scholar]
- Tjamos, S.E.; Antoniou, P.P.; Kazantzidou, A.; Antonopoulos, D.F.; Papageorgiou, I.; Tjamos, E.C. Aspergillus niger and A. carbonarius in Corinth raisin and wine-producing vineyards in Greece: population composition, ochratoxin A production and chemical control. J. Phytopathol. 2004, 152, 250–255. [Google Scholar] [CrossRef]
- Kappes, M.E.; Serrati, L.; Drouillard, J.B.; Cantus, J.M.; Kazantzidou, M. A crop protection approach to Aspergillus and OTA management in Southern European vineyards. International Workshop on Ochratoxin A in Grapes and Wine: Prevention and Control, Marsala (TP), Italy, 20-21 October 2005; p. 24.
- Bellí, N.; Marín, S.; Sanchis, V.; Ramos, A.J. Impact of fungicides on Aspergillus carbonarius growth and ochratoxin A production on synthetic grape-like medium and on grapes. Food Addit. Contam. 2006, 23, 1021–1029. [Google Scholar]
- Chernin, L.; Brandis, A.; Ismailov, Z.; Chet, I. Pyrrolnitrin production by an Enterobacter agglomerans strain with a broad spectrum of antagonistic activity towards fungal and bacterial phytopathogens. Curr. Microbiol. 1996, 32, 208–212. [Google Scholar]
- Molot, B.; Solanet, D. Ochratoxine: prévention du risque. In Etude au vignoble de fongicides actifs contre Aspergillus carbonarius incidences sur la présence aux vendanges, Les Entretiens Viti-Vinicoles Rhone-Mediterranée, Nimes; 2003; pp. 18–21. [Google Scholar]
- Visconti, A.; Perrone, G.; Cozzi, G.; Solfrizzo, M. Managing ochratoxin A risk in the grape-wine food chain. Food Addit. Contam. 2008, 25, 193–202. [Google Scholar]
- Battilani, P.; Pietri, A.; Bertuzzi, T.; Languasco, L.; Giorni, P.; Kozakiewicz, Z. Occurrence of ochratoxin A-producing fungi in grapes grown in Italy. J. Food Prot. 2003, 66, 633–636. [Google Scholar]
- Medina, A.; Mateo, R.; Valle-Algarra, F.M.; Mateo, E.M.; Jimenez, M. Effect of carbendazim and physicochemical factors on the growth and ochratoxin A production of Aspergillus carbonarius isolated from grapes. Int. J. Food Microbiol. 2007, 119, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Jimenez, M.; Mateo, R.; Magan, N. Efficacy of natamycin for control of growth and ochratoxin A production by Aspergillus carbonarius strains under different environmental conditions. J. Appl. Microbiol. 2007, 103, 2234–2239. [Google Scholar]
- Favilla, M.; Pascale, M.; Ricelli, A.; Evidente, A.; Amalfitano, C.; Altomare, C. Inhibition of species of the Aspergillus section Nigri and ochratoxin A production in grapes by fusapyrone. Appl. Environ. Microbiol. 2008, 74, 2248–2253. [Google Scholar]
- Cozzi, G.; Pascale, M.; Perrone, G.; Visconti, A.; Logrieco, A. Effect of Lobesia botrana damages on black aspergilli rot and ochratoxin A content in grapes. Int. J. Food Microbiol. 2006, 111S1, S88–S92. [Google Scholar]
- Rousseau, J. Ochratoxin A in wines: current knowledge. Vinidea.net Wine Internet Technical J. 2004, No. 5. Available online: http://www.icv.fr/kiosqueuk/refs/vinideaOTAenglish1.pdf (accessed on 28 April 2006).
- Merrien, O. Influence de différents facteurs sor l’OTA dans vins et prévention du risqué d’apparition. Conference Européenne, Direction de la Santé et de la Protection du Consommateur. 3rd Forum on ochratoxin A, Brussels, Belgium; 2003. [Google Scholar]
- Bae, S.; Fleet, G.H.; Heard, G.M. Occurrence and significance of Bacillus thuringiensis on wine grapes. Int. J. Food Microbiol. 2004, 94, 301–312. [Google Scholar]
- Vega, F.E.; Mercadier, G. Insects, coffee and ochratoxin A. Florida Entomol. 1998, 81, 543–544. [Google Scholar]
- Arrieta, G.; Hemandez, A.; Espinoza, A.M. Diversity of Bacillus thuringiensis strains isolated from coffee plantations infested with the coffee berry borer Hypothenemus hampei. Rev. Biol. Trop. 2004, 52, 757–764. [Google Scholar] [PubMed]
- Damon, A. A review of the biology and control of the coffee berry borer, Hypothenemus hampei (Coleoptera: Scolytidae). Bull. Entomol. Res. 2000, 90, 453–465. [Google Scholar]
- Vega, F.E.; Posada, F.; Gianfagna, T.J.; Chaves, F.C.; Peterson, S.W. An insect parasitoid carrying an ochratoxin producing fungus. Naturwissenschaften 2006, 93, 297–299. [Google Scholar]
- de Felice, D.V.; Solfrizzo, M.; De Curtis, F.; Lima, G.; Visconti, A.; Castoria, R. Strains of Aureobasidium pullulans can lower ochratoxin A contamination in wine grapes. Phytopathology 2008, 98, 1261–1270. [Google Scholar]
- Zahavi, T.; Cohen, L.; Weiss, B.; Schena, L.; Daus, A.; Kaplunov, T.; Zutkhi, J.; Ben-Arie, R.; Droby, S. Biological control of Botrytis, Aspergillus and Rhizopus rots on table and wine grapes in Israel. Postharvest Biol. Technol. 2000, 20, 115–124. [Google Scholar]
- Bleve, G.; Grieco, F.; Cozzi, G.; Logrieco, A.; Visconti, A. Isolation of epiphytic yeast with potential for biocontrol of Aspergillus carbonarius and A. niger on grape. Int. J. Food Microbiol. 2006, 108, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Munimbazi, C.; Bullerman, L.B. Isolation and partial characterization of antifungal metabolites of Bacillus pumilus. J. Appl. Microbiol. 1998, 84, 959–968. [Google Scholar]
- Valero, A.; Oliván, S.; Marín, S.; Sanchis, V.; Ramos, A.J. Effect of intra- and interspecific interaction on OTA production by Aspergillus section Nigri in grapes during dehydration. Food Microbiol. 2007, 24, 254–259. [Google Scholar]
- Podile, A.R.; Prakash, A.P. Lysis and biological control of Aspergillus niger by Bacillus subtilis AF 1. Can. J. Microbiol. 1996, 42, 533–538. [Google Scholar]
- Chelkowski, J.; Dopierala, G.; Godlewska, B.; Radomyska, W.; Szebiotko, K. Mycotoxins in cereal grain. Part III; Production of ochratoxin A in different varieties of wheat, rye and barley. Food 1981, 25, 625–629. [Google Scholar] [PubMed]
- Battilani, P.; Logrieco, A.; Giorni, P.; Cozzi, G.; Bertuzzi, T.; Pietri, A. Ochratoxin A production by Aspergillus carbonarius on some grape varieties grown in Italy. J. Sci. Food Agric. 2004, 84, 1736–1740. [Google Scholar]
- Silva, M.C.; Varzea, V.; Guerra-Guimaraes, L.; Azinheira, H.G.; Fernandez, D.; Petitot, A.S.; Bertrand, B.; Lashermes, P.; Nicole, M. Coffee resistance to the main diseases: leaf rust and coffee berry disease. Braz. J. Plant Physiol. 2006, 18, 119–147. [Google Scholar]
- Irulandi, S.; Rajendran, R.; Samuel Stephen, D. Screening of coffee genotypes/cultivars for resistance to berry borer, Hypothenemus hampei (Ferrari) (Scolytidae: Coleoptera) in the field. Pest Management Horticult. Ecosyst. 2007, 13, 34–45. [Google Scholar]
- Varga, J.; Tóth, B. Strategies to control mycotoxins in feeds: a review. Acta Vet. Hung. 2005, 53, 189–203. [Google Scholar]
- Scudamore, K.A. Prevention of ochratoxin A in commodities and likely effects of processing fractionation and animal feeds. Food Addit. Contam. Part A 2005, 22, 17–25. [Google Scholar]
- Castellucci, F. Code of sound vitivinicultural practices in order to minimise levels of ochratoxin A in vine-based products. Resolution VITI-OENO 1/2005; OIV: Paris, France, 2005; pp. 1–5. Available online: http://news.reseau-concept.net/images/oiv_uk/Client/VITI-OENO_1-2005_EN.pdf (accessed on 20 February 2010).
- Kozakiewicz, Z.; Battilani, P.; Cabanes, J.; Venancio, A.; Mule, G.; Tjamos, E.; Lichter, A.; Magan, N.; Sanchis, V.; Lebrihi, A.; Zinzani, G.; Minguez, S. Making wine safer: the case of ochratoxin A. In Meeting the Mycotoxin Menace; van Egmond, H., van Osenbruggen, T., Lopez-Garcia, R., Barug, D., Visconti, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2003; pp. 131–140. [Google Scholar]
- Suarez-Quiroz, M.L.; Gonzalez-Rios, O.; Barel, M.; Guyot, B.; Schorr-Galindo, S.; Guiraud, J.P. Effect of chemical and environmental factors on Aspergillus ochraceus growth and toxigenesis in green coffee. Food Microbiol. 2004, 21, 629–634. [Google Scholar]
- Abramson, D. Mycotoxin formation and environmental factors. In Mycotoxins in Agriculture and Food Safety; Sinha, K.K., Bhatnagar, D., Eds.; Marcel Dekker: New York, NY, USA, 1998; pp. 255–277. [Google Scholar]
- Arroyo, M.; Aldred, D.; Magan, N. Environmental factors and weak organic acid interactions have differential effects on control of growth and ochratoxin A production by Penicillium verrucosum isolates in bread. Int. J. Food Microbiol. 2005, 98, 223–231. [Google Scholar]
- Marin, S.; Guynot, M.E.; Neira, P.; Bernado, M.; Sanchis, V.; Ramos, A.J. Risk assessment of the use of sub-optimal levels of weak acid preservatives in the control of mold growth on bakery products. Int. J. Food Microbiol. 2002, 79, 203–211. [Google Scholar]
- Tong, C.H.; Draughon, F.A. Inhibition by antimicrobial food additives of ochratoxin A production by Aspergillus sulphureus and Penicillium viridicatum. Appl. Environ. Microbiol. 1985, 49, 1407–1411. [Google Scholar]
- Barberis, C.; Astoreca, A.; Fernandez-Juri, G.; Chulze, S.; Dalcero, A.; Magnoli, C. Use of propyl-paraben to control growth and ochratoxin A production by Aspergillus section Nigri onpeanut meal extract agar. Int. J. Food. Microbiol. 2009, 136, 133–136. [Google Scholar]
- Palumbo, J.D.; O’Keeffe, T.L.; Mahoney, N.E. Inhibition of ochratoxin A production and growth of Aspergillus species by phenolic antioxidant compounds. Mycopathologia 2007, 164, 241–248. [Google Scholar]
- Aroyeun, S.O.; Adegoke, G.O. Reduction of ochratoxin A (OTA) in spiked cocoa powder and beverage using aqueous extracts and essential oils of Aframomum danielli. Afr. J. Biotechnol. 2007, 6, 612–616. [Google Scholar]
- Nguefack, J.; Nguikwie, S.K.; Fotio, D.; Dongmo, B.; Amvam Zollo, P.H.; Leth, V. Fungicidal potential of essential oils and fractions from Cymbopogon citratus, Ocimum gratissimum and Thymus vulgaris to control Alternaria padwickii and Bipolaris oryzae, two seed-borne fungi of rice (Oryza Sativa L.). J. Essent. Oil Res. 2007, 19, 581–587. [Google Scholar]
- Magan, N.; Aldred, D. Postharvest control strategies: minimizing mycotoxins in the food chain. Int. J. Food Microbiol. 2007, 119, 131–139. [Google Scholar]
- Solimar, K.M.; Badeaa, R.I. Effect of oil extracted from some medicinal plants on different mycotoxigenic fungi. Food Chem. Toxicol. 2002, 40, 1669–1675. [Google Scholar]
- Basilico, M.Z.; Basılico, J.C. Inhibitory effects of some spice essential oils on Aspergillus ochraceus NRRL3174 growth and ochratoxin A production. Lett. Appl. Microbiol. 1999, 29, 238–241. [Google Scholar]
- Juglal, S.; Govinden, R.; Odhav, B. Spice oils for the control of co-occurring mycotoxin-producing fungi. J. Food Prot. 2002, 65, 683–687. [Google Scholar]
- Romero, S.M.; Alberto, M.R.; de Nadra, M.C.M.; Vaamonde, G. Inhibition of growth and ochratoxin A biosynthesis in Aspergillus carbonarius by flavonoid and nonflavonoid compounds. Mycotoxin Res. 2009, 25, 165–170. [Google Scholar]
- Lee, S.E.; Park, B.S.; Bayman, P.; Baker, J.L.; Choi, W.S.; Campbell, B.C. Suppression of ochratoxin biosynthesis by naturally occurring alkaloids. Food Addit. Contam. Part A 2007, 24, 391–397. [Google Scholar]
- Schnürer, J.; Magnusson, J. Antifungal lactic acid bacteria as biopreservatives. Trends Food Sci. Technol. 2005, 16, 70–78. [Google Scholar]
- Cairns-Fuller, V.; Aldred, D.; Magan, N. Water, temperature and gas composition interactions affect growth and ochratoxin A production by isolates of Penicillium verrucosum on wheat grain. J. Appl. Microbiol. 2005, 99, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Paster, N.; Lisker, N.; Chet, I. Ochratoxin production by Aspergillus ochraceus Wilhem grown under controlled atmospheres. Appl. Environ. Microbiol. 1983, 45, 1136–1139. [Google Scholar]
- Avissar, I.; Marinansky, R.; Pesis, E. Postharvest decay control of grape by acetaldehyde vapors. ISHS Acta Horticult. 1989, 258, 655–660. [Google Scholar]
- Aziz, N.H.; Moussa, L.A.A.; Far, F.M.E. Reduction of fungi and mycotoxins formation in seeds by gamma-radiation. J. Food Safety 2004, 24, 109–127. [Google Scholar]
- Kostecki, M.; Golinski, P.; Uchman, W.; Grabarkiewicz-Szczesna, J. Decomposition of ochratoxin A by heat and gamma-irradiation. IARC Sci. Publ. 1991, 115, 109–111. [Google Scholar]
- Meissner, R.; Huser-Hahn, I.; Wieczorek, K. Method for reducing afla- and ochratoxin contamination in cereals, nuts, fruits and spices. Patent number WO/2009/068213, 2009. [Google Scholar]
- Magan, N. Mycotoxin contamination of food in Europe: early detection and prevention strategies. Mycopathologia 2006, 162, 245–253. [Google Scholar]
- Frank, M. HACCP and its mycotoxin control potential: an evaluation of ochratoxin A in coffee production. Third Joint FAO/WHO/UNEP International Conference on Mycotoxins, Tunis, 3–6 March1999; p. 34.
- Lopez-Garcia, R.; Mallmann, C.A.; Pineiro, M. Design and implementation of an integrated management system for ochratoxin A in the coffee production chain. Food Addit. Contam. 2008, 25, 231–240. [Google Scholar]
- Krogh, P.; Hald, B.; Giersten, P.; Myken, F. Fate of ochratoxin A and citrinin during malting and brewing experiments. Appl. Microbiol. 1974, 28, 31–34. [Google Scholar]
- Scott, P.M. Effects of processing and detoxification treatments on ochratoxin A: introduction. Food Addit. Contam. 1996, 13, 19–21, (Suppl.).. [Google Scholar] [PubMed]
- Rotter, R.G.; Marquardt, R.R.; Frohlich, A.A.; Abramson, D. Ensiling as a means of reducing ochratoxin A concentrations in contaminated barley. J. Sci. Food Agric. 1990, 50, 155–160. [Google Scholar]
- Osborne, B.G.; Ibe, F.; Brown, G.L.; Petagine, F.; Scudamore, K.A.; Banks, J.N.; Hetminski, M.T.; Leonard, C.T. The effects of milling and processing on wheat contaminated with ochratoxin A. Food Addit. Contam. 1996, 13, 141–153. [Google Scholar]
- Boudra, H.; Le Bars, P.; Le Bars, J. Thermostability of ochratoxin A in wheat under two moisture conditions. Appl. Environ. Microbiol. 1995, 61, 1156–1158. [Google Scholar]
- Levi, C.P.; Trenk, H.L.; Mohr, H.K. Study of the occurrence of ochratoxin A in green coffee beans. J. Assoc. Off. Anal. Chem. Int. 1974, 57, 866–870. [Google Scholar]
- Gallaz, L.; Stalder, R. Ochratoxin A in coffee. J. Microbiol. Technol. Chem. 1976, 4, 147–149. [Google Scholar]
- Micco, C.; Grossi, M.; Miraglia, M.; Brera, C.A. Study of the contamination by ochratoxin A of green and roasted coffee beans. Food Addit. Contam. 1989, 6, 333–339. [Google Scholar]
- Tsubouchi, H.; Yamamoto, K.; Hisada, K.; Sakabe, Y.; Udagawa, S. Effect of roasting on ochratoxin A level in green coffee beans inoculated with Aspergillus ochraceus. Mycopathologia 1987, 97, 111–115. [Google Scholar]
- Studer-Rohr, I.; Dietrich, D.R.; Schlatter, J.; Schlatter, C. Ochratoxin A in coffee: new evidence and toxicology. Lebensm. Technol. 1994, 27, 435–441. [Google Scholar]
- Blanc, M.; Pittet, A.; Munoz-Boksz, R.; Viani, R. Behavior of ochratoxin A during green coffee roasting and soluble coffee manufacture. J. Agric. Food Chem. 1998, 46, 673–675. [Google Scholar]
- Van der Stegen, G.H.D.; Essens, P.J.M.; van der Lijn, J. Effect of roasting conditions on reduction of ochratoxin A in coffee. J. Agric. Food Chem. 2001, 49, 4713–4715. [Google Scholar]
- Romani, S.; Pinnavaia, G.G.; Dalla Rosa, M. Influence of roasting levels on ochratoxin A content in coffee. J. Agric. Food Chem. 2003, 51, 5168–5171. [Google Scholar]
- Perez de Obanos, A.; Gonzalez-Penas, E.; Lopez de Cerain, A. Influence of roasting and brew preparation on the ochratoxin A content in coffee infusion. Food Addit. Contam. 2005, 22, 463–471. [Google Scholar]
- Urbano, G.R.; de Freitas Leitao, M.F.; Vicentini, M.C.; Taniwaki, M.H. Preliminary studies on the destruction of ochratoxin A in coffee during roasting. Food Sci. Technol. Int. 2004, 10, 45–49. [Google Scholar]
- Ferraz, M.B.M.; Farah, A.; Iamanaka, B.T.; Perrone, D.; Copetti, M.V.; Marques, V.X.; Vitali, A.A.; Taniwaki, M.H. Kinetics of ochratoxin A destruction during coffee roasting. Food Control 2010, 21, 872–877. [Google Scholar]
- Nehad, E.A.; Farag, M.M.; Kawther, M.S.; Abdel-Samed, A.K.; Naguib, K. Stability of ochratoxin A (OTA) during processing and decaffeination in commercial roasted coffee beans. Food Addit. Contam. 2005, 22, 761–767. [Google Scholar]
- La Pera, L.; Avellone, G.; Lo Turco, V.; Di Bella, G.; Agozzino, P.; Dugo, G. Influence of roasting and different brewing processes on the ochratoxin A content in coffee determined by high-performance liquid chromatography-fluorescence detection (HPLC-FLD). Food Addit. Contam. Part A 2008, 25, 1257–1263. [Google Scholar]
- Wilkens, J.; Jörissen, U. Degradation of ochratoxin A during manufacture of roasted coffee. 21st Mycotoxin Workshop, Jena, Germany; 1999. [Google Scholar]
- Heilmann, W.; Rehfeldt, A.G.; Rotzoll, F. Behaviour and reduction of ochratoxin A in green coffee beans in response to various processing methods. Eur. Food Res. Technol. 1999, 209, 297–300. [Google Scholar]
- Deberghes, P.; Betbeder, A.M.; Boisard, F.; Blanc, R.; Delaby, J.F.; Krivobok, S.; Steiman, R.; Seigle-Murandi, F.; Creppy, E.E. Detoxification of ochratoxin A, a food contaminant: prevention of growth of Aspergillus ochraceus and its production of ochratoxin A. Mycotoxin Res. 1995, 11, 37–47. [Google Scholar]
- Deberghes, P.; Deffieux, G.; Gharbi, A.; Betbeder, A.M.; Boisard, F.; Blanc, R.; Delaby, J.F.; Creppy, E.E. Detoxification de l’ochratoxine A par des moyens physiques, chimiques et enzymatiques. Hum. Ochratoxicosis Pathol. 1993, 231, 75–82. [Google Scholar]
- Refai, M.K.; Aziz, N.H.; El-Far, F.; Hassan, A.A. Detection of ochratoxin produced by Aspergillus ochraceus in feedstuffs and its control by γ-radiation. Appl. Radiat. Isot. 1996, 47, 617–621. [Google Scholar]
- Gambuti, A.; Strollo, D.; Genovese, A.; Ugliano, M.; Ritieni, A.; Moio, L. Influence of enological practices on ochratoxin A concentration in wine. Am. J. Enol. Viticult. 2005, 56, 155–162. [Google Scholar]
- Solfrizzo, M.; Avantaggiato, G.; Panzarini, G.; Visconti, A. Removal of ochratoxin A from contaminated red wines by repassage over grape pomaces. J. Agric. Food Chem. 2010, 58, 317–323. [Google Scholar]
- Lindner, W. Decontamination and detoxification of cereals contaminated with mycotoxins. US Patent number 5498431, 1996. [Google Scholar]
- Lindner, W. Decontamination and detoxication of cereals contaminated by mycotoxins. Patent number WO9216116, 1992. [Google Scholar]
- Castellari, M.; Versari, A.; Fabiani, A.; Parpinello, G.P.; Galassi, S. Removal of ochratoxin A in red wines by means of adsorption treatments with commercial fining agents. J. Agric. Food Chem. 2001, 49, 3917–3921. [Google Scholar]
- Dumeau, F.; Trione, D. Influence of different treatments on the concentration of ochratoxin A in red wines. Rev. Oenol. France 2000, 27, 37–38. [Google Scholar]
- Savino, M.; Limosani, P.; Garcia-Moruno, E. Reduction of ochratoxin A contamination in red wines by oak wood fragments. Am. J. Enol. Viticult. 2007, 58, 97–101. [Google Scholar]
- Olivares-Marín, M.; Del Prete, V.; Garcia-Moruno, E.; Fernández-González, C.; Macías-García, A.; Gómez-Serrano, A. The development of an activated carbon from cherry stones and its use in the removal of ochratoxin A from red wine. Food Control 2009, 20, 298–303. [Google Scholar]
- Belajová, E.; Rauová, D.; Daško, L. Retention of ochratoxin A and fumonisin B1 and B2 from beer on solid surfaces: comparison of efficiency of adsorbents with different origin. Eur. Food Res. Technol. 2007, 224, 301–308. [Google Scholar]
- Kurtbay, H.M.; Bekci, Z.; Merdivan, M.; Yurdakoc, K. Reduction of ochratoxin a levels in red wine by bentonite, modified bentonites, and chitosan. J. Agric. Food Chem. 2008, 56, 2541–2545. [Google Scholar]
- Var, I.; Kabak, B.; Erginkaya, Z. Reduction in ochratoxin A levels in white wine, following treatment with activated carbon and sodium bentonite. Food Control 2008, 19, 592–598. [Google Scholar]
- Del Prete, V.; Rodriguez, H.; Carrascosa, A.V.; De Las Rivas , B.; Garcia-Moruno, E.; Munoz, R. In vitro removal of ochratoxin A by wine lactic acid bacteria. J. Food Prot. 2007, 70, 2155–2160. [Google Scholar]
- Mateo, E.M.; Medina, A.; Mateo, R.; Jiménez, M. Effect of ethanol on the ability of Oenococcus oeni to remove ochratoxin A in synthetic wine-like media. Food Control 2010, 21, 935–941. [Google Scholar]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Ochratoxin A removal in synthetic and natural grape juices by selected oenological Saccharomyces strains. J. Appl. Microbiol. 2004, 97, 1038–1044. [Google Scholar]
- Petersson, S.; Schnurer, J. Biocontrol of mold growth in high-moisture wheat stored under airtight conditions by Pichia anomala, Pichia guilliermondii, and Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1995, 61, 1027–1032. [Google Scholar]
- Moruno, E.G.; Sanlorenzo, C.; Boccaccino, B.; Di Stefano, R. Treatment with yeast to reduce the concentration of ochratoxin A in red wine. Am. J. Enol. Vitic. 2005, 56, 73–76. [Google Scholar]
- Caridi, A.; Galvano, F.; Tafuri, A.; Ritieni, A. Ochratoxin A removal during winemaking. Enzyme Microb. Technol. 2006, 40, 122–126. [Google Scholar]
- Cecchini, F.; Morassut, M.; Garcia Moruno, E.; Di Stefano, R. Influence of yeast strain on ochratoxin A content during fermentation of white and red must. Food Microbiol. 2006, 23, 411–417. [Google Scholar]
- Meca, G.; Blaiotta, G.; Ritieni, A. Reduction of ochratoxin A during the fermentation of Italian red wine Moscato. Food Control 2010, 21, 579–583. [Google Scholar]
- Raju, M.V.L.N.; Devegowda, G. Esterified-glucomannan in broiler chicken diets-contaminated with aflatoxin, ochratoxin and T-2 toxin: Evaluation of its binding ability (in vitro) and efficacy as immunomodulator. Asian-Australas. J. Anim. Sci. 2002, 15, 1051–1056. [Google Scholar]
- Raju, M.V.L.N.; Devegowda, G. Influence of esterified-glucomannan on performance and organ morphology, serum biochemistry and haematology in broilers exposed to individual and combined mycotoxicosis (aflatoxin, ochratoxin and T-2 toxin). Br. Poult. Sci. 2000, 41, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Ringot, D.; Lerzy, B.; Bonhoure, J.P.; Auclair, E.; Oriol, E.; Larondelle, Y. Effect of temperature on in vitro ochratoxin A biosorption onto yeast cell wall derivatives. Proc. Biochem. 2005, 40, 3008–3016. [Google Scholar]
- Ringot, D.; Lerzy, B.; Chaplain, K.; Bonhoure, J.P.; Auclair, E.; Larondelle, Y. In vitro biosorption of ochratoxin A on the yeast industry by-products: Comparison of isotherm models. Biores. Technol. 2007, 98, 1812–1821. [Google Scholar]
- Espejo, F.J.; Armada, S. Effect of activated carbon on ochratoxin A reduction in “Pedro Ximenez” sweet wine made from off-vine dried grapes. Eur. Food Res. Toxicol. 2009, 229, 255–622. [Google Scholar]
- Huwig, A.; Freimund, S.; Kappeli, O.; Dutler, H. Mycotoxin detoxication of animal feed by different adsorbents. Toxicol. Lett. 2001, 122, 179–188. [Google Scholar]
- Tangni, E.K.; Simonis, J.; Larondelle, Y.; De Meeus, D.A.L. Decontaminating and detoxifying liquid food media, especially beer, using insoluble vegetable fibers to adsorb mycotoxins. Patent number WO2005007794-A1, 2005. [Google Scholar]
- Dakovic, A.; Tomaševic-Canovic, M.; Rottinghaus, G.E.; Dondur, V.; Mašic, A. Adsorption of ochratoxin A on octadecyldimethyl benzyl ammonium exchanged-clinoptilolite-heulandite tuff. Coll. Surf. B 2003, 30, 157–165. [Google Scholar]
- Dakovic, A.; Tomaševic-Canovic, M.; Dondur, V.; Rottinghaus, G.E.; Medakovic, V.; Zaric, S. Adsorption of mycotoxins by organozeolites. Coll. Surf. B 2005, 46, 20–25. [Google Scholar]
- Amezqueta, S.; Golzales-Penas, E.; Murillo-Arbizu, M.; Lopez de Cerain, A. Ochratoxin A decontamination: a review. Food Control 2009, 20, 326–333. [Google Scholar]
- Tomasevic-Canovic, M.; Dakovic, A.; Rottinghaus, G.E.; Matijasevic, S.; Duricicc, M. Surfactant modified zeolites-new efficient adsorbents for mycotoxins. Microporous Mesoporous Mater. 2003, 61, 173–180. [Google Scholar]
- Thimm, N.; Schwaighofer, B.; Ottner, F.; Fröschl, H.; Greifenender, S.; Binder, E.M. Adsorption of Mycotoxins. Mycotoxin Res. 2001, 17A, 219–223. [Google Scholar]
- Chelkowski, J.; Szebiotko, K.; Golinski, P.; Buchowski, M.; Godlewska, B.; Radomyrska, W.; Wiewiorowska, M. Mycotoxins in cereal grain. 5; Changes of cereal grain biological value after ammoniation and mycotoxins (ochratoxins) inactivation. Nahrung 1982, 26, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Peraica, M.; Domijan, A.M.; Jurjevic, Z.; Cvjetkovic, B. Prevention of exposure to mycotoxins from food and feed. Arh. Hig. Rada. Toxikol. 2002, 53, 229–237. [Google Scholar]
- Madsen, A.; Hald, B.; Mortensen, H.P. Feeding experiments with ochratoxin A contaminated barley to bacon pigs 3. Detoxification by ammoniation heating + NaOH, or autoclaving. Acta Agric. Scand. 1983, 33, 171–175. [Google Scholar] [CrossRef]
- Kabak, B.; Dobson, A.D.W. Biological strategies to counteract the effects of mycotoxins. J. Food Prot. 2009, 72, 2006–2016. [Google Scholar]
- Dimanche, P.; Kane, A. Senegal’s confectionery peanut supply chain: the challenge of controlling aflatoxin levels. In Food Safety Management in Developing Countries. Proceedings of the International Workshop, CIRAD-FAO, Montpellier, France; Hanak, E., Boutrif, E., Fabre, P., Pineiro, M., Eds.; CIRAD-FAO, 2002; pp. 1–4. [Google Scholar]
- Kabak, B. Prevention and management of mycotoxins in food and feed. In Mycotoxins in Food,Feed and Bioweapons; Rai, M., Varma, A., Eds.; Springer-Verlag: Berlin, Heidelberg, Germany, 2009; pp. 201–227. [Google Scholar]
- Bortoli, G.; Fabian, M. A process to remove mycotoxins from green coffee. ASIC Congress: Trieste, 2001. Available online: http://www.demus.it/download/remove_mycotoxins.pdf (accessed on 20 February, 2010).
- Bortoli, G.; Fabian, M. A process to remove mycotoxins from green coffee. Patent number PCT/EP 97/02014 (1997/04/22), 1997. [Google Scholar]
- Amezqueta, S.; Gonzalez-Penas, E.; Lizarraga, T.; Murillo-Arbizu, M.; Lopez de Cerain, A. A simple chemical method reduces ochratoxin A in contaminated cocoa shells. J. Food Prot. 2008, 71, 1422–1426. [Google Scholar]
- Denvir, A.J.; Mckenzie, K.S.; Rogers, T.D.; Miller, D.R.; Hitchens, G.D.; Andrews, C.C. Method of food decontamination by treatment with ozone. US Patent number 6171625, 2001. [Google Scholar]
- McKenzie, K.S.; Sarr, A.B.; Mayura, K.; Bailey, R.H.; Miller, D.R.; Rogers, T.D.; Norred, W.P.; Voss, K.A.; Plattner, R.D.; Kubena, L.F.; Phillips, T.D. Oxidative degradation and detoxification of mycotoxins using a novel source of ozone. Food Chem. Toxicol. 1997, 35, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Castegnaro, M.; Barek, J.; Fremy, J.M.; Lafontaine, M.; Miraglia, M.; Sansone, E.G.; Telling, G.M. Laboratory Decontamination and Destruction of Carcinogens in Laboratory Wastes: Some Mycotoxins; International Agency for Research on Cancer: Lyon, France, 1991; pp. 1–80. [Google Scholar]
- Fouler, S.G.; Trivedi, A.B.; Kitabatake, N. Detoxification of citrinin and ochratoxin A by hydrogen peroxide. J. Assoc. Off. Anal. Chem. Int. 1994, 77, 631–637. [Google Scholar]
- Sweeney, M.J.; Dobson, A.D.W. Mycotoxin production by Aspergillus, Fusarium and Penicillium species. Int. J. Food Microbiol. 1998, 43, 141–158. [Google Scholar]
- Halász, A.; Lásztity, R.; Abonyi, T.; Bata, Á. Decontamination of mycotoxin-containing food and feed by biodegradation. Food Rev. Int. 2009, 25, 284–298. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Zehner, F.; Taubel, M.; Schatzmayr, D.; Klimitsch, A.; Loibner, A.P.; Binder, E.M. Microbiologicals for deactivating mycotoxins. Mol. Nutr. Food Res. 2006, 50, 543–551. [Google Scholar]
- Galtier, P.; Alvinerie, M. In vitro transformation of ochratoxin A by animal microbial floras. Ann. Vet. Res. 1976, 7, 91–98. [Google Scholar]
- Hult, K.; Teiling, A.; Gatenbeck, S. Degradation of ochratoxin A by a ruminant. Appl. Environ. Microbiol. 1976, 32, 443–444. [Google Scholar]
- Galtier, P. Contribution of pharmacokinetic studies to mycotoxicology-ochratoxin A. Vet. Sci. Commun. 1978, 1, 349–358. [Google Scholar]
- Madhyastha, M.S.; Marquardt, R.R.; Frohlich, A.A. Hydrolysis of ochratoxin A by the microbial activity of digesta in the gastrointestinal tract of rats. Arch. Environ. Contam. Toxicol. 1992, 23, 468–472. [Google Scholar]
- Suzuki, S.; Satoh, T.; Yamazaki, M. The pharmacokinetics of ochratoxin A in rats. Jpn. J. Pharmacol. 1977, 27, 735–744. [Google Scholar]
- Hansen, C.E.; Dueland, S.; Drevon, C.A.; Stormer, F.C. Metabolism of ochratoxin A by primary cultures of rat hepatocytes. Appl. Environ. Microbiol. 1982, 43, 1267–1271. [Google Scholar]
- Stormer, F.C.; Storen, O.; Hansen, C.E.; Pedersen, J.I.; Aasen, A.J. Formation of (4R)- and (4S)-4-hydroxyochratoxin A and 10-hydroxyochratoxin A from ochratoxin A by rabbit liver microsomes. Appl. Environ. Microbiol. 1983, 45, 1183–1187. [Google Scholar]
- Kiessling, K.-H.; Pettersson, H.; Sandholm, K.; Olsen, M. Metabolism of aflatoxin, ochratoxin, zearalenon, and three trichothecenes by intact rumen fluid, rumen protozoa, and rumen bacteria. Appl. Environ. Microbiol. 1984, 47, 1070–1073. [Google Scholar]
- Moroi, K.; Suzuki, S.; Kuga, T.; Yamazaki, M.; Kanisawa, M. Reduction of ochratoxin A toxicity in mice treated with phenylalanine and phenobarbital. Toxicol. Lett. 1985, 25, 1–5. [Google Scholar]
- Ozpinar, H.; Bilal, T.; Abas, I.; Kutay, C. Degradation of ochratoxin A in rumen fluid in vitro. Med. Biol. 2002, 9, 66–69. [Google Scholar]
- Pitout, M.J. The hydrolysis of ochratoxin A by some proteolytic enzymes. Biochem. Pharmacol. 1969, 18, 485–491. [Google Scholar]
- Akiyama, H.; Toyoda, M.; Kato, M.; Igimi, S.; Kumagai, S. The degradation of several mycotoxins by human intestinal microflora cultured by continuous flow culture system. Mycotoxins 1997, 44, 21–27. [Google Scholar]
- Doster, R.C.; Sinnhuber, R.O. Comparative rates of hydrolysis of ochratoxins A and B in vitro. Food Cosm. Toxicol. 1972, 10, 389–394. [Google Scholar]
- Stander, M.A.; Steyn, P.S.; van der Westhuizen, F.H.; Payne, B.E. A kinetic study into the hydrolysis of the ochratoxins and analogs by carboxypeptidase A. Chem. Res. Toxicol. 2001, 14, 302–304. [Google Scholar]
- Stander, M.A.; Bornscheurer, U.T.; Henke, E.; Steyn, P.S. Screening of commercial hydrolases for the degradation of ochratoxin A. J. Agric. Food Chem. 2000, 48, 5736–5739. [Google Scholar]
- Abrunhosa, L.; Santos, L.; Venancio, A. Degradation of ochratoxin A by proteases and by a crude enzyme of Aspergillus niger. Food Biotechnol. 2006, 20, 231–242. [Google Scholar]
- Hwang, C.-A.; Draughon, F.A. Degradation of ochratoxin A by Acinetobacter calcoaceticus. J. Food Prot. 1994, 57, 410–414. [Google Scholar]
- Yamazaki, M.; Suzuki, S.; Sakakibara, Y.; Miyaki, K. The toxicity of 5-chloro-8-hydroxy-3,4-dihydro-3-methyl-isocoumarin-7-carboxylic acid, a hydrolyzate of ochratoxin A. Jpn. J. Med. Sci. Biol. 1971, 24, 245–250. [Google Scholar]
- Bruinink, A.; Rasonyi, T.; Sidler, C. Differences in neurotoxic effects of ochratoxin A, ochracin and ochratoxin-alpha in vitro. Nat. Toxins 1998, 6, 173–177. [Google Scholar]
- Follmann, W.; Hillebrand, I.E.; Creppy, E.E.; Bolt, H.M. Sister chromatid exchange frequency in cultured isolated porcine urinary bladder epithelial cells (PUBEC) treated with ochratoxin A and alpha. Arch. Toxicol. 1995, 69, 280–286. [Google Scholar]
- Varga, J.; Péteri, Z.; Tábori, K.; Téren, J.; Vágvölgyi, C. Degradation of ochratoxin A and other mycotoxins by Rhizopus isolates. Int. J. Food Microbiol. 2005, 99, 321–328. [Google Scholar]
- Angioni, A.; Caboni, P.; Garau, A.; Farris, A.; Orro, D.; Budroni, M.; Cabras, P. In vitro interaction between ochratoxin A and different strains of Saccharomyces cerevisiae and Kloeckera apiculata. J. Agric. Food Chem. 2007, 55, 2043–2048. [Google Scholar]
- Silva, A.; Galli, R.; Grazioli, B. Metodi di riduzione di residui di ocratossina nei vini. Ind. BeVande 2003, 32, 467–472. [Google Scholar]
- Petchkongkaew, A.; Taillandier, P.; Gasaluck, P.; Lebrihi, A. Isolation of Bacillus spp. from Thai fermented soybean (Thua-nao): screening for aflatoxin B1 and ochratoxin A detoxification. J. Appl. Microbiol. 2008, 104, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Schatzmayr, G.; Heidler, D.; Fuchs, E.; Loibner, A.P.; Braun, R.; Binder, E.M. Evidence of ochratoxin A-detoxification ctivity of rumen fluid, intestinal fluid and soil samples as well as isolation of relevant microorganisms from these environments. Mycotoxin Res. 2002, 18, 183–187, (Suppl. 2).. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Heidler, D.; Fuchs, E.; Binder, E.M. Microorganisms for biological detoxification of mycotoxins,namely ochratoxins and/or zearalenons, as well as method and use thereof. US Patent number 2009/0098244 A1, 2009. [Google Scholar]
- Skrinjar, M.; Rasic, J.L.; Stojicic, V. Lowering ochratoxin A level in milk by yoghurt bacteria and bifidobacteria. Folia Microbiol. 1996, 41, 26–28. [Google Scholar]
- Varga, J.; Rigó, K.; Téren, J. Degradation of ochratoxin A by Aspergillus species. Int. J. Food Microbiol. 2000, 59, 1–7. [Google Scholar]
- Péteri, Z. Examination of mycotoxin detoxification ability of microscopical fungi. Ph.D. Thesis, University of Szeged, Szeged, Hungary, 2010. [Google Scholar]
- Abrunhosa, L.; Serra, R.; Venancio, A. Biodegradation of ochratoxin A by fungi isolated from grapes. J. Agric. Food Chem. 2002, 50, 7493–7496. [Google Scholar]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Biodegradation of ochratoxin A by Aspergillus section Nigri species isolated from French grapes: a potential means of ochratoxin A decontamination in grape juices and musts. FEMS Microbiol. Lett. 2006, 255, 203–208. [Google Scholar]
- Stander, M.; Steyn, P.S.; Smit, M.S. Metabolic degradation of ochratoxin A by certain yeasts. Patent number WO02099142 A3, 2002. [Google Scholar]
- Péteri, Z.; Téren, J.; Vágvölgyi, C.; Varga, J. Ochratoxin degradation and adsorption caused by astaxanthin-producing yeasts. Food Microbiol. 2007, 24, 205–210. [Google Scholar]
- Ruhland, M.; Engelhardt, G.; Wallnofer, P.R. Transformation of the mycotoxin ochratoxin A in plants. 2. Time course and rates of degradation and metabolite production in cell-suspension cultures of different crop plants. Mycopathologia 1996, 134, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Ruhland, M.; Engelhardt, G.; Schafer, W.; Wallnofer, P.R. Transformation of the mycotoxin ochratoxin A in plants: 1. Isolation and identification of metabolites formed in cell suspension cultures of wheat and maize. Nat. Toxins 1996, 4, 254–260. [Google Scholar] [PubMed]
- Ruhland, M.; Engelhardt, G.; Wallnofer, P.R.; Schafer, W. Transformation of the mycotoxin ochratoxin A in wheat and maize cell suspension cultures. Naturwissenschaften 1994, 81, 453–454. [Google Scholar]
- Karlovsky, P. Biological detoxification of fungal toxins and its use in plant breeding, feed and food production. Nat. Toxins 1999, 7, 1–23. [Google Scholar]
- Wright, W.R.; Harein, P.K. Effects of some mycotoxins on the Mediterranean flour moth. Environ. Entomol. 1982, 11, 1043–1045. [Google Scholar]
- European Food Safety Authority (EFSA). Review of mycotoxin-detoxifying agents used as feed additives: mode of action, efficacy and feed/food safety. 2009. Available online: http://www.efsa.europa.eu/en/scdocs/doc/22e.pdf (accessed on 28 January 2010).
- Westlake, K.; Mackie, R.I.; Dutton, M.F. Effects of several mycotoxins on specific growth rate of Butyrivibrio fibrisolvens and toxin degradation in vitro. Appl. Environ. Microbiol. 1987, 53, 613–614. [Google Scholar]
- Böhm, J.; Grajewski, J.; Asperger, H.; Cecon, B.; Rabus, B.; Razzazi, E. Study on biodegradation of some A- and B-trichothecenes and ochratoxin A by use of probiotic microorganisms. Mycotoxin Res. 2000, 16A, 70–74. [Google Scholar]
- Wegst, W.; Lingens, F. Bacterial degradation of ochratoxin A. FEMS Microbiol. Lett. 1983, 17, 341–344. [Google Scholar]
- Holzapfel, W.; Brost, I.; Farber, P.; Geisen, R.; Bresch, H.; Jany, K.D.; Mengu, M.; Jakobsen, M.; Steyn, P.S.; Teniola, D.; Addo, P. Actinomycetes for breaking down aflatoxin B1, ochratoxin A, and/or zearalenon. Patent number WO02/099142 A3, 2002. [Google Scholar]
- Piotrowska, M.; Zakowska, Z. The biodegradation of ochratoxin A in food products by lactic acid bacteria and baker’s yeast. In Food Biotechnology; Bielecki, S., Tramper , J., Polak, J, Eds.; Elsevier: Amsterdam, The Netherlands, 2000; pp. 307–310. [Google Scholar]
- Fuchs, S.; Sontag, G.; Stidl, R.; Ehrlich, V.; Kundi, M.; Knasmüller, S. Detoxification of patulin and ochratoxin A, two abundant mycotoxins, by lactic acid bacteria. Food Chem. Toxicol. 2008, 46, 1398–1407. [Google Scholar]
- Engelhardt, G. Degradation of ochratoxin A and B by the white rot fungus Pleurotus ostreatus. Mycotoxin Res. 2002, 18, 37–43. [Google Scholar]
- Scott, P.M.; Kanhere, S.R.; Lawrence, G.A.; Daley, E.F.; Farber, J.M. Fermentation of wort containing added ochratoxin A and fumonisins B1 and B2. Food Addit. Contam. 1995, 12, 31–40. [Google Scholar] [PubMed]
- Molnar, O.; Schatzmayr, G.; Fuchs, E.; Prillinger, H. Trichosporon mycotoxinivorans sp. nov., a new yeast species useful in biological detoxification of various mycotoxins. Syst. Appl. Microbiol. 2004, 27, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Schatzmayr, G.; Heidler, D.; Fuchs, E.; Nitsch, S.; Mohnl, M.; Tiubel, M.; Loibner, A.P.; Braun, R.; Binder, E.M. Investigation of different yeast strains for the detoxification of ochratoxin A. Mycotoxin Res. 2003, 19, 124–128. [Google Scholar]
- Abrunhosa, L.; Venancio, A. Isolation and purification of an enzyme hydrolyzing ochratoxin A from Aspergillus niger. Biotechnol. Lett. 2007, 29, 1909–1914. [Google Scholar]
- Haubeck, H.D.; Lorkowski, G.; Kolsch, E.; Roschenthaler, R. Immunosuppression by ochratoxin A and its prevention by phenylalanine. Appl. Environ. Microbiol. 1981, 41, 1040–1042. [Google Scholar]
- Bruinink, A.; Sidler, C. The neurotoxic effects of ochratoxin A are reduced by protein binding but are not affected by l-phenylalanine. Toxicol. Appl. Pharmacol. 1997, 146, 173–179. [Google Scholar]
- Stoev, S.D. Studies on some feed additives and materials giving partial protection against the suppressive effect of ochratoxin A on egg production of laying hens. Res. Vet. Sci. 2010, 88, 486–491. [Google Scholar]
- Stoev, S.D.; Djuvinov, D.; Mirtcheva, T.; Pavlov, D.; Mantle, P. Studies on some feed additives giving partial protection against ochratoxin A toxicity in chicks. Toxicol. Lett. 2002, 135, 33–50. [Google Scholar]
- El Adlouni, C.; Pinelli, E.; Azemar, B.; Zaoui, D.; Beaune, P.; Pfohl-Leszkowicz, A. Role of CYP 2C and microsomal glutathione-S-transferase in modulating susceptibility to ochratoxin A genotoxicity. Environ. Mol. Mutagen. 2000, 35, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.E.; Baudrimont, I.; Betbeter, A.M. How aspartame prevents the toxicity of ochratoxin A. J. Toxicol. Sci. 1998, 23, 165–172, (Suppl. 2).. [Google Scholar] [PubMed]
- Baudrimont, I.; Betbeder, A.M.; Creppy, E.E. Reduction of the ochratoxin A-induced cytotoxicity in Vero cells by aspartame. Arch. Toxicol. 1997, 71, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Phillips, T.D.; Kubena, L.F.; Harvey, R.B.; Taylor, D.R.; Heidelbaugh, N.D. Hydrated sodium calcium aluminosilicate: a high affinity sorbent for aflatoxin. Poult. Sci. 1988, 67, 243–247. [Google Scholar]
- Santin, E.; Maiorka, A.; Krabbe, E.L.; Paulillo, A.C.; Alessi, A.C. Effect of hydrated sodium calcium aluminosilicate on the prevention of the toxic effects of ochratoxin. J. Appl. Poult. Res. 2002, 11, 22–28. [Google Scholar]
- Denli, M.; Blandon, J.C.; Guynot, M.E.; Salado, S.; Perez, J.F. Efficacy of a new ochratoxin-binding agent (OcraTox) to counteract the deleterious effects of ochratoxin A in laying hens. Poult. Sci. 2008, 87, 2266–2272. [Google Scholar]
- Galvano, F.; Pietri, A.; Bertuzzi, T.; Piva, A.; Chies, L.; Galvano, M. Activated carbons: in vitro affinity for ochratoxin A and deoxynivalenol and relation of adsorption ability to physicochemical parameters. J. Food Prot. 1998, 61, 469–475. [Google Scholar]
- Rotter, R.G.; Frohlich, A.A.; Marquardt, R.R. Influence of dietary charcoal on ochratoxin A toxicity in Leghorn chicks. Can. J. Vet. Res. 1989, 53, 449–453. [Google Scholar]
- Avantaggiato, G.; Havenaar, R.; Visconti, A. Assessment of the multimycotoxin-binding efficacy of a carbon/aluminosilicate-based product in an in vitro gastrointestinal model. J. Agric. Food Chem. 2007, 55, 4810–4819. [Google Scholar]
- Aoudia, N.; Callu, P.; Grosjean, F.; Larondelle, Y. Effectiveness of mycotoxin sequestration activity of micronized wheat fibres on distribution of ochratoxin A in plasma, liver and kidney of piglets fed a naturally contaminated diet. Food Chem. Toxicol. 2009, 47, 1485–1489. [Google Scholar]
- Aravind, K.L.; Patil, V.S.; Devegowda, G.; Umakantha, B.; Ganpule, S.P. Efficacy of esterified glucomannan to counteract mycotoxicosis in naturally contaminated feed on performance and serum biochemical and hematological parameters in broilers. Poult. Sci. 2003, 82, 571–576. [Google Scholar]
- Bursian, S.J.; Mitchell, R.R.; Yamini, B.; Fitzgerald, S.D.; Murphy, P.A.; Fernandez, G.; Rottinghaus, G.E.; Moran, L.; Leefers, K.; Choi, I. Efficacy of a commercial mycotoxin binder in alleviating effects of ochratoxin A, fumonisin B1, moniliformin and zearalenone in adult mink. Vet. Hum. Toxicol. 2004, 46, 122–129. [Google Scholar]
- Turbic, A.; Ahokas, J.T.; Haskard, C.A. Selective in vitro binding of dietary mutagens, individually or in combination, by lactic acid bacteria. Food Addit. Contam. 2002, 19, 144–152. [Google Scholar]
- Kerkadi, A.; Barriault, C.; Marquardt, R.R.; Frohlich, A.A.; Yousef, I.M.; Zhu, X.X.; Tuchweber, B. Cholestyramine protection against ochratoxin A toxicity: role of ochratoxin A sorption by the resin and bile acid enterohepatic circulation. J. Food Prot. 1999, 62, 1461–1465. [Google Scholar]
- Obrecht-Pflumio, S.; Grosse, Y.; Pfohl-Leszkowicz, A.; Dirheimer, G. Protection by indomethacin and aspirin against genotoxicity of ochratoxin A, particularly in the urinary bladder and kidney. Arch. Toxicol. 1996, 70, 244–248. [Google Scholar]
- Hoehler, D.; Marquardt, R.R. Influence of vitamins E and C on the toxic effects of ochratoxin A and T-2 toxin in chicks. Poult. Sci. 1996, 75, 1508–1515. [Google Scholar]
- Grosse, Y.; Chekir-Ghedira, L.; Huc, A.; Obrecht-Pflumio, S.; Dirheimer, G.; Bacha, H.; Pfohl-Leszkowicz, A. Retinol, ascorbic acid and alpha-tocopherol prevent DNA adduct formation in mice treated with the mycotoxins ochratoxin A and zearalenone. Cancer Lett. 1997, 114, 225–229. [Google Scholar]
- Abdel-Wahhab, M.A.; Abdel-Galil, M.M.; El-Lithey, M. Melatonin counteracts oxidative stress in rats fed with an ochratoxin A contaminated diet. J. Pineal Res. 2005, 38, 130–135. [Google Scholar]
- Schaaf, G.J.; Nijmeijer, S.M.; Mass, R.F.M.; Roestenberg, P.; de Groene, E.M.; Fink-Gremmels, J. The role of oxidative stress in the ochratoxin A-Mediated toxicity in proximal tubular cells. Biochem. Biophys. Acta 2002, 1588, 149–158. [Google Scholar]
- Renzulli, C.; Galvano, F.; Piredomenico, L.; Speroni, E.; Guerra, M.C. Effects of Rosmarinic acid against aflatoxin B1 and ochratoxin A-induced cell damage in a human hepatoma cell line (Hep G2). J. Appl. Toxicol. 2004, 24, 289–296. [Google Scholar]
- Baldi, A.; Losio, M.N.; Cheli, F.; Rebucci, R.; Sangalli, L.; Fusi, E.; Bertasi, B.; Pavoni, E.; Carli, S.; Politis, I. Evaluation of the protective effects of alpha-tocopherol and retinol against ochratoxin A cytotoxicity. Br. J. Nutr. 2004, 91, 507–512. [Google Scholar]
- Guerra, M.C.; Galvano, F.; Bonsi, L.; Speroni, E.; Costa, S.; Renzulli, C.; Cervellati, R. Cyanidin-3-O-β-glucopyranoside, a natural free-radical scavenger against aflatoxin B1 and ochratoxin A-induced cell damage in a human hepatoma cell line (Hep G2) and a human colonic adenocarcinoma cell line (CaCo2). Br. J. Nutr. 2005, 94, 211–220. [Google Scholar]
- Kumari, N.; Kumar, P.; Mitra, D.; Prasad, B.; Tiwary, B.N.; Varshney, L. Effects of ionizing radiation on microbial decontamination, phenolic contents, and antioxidant properties of triphala. J Food Sci. 2009, 74, 109–113. [Google Scholar]
- Ibeh, I.N.; Saxena, D.K. Effect of a-tocopherol supplementation on the impact of aflatoxin B1 on the testes of rats. Exp. Toxicol. Pathol. 1998, 50, 221–224. [Google Scholar]
- Abdel-Wahhab, M.A.; Nada, A.S.; Arbid, M.S. Ochratoxicosis: prevention of developmental toxicity by L-methionine in rats. J. Appl. Toxicol. 1999, 19, 7–12. [Google Scholar]
- Costa, S.; Utan, A.; Cervellati, R.; Speroni, E.; Guerra, M.C. Catechins: Natural free-radical scavengers against ochratoxin A-induced cell damage in a pig kidney cell line (LLC-PK1). Food Chem. Toxicol. 2007, 45, 1910–1917. [Google Scholar]
- Atroshi, F.; Biese, I.; Saloniemi, H.; Ali-Vehmas, T.; Saari, S.; Rizzo, A.; Veijalainen, P. Significance of apoptosis and its relationship to antioxidants after ochratoxin A administration in mice. J. Pharm. Pharmaceut. Sci. 2000, 3, 281–291. [Google Scholar]
- Lee, S.E.; Park, B.S.; Bayman, P.; Baker, J.L.; Choi, W.S.; Campbell, B.C. Suppression of ochratoxin biosynthesis by naturally occurring alkaloids. Food Addit. Contam. Part A 2007, 24, 391–397. [Google Scholar]
- Verma, R.; Chakraborty, D. Emblica officinalis aqueous extract ameliorates ochratoxin-induced lipid peroxidation in the testis of mice. Acta Pol. Pharm. 2008, 65, 187–194. [Google Scholar]
- Pfohl-Leszkowicz, A.; Bartsch, H.; Azemar, B.; Mohr, U.; Esteve, J.; Castegnaro, M. MESNA protects rats against nephrotoxicity but not carconigenicity induced by ochratoxin A, implicating two separate pathways. Fact. Univ. Ser. Med. Biol. 2002, 9, 37–43. [Google Scholar]
- Baudrimont, I.; Betbeder, A.M.; Gharbi, A.; Pfohl-Leszkowicz, A.; Dirheimer, G.; Creppy, E.E. Effect of superoxide dismutase and catalase on the nephrotoxicity induced by subchronical administration of ochratoxin A in rats. Toxicology 1994, 89, 101–111. [Google Scholar]
- Biernasiak, J.; Piotrowska, M.; Libudzisz, Z. Detoxification of mycotoxins by probiotic preparation for broiler chickens. Mycotoxin Res. 2006, 22, 230–235. [Google Scholar]
- Politis, I.; Fegeros, K.; Nitsch, S.; Schatzmayr, G.; Kantas, D. Use of Trichosporon mycotoxinivorans to suppress the effects of ochratoxicosis on the immune system of broiler chicks. Br. Poult. Sci. 2005, 46, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Schatzmayr, G.; Schatzmayr, D.; Starkl, V.; Nitsch, S.; Forat, M.; Binder, E. A novel concept for simultaneous deactivation of various mycotoxinsin piglets. J. Anim. Sci. 2005, 83, 236, (Suppl. 1).. [Google Scholar]
- Hanif, N.Q.; Muhammad, G.; Siddique, M.; Khanum, A.; Ahmed, T.; Gadahai, J.A.; Kaukab, G. Clinico-pathomorphological, serum biochemical and histological studies in broilers fed ochratoxin A and a toxin deactivator (Mycofix ® Plus). Br. Poult. Sci. 2008, 49, 632–642. [Google Scholar]
- Karaman, M.; Basmacioglu, H.; Ortatatli, M.; Oguz, H. Evaluation of the detoxifying effect of yeast glucomannan on aflatoxicosis in broilers as assessed by gross examination and histopathology. Br. Poult. Sci. 2005, 46, 394–400. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Varga, J.; Kocsubé, S.; Péteri, Z.; Vágvölgyi, C.; Tóth, B. Chemical, Physical and Biological Approaches to Prevent Ochratoxin Induced Toxicoses in Humans and Animals. Toxins 2010, 2, 1718-1750. https://doi.org/10.3390/toxins2071718
Varga J, Kocsubé S, Péteri Z, Vágvölgyi C, Tóth B. Chemical, Physical and Biological Approaches to Prevent Ochratoxin Induced Toxicoses in Humans and Animals. Toxins. 2010; 2(7):1718-1750. https://doi.org/10.3390/toxins2071718
Chicago/Turabian StyleVarga, János, Sándor Kocsubé, Zsanett Péteri, Csaba Vágvölgyi, and Beáta Tóth. 2010. "Chemical, Physical and Biological Approaches to Prevent Ochratoxin Induced Toxicoses in Humans and Animals" Toxins 2, no. 7: 1718-1750. https://doi.org/10.3390/toxins2071718
APA StyleVarga, J., Kocsubé, S., Péteri, Z., Vágvölgyi, C., & Tóth, B. (2010). Chemical, Physical and Biological Approaches to Prevent Ochratoxin Induced Toxicoses in Humans and Animals. Toxins, 2(7), 1718-1750. https://doi.org/10.3390/toxins2071718