Spatial Patterns of Aflatoxin Levels in Relation to Ear-Feeding Insect Damage in Pre-Harvest Corn


Abstract
:1. Introduction

2. Results and Discussion
2.1. Results
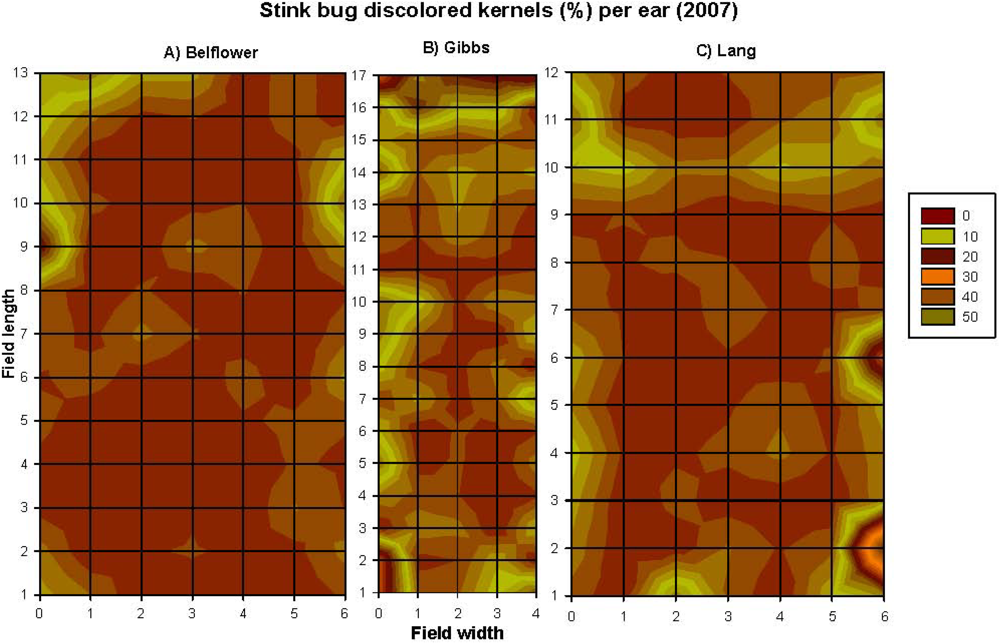
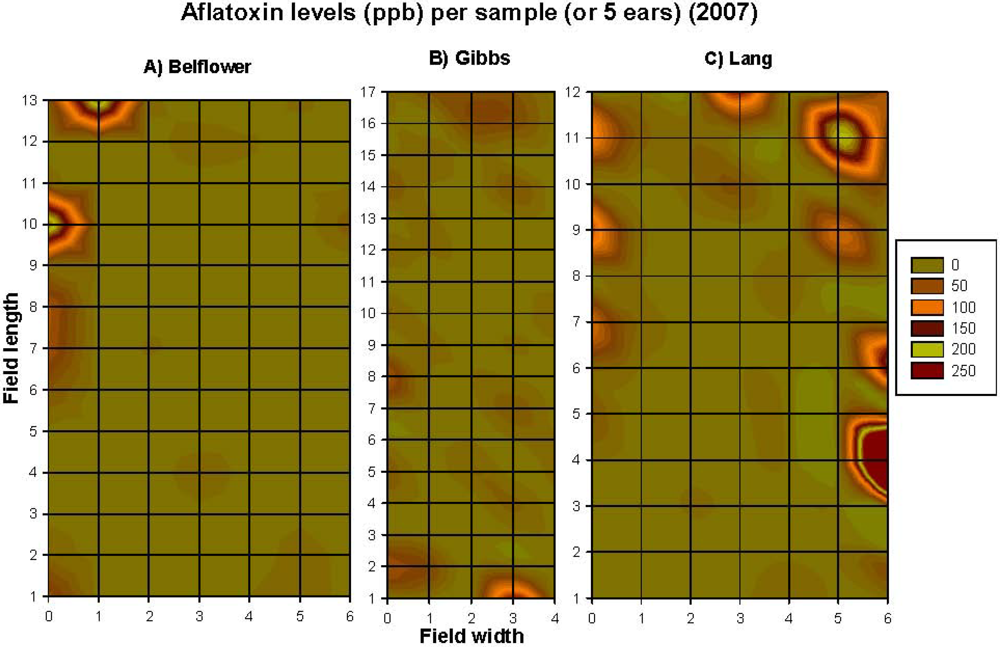
2.1.1. Grid-Sampling Data of 2006 Data


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Husk Coverage | Discolored Kernels | Maize Weevil | Aflatoxin (ppb) | |
|---|---|---|---|---|
| Corn earworm | −0.09 | −0.17 | 0.08 | 0.1 |
| 0.46 | 0.15 | 0.49 | 0.37 | |
| Husk coverage | 0.12 | −0.01 | −0.12 | |
| 0.31 | 0.93 | 0.29 | ||
| Stink bug- | 0.05 | 0.36 | ||
| discolored kernels | 0.68 | 0.001 | ||
| Maize weevil | 0.25 | |||
| 0.03 |
2.1.2. Grid-Sampling Data of 2007
| Husk Coverage | Discolored Kernels | Maize Weevil | Aflatoxin (ppb) | |
|---|---|---|---|---|
| Corn earworm | −0.13 | 0.16 | 0.28 | 0.14 |
| 0.04 | 0.009 | 0.0001 | 0.02 | |
| Husk coverage | −0.08 | −0.11 | 0.01 | |
| 0.2 | 0.09 | 0.84 | ||
| Stink bug- | 0.14 | 0.18 | ||
| Discolored kernels | 0.02 | 0.003 | ||
| Maize weevil | 0.19 | |||
| 0.002 |



2.2. Discussion
3. Experimental Section
3.1. Fields
3.2. Corn Sampling
3.3. Insect and Associated Damage Sampling
3.4. Aflatoxin Quantification
3.5. Experimental Design and Data Analysis
4. Conclusions and Further Research Directions
Acknowledgments
Supplementary Files
References
- Council for Agricultural Science and Technology, Mycotoxins: Economic and Health Risks. Task Force Report No. 116; Council for Agricultural Science and Technology.: Ames, IA, USA, 1989; p. 91.
- Wu, F.; Munkvold, G.P. Mycotoxins in ethanol co-products: Modeling economic impacts on the livestock industry and management strategies. J. Agric. Food Chem. 2008, 56, 3900–3911. [Google Scholar]
- Payne, G.A. Aflatoxin in maize. Crit. Rev. Plant Sci. 1992, 10, 423–440. [Google Scholar]
- Widstrom, N.W. Aflatoxin in developing maize:In teractions among involved Biota and pertinent econiche factors. In Handbook of Applied Mycology; Bhatnagar, D., Lillehoj, E.B., Arora, D.K., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1992; Volume 5, pp. 23–58. [Google Scholar]
- Widstrom, N.W. The aflatoxin problem with corn grain. Adv. Agron. 1996, 56, 219–280. [Google Scholar]
- Scully, B.T.; Krakowsky, M.D.; Ni, X.; Wilson, J.P.; Lee, R.D.; Guo, B. Preharvest aflatoxin contamination of corn and other grain crops grown on the U.S. Southeastern Coastal Plain. Toxin Rev. 2009, 28, 169–179. [Google Scholar] [CrossRef]
- Widstrom, N.W.; Sparks, A.N.; Lillehoj, E.B.; Kwolek, W.F. Aflatoxin production and lepidopteran insect injury on corn in Georgia. J. Econ. Entomol. 1975, 68, 855–856. [Google Scholar]
- Widstrom, N.W. The role of insects and other plant pests in aflatoxin contamination of corn, cotton, and peanuts—A review. J. Environ. Qual. 1979, 8, 5–11. [Google Scholar] [CrossRef]
- Barry, D.; Zuber, M.S.; Lillehoj, E.B.; McMillian, W.W.; Adams, N.J.; Kwolek, W.F.; Widstrom, N.W. Evaluation of two arthropod vectors as inoculators of developing maize ears with Aspergillus flavus. Environ. Entomol. 1985, 14, 634–636. [Google Scholar]
- Barry, D.; Lillehoj, E.B.; Widstrom, N.W.; McMillian, W.W.; Zuber, M.S.; Kwolek, W.F.; Guthrie, W.D. Effect of husk tightness and insect (Lepidoptera) infestation on aflatoxin contamination of preharvest maize. Environ. Entomol. 1986, 15, 1116–1118. [Google Scholar]
- Widstrom, N.W.; Wilson, D.M.; McMillian, W.W. Aflatoxin contamination of preharvest corn as influenced by timing and method of inoculation. Appl. Environ. Microbiol. 1981, 42, 249–251. [Google Scholar]
- Zummo, N.; Scott, G.E. Evaluation of field inoculation techniques for screening maize genotypes against kernel infection by Aspergillus flavus. Mississippi Plant Dis. 1989, 73, 313–316. [Google Scholar]
- Zuber, M.S.; Calvert, O.H.; Kwolek, W.F.; Lillehoj, E.B.; Kang, M.S. Aflatoxin B1 production in an eight-line diallel of Zea mays infected with Aspergillus flavus. Phytopathology 1978, 68, 1346–1349. [Google Scholar] [CrossRef]
- Widstrom, N.W.; Wilson, D.M.; McMillian, W.W. Ear resistance of maize inbreds to field aflatoxin contamination. Crop Sci. 1984, 24, 1155–1157. [Google Scholar]
- Widstrom, N.W.; McMillian, W.W.; Wilson, D.M. Segregation for resistance to aflatoxin contamination among seeds on an ear of hybrid maize. Crop Sci. 1987, 27, 961–963. [Google Scholar]
- Campbell, K.W.; White, D.G. Inheritance of resistance to Aspergillus ear rot and aflatoxin in corn genotypes. Phytopathology 1995, 85, 886–896. [Google Scholar] [CrossRef]
- Williams, W.P.; Windham, G.L. Registration of maize germplasm line Mp715. Crop Sci. 2001, 41, 1374–1375. [Google Scholar]
- Widstrom, N.W.; Lillehoj, E.B.; Sparks, A.N.; Kwolek, W.F. Corn earworm damage and aflatoxin B1 on corn ears protected with insecticide. J. Econ. Entomol. 1976, 69, 677–679. [Google Scholar]
- Dowd, P.F. Involvement of arthropods in the establishment of mycotoxigenic fungi under field conditions. In Mycotoxins in Agriculture and Food Safety; Sinha, K.K., Bhatnagar, D., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1998; pp. 307–350. [Google Scholar]
- Brown, S.L.; Lee, R.D. Effect of planting date, variety and degree of ear maturation on the colonization of field corn by maize weevils (Coleoptera: Curculionidae). J. Entomol. Sci. 2002, 37, 137–142. [Google Scholar]
- Widstrom, N.W.; Guo, B.Z.; Wilson, D.M. Integration of crop management and genetics for control of preharvest aflatoxin contamination of corn. J. Toxicol. Toxin Rev. 2003, 22, 199–227. [Google Scholar]
- Dowd, P.F. Insect management to facilitate preharvest mycotoxin management. J. Toxicol.-Toxin Rev. 2003, 22, 327–350. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ni, X.; Wilson, J.P.; Buntin, G.D.; Guo, B.; Krakowsky, M.D.; Lee, R.D.; Cottrell, T.E.; Scully, B.T.; Huffaker, A.; Schmelz, E.A. Spatial Patterns of Aflatoxin Levels in Relation to Ear-Feeding Insect Damage in Pre-Harvest Corn. Toxins 2011, 3, 920-931. https://doi.org/10.3390/toxins3070920
Ni X, Wilson JP, Buntin GD, Guo B, Krakowsky MD, Lee RD, Cottrell TE, Scully BT, Huffaker A, Schmelz EA. Spatial Patterns of Aflatoxin Levels in Relation to Ear-Feeding Insect Damage in Pre-Harvest Corn. Toxins. 2011; 3(7):920-931. https://doi.org/10.3390/toxins3070920
Chicago/Turabian StyleNi, Xinzhi, Jeffrey P. Wilson, G. David Buntin, Baozhu Guo, Matthew D. Krakowsky, R. Dewey Lee, Ted E. Cottrell, Brian T. Scully, Alisa Huffaker, and Eric A. Schmelz. 2011. "Spatial Patterns of Aflatoxin Levels in Relation to Ear-Feeding Insect Damage in Pre-Harvest Corn" Toxins 3, no. 7: 920-931. https://doi.org/10.3390/toxins3070920
APA StyleNi, X., Wilson, J. P., Buntin, G. D., Guo, B., Krakowsky, M. D., Lee, R. D., Cottrell, T. E., Scully, B. T., Huffaker, A., & Schmelz, E. A. (2011). Spatial Patterns of Aflatoxin Levels in Relation to Ear-Feeding Insect Damage in Pre-Harvest Corn. Toxins, 3(7), 920-931. https://doi.org/10.3390/toxins3070920