Molecular Analysis of the Interaction of the Snake Venom Rhodocytin with the Platelet Receptor CLEC-2


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Protein Interfaces, Surfaces and Assemblies
2.2. Molecular Dynamics
3. Results and Discussion
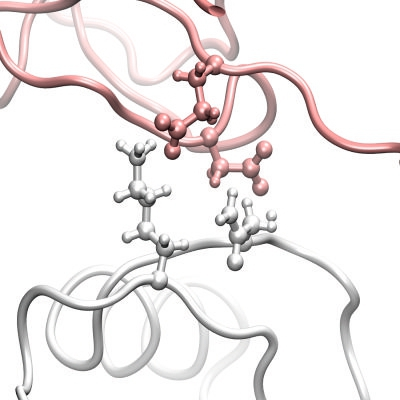
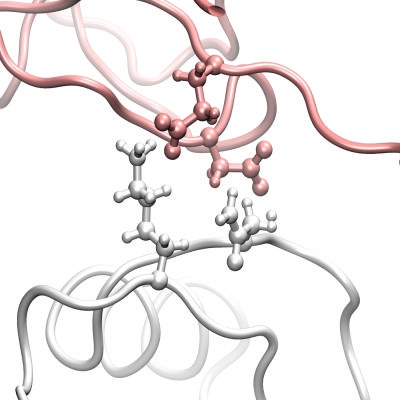
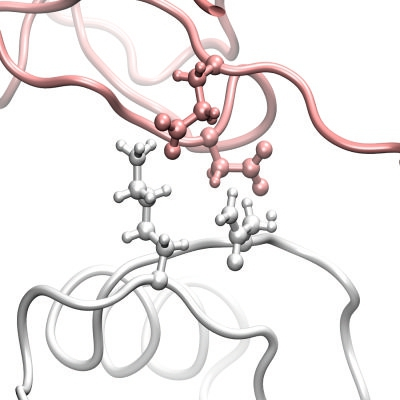
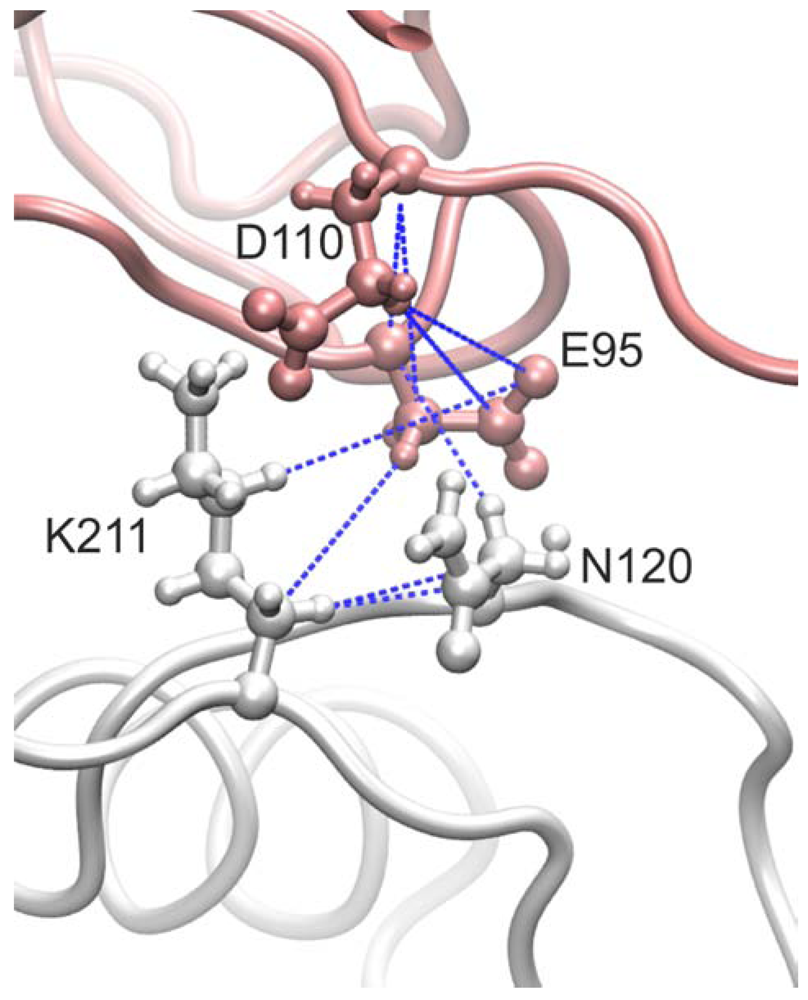
3.1. The Interaction Surfaces of the Rhodocytin-CLEC-2 Interaction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 |
 |

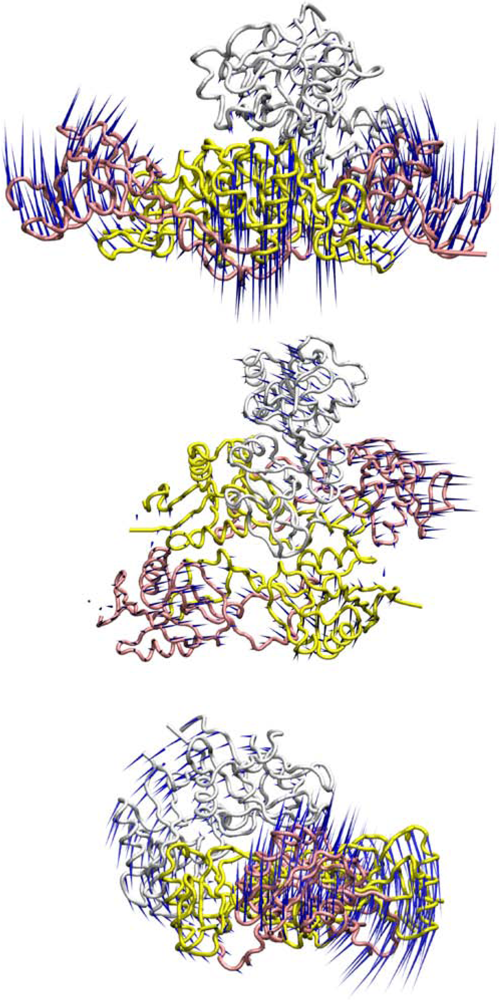
3.2. Dynamic Flexibility of the Rhodocytin-CLEC-2 Complexes


4. Conclusions
Acknowledgments
Conflict of Interest
References
- Pirkle, H.; Markland, F.S., Jr. Hemostasis and Animal Venoms; Dekker: New York, NY, USA, 1987. [Google Scholar]
- Clemetson, K.J.; Lu, Q.; Clemetson, J.M. Snake venom proteins affecting platelets and their applications to anti-thrombotic research. Curr. Pharm. Des. 2007, 13, 2887–2892. [Google Scholar]
- Clemetson, K.J.; Lu, Q.; Clemetson, J.M. Snake C-type lectin-like proteins and platelet receptors. Pathophysiol. Haemost. Thromb. 2005, 34, 150–155. [Google Scholar]
- Ruggeri, Z.M. Platelets in atherothrombosis. Nat. Med. 2002, 8, 1227–1234. [Google Scholar]
- Lusis, A.J. Atherosclerosis. Nature 2000, 407, 233–241. [Google Scholar]
- Ho, M.; Warrell, D.A.; Looareesuwan, S.; Phillips, R.E.; Chanthavanich, P.; Karbwang, J.; Supanaranond, W.; Viravan, C.; Hutton, R.A.; Vejcho, S. Clinical significance of venom antigen levels in patients envenomed by the malayan pit viper (Calloselasma rhodostoma). Am. J. Trop. Med. Hyg. 1986, 35, 579–587. [Google Scholar] [PubMed]
- Huang, T.F.; Liu, C.Z.; Yang, S.H. Aggretin, a novel platelet-aggregation inducer from snake (Calloselasma rhodostoma) venom, activates phospholipase c by acting as a glycoprotein ia/iia agonist. Biochem. J. 1995, 309, 1021–1027. [Google Scholar] [PubMed]
- Shin, Y.; Morita, T. Rhodocytin, a functional novel platelet agonist belonging to the heterodimeric c-type lectin family, induces platelet aggregation independently of glycoprotein ib. Biochem. Biophys. Res. Commun. 1998, 245, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.H.; Au, L.C.; Huang, T.F. Molecular cloning and sequence analysis of aggretin, a collagen-like platelet aggregation inducer. Biochem. Biophys. Res. Commun. 1999, 263, 723–727. [Google Scholar]
- Suzuki-Inoue, K.; Fuller, G.L.; Garcia, A.; Eble, J.A.; Pohlmann, S.; Inoue, O.; Gartner, T.K.; Hughan, S.C.; Pearce, A.C.; Laing, G.D.; et al. A novel Syk-dependent mechanism of platelet activation by the C-type lectin receptor CLEC-2. Blood 2006, 107, 542–549. [Google Scholar] [PubMed]
- Fuller, G.L.; Williams, J.A.; Tomlinson, M.G.; Eble, J.A.; Hanna, S.L.; Pohlmann, S.; Suzuki-Inoue, K.; Ozaki, Y.; Watson, S.P.; Pearce, A.C. The C-type lectin receptors CLEC-2 and Dectin-1, but not DC-SIGN, signal via a novel YXXL-dependent signaling cascade. J. Biol. Chem. 2007, 282, 12397–12409. [Google Scholar] [PubMed]
- O’Callaghan, C.A. Thrombomodulation via CLEC-2 targeting. Curr. Opin. Pharmacol. 2009, 9, 90–95. [Google Scholar]
- O’Callaghan, C.A.; Tormo, J.; Willcox, B.E.; Braud, V.M.; Jakobsen, B.K.; Stuart, D.I.; McMichael, A.J.; Bell, J.I.; Jones, E.Y. Structural features impose tight peptide binding specificity in the non-classical MHC molecule HLA-E. Mol. Cell 1998, 1, 531–541. [Google Scholar]
- Braud, V.; Allan, D.S.J.; O’Callaghan, C.A.; Ogg, G.S.; Lazetic, S.; Young, N.T.; Bell, J.I.; Phillips, J.H.; Lanier, L.L.; McMichael, A.J. HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 1998, 391, 795–799. [Google Scholar]
- O’Callaghan, C.A.; Cerwenka, A.; Willcox, B.E.; Lanier, L.L.; Bjorkman, P.J. Molecular competition for NKG2D: H60 and RAE1 compete unequally for NKG2D with dominance of H60. Immunity 2001, 15, 201–211. [Google Scholar]
- Christou, C.M.; Pearce, A.C.; Watson, A.A.; Mistry, A.R.; Pollitt, A.Y.; Fenton-May, A.E.; Johnson, L.A.; Jackson, D.G.; Watson, S.P.; O’Callaghan, C.A. Renal cells activate the platelet receptor CLEC-2 through podoplanin. Biochem. J. 2008, 411, 133–140. [Google Scholar]
- O’Callaghan, C.A.; Jones, E.Y. Structural and energetic aspects of multispecific immune recognition by NKG2D. Structure (Camb.) 2003, 11, 360–361. [Google Scholar] [CrossRef]
- Cueni, L.N.; Chen, L.; Zhang, H.; Marino, D.; Huggenberger, R.; Alitalo, A.; Bianchi, R.; Detmar, M. Podoplanin-fc reduces lymphatic vessel formation in vitro and in vivo and causes disseminated intravascular coagulation when transgenically expressed in the skin. Blood 2010, 116, 4376–4384. [Google Scholar] [CrossRef] [PubMed]
- Suzuki-Inoue, K.; Kato, Y.; Inoue, O.; Kaneko, M.K.; Mishima, K.; Yatomi, Y.; Narimatsu, H.; Ozaki, Y. Involvement of the snake toxin receptor CLEC-2 in podoplanin-mediated platelet activation by cancer cells. J.Biol.Chem. 2007, 282, 25993–26001. [Google Scholar]
- Martin-Villar, E.; Scholl, F.G.; Gamallo, C.; Yurrita, M.M.; Munoz-Guerra, M.; Cruces, J.; Quintanilla, M. Characterization of human pa2.26 antigen (t1alpha-2, podoplanin), a small membrane mucin induced in oral squamous cell carcinomas. Int. J. Cancer 2005, 113, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Uhrin, P.; Zaujec, J.; Breuss, J.M.; Olcaydu, D.; Chrenek, P.; Stockinger, H.; Fuertbauer, E.; Moser, M.; Haiko, P.; Fassler, R.; et al. Novel function for blood platelets and podoplanin in developmental separation of blood and lymphatic circulation. Blood 2010, 115, 3997–4005. [Google Scholar] [CrossRef] [PubMed]
- Schacht, V.; Ramirez, M.I.; Hong, Y.K.; Hirakawa, S.; Feng, D.; Harvey, N.; Williams, M.; Dvorak, A.M.; Dvorak, H.F.; Oliver, G.; et al. T1alpha/podoplanin deficiency disrupts normal lymphatic vasculature formation and causes lymphedema. EMBO J. 2003, 22, 3546–3556. [Google Scholar] [CrossRef]
- Mahtab, E.A.; Vicente-Steijn, R.; Hahurij, N.D.; Jongbloed, M.R.; Wisse, L.J.; DeRuiter, M.C.; Uhrin, P.; Zaujec, J.; Binder, B.R.; Schalij, M.J.; et al. Podoplanin deficient mice show a rhoa-related hypoplasia of the sinus venosus myocardium including the sinoatrial node. Dev. Dyn. 2009, 238, 183–193. [Google Scholar] [PubMed]
- Douglas, Y.L.; Mahtab, E.A.; Jongbloed, M.R.; Uhrin, P.; Zaujec, J.; Binder, B.R.; Schalij, M.J.; Poelmann, R.E.; Deruiter, M.C.; Gittenberger-de Groot, A.C. Pulmonary vein, dorsal atrial wall and atrial septum abnormalities in podoplanin knockout mice with disturbed posterior heart field contribution. Pediatr. Res. 2009, 65, 27–32. [Google Scholar]
- Hughes, C.E.; Navarro-Núñez, L.; Finney, B.A.; Mourão-Sá, D.; Pollitt, A.Y.; Watson, S.P. CLEC-2 is not required for platelet aggregation at arteriolar shear. J. Thromb. Haemost. 2010, 8, 2328–2332. [Google Scholar]
- Tang, T.; Li, L.; Tang, J.; Li, Y.; Lin, W.Y.; Martin, F.; Grant, D.; Solloway, M.; Parker, L.; Ye, W.; et al. A mouse knockout library for secreted and transmembrane proteins. Nat. Biotechnol. 2010, 28, 749–755. [Google Scholar] [PubMed]
- Bertozzi, C.C.; Schmaier, A.A.; Mericko, P.; Hess, P.R.; Zou, Z.; Chen, M.; Chen, C.Y.; Xu, B.; Lu, M.M.; Zhou, D.; et al. Platelets regulate lymphatic vascular development through CLEC-2-SLP-76 signaling. Blood 2010, 116, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Suzuki-Inoue, K.; Inoue, O.; Ding, G.; Nishimura, S.; Hokamura, K.; Eto, K.; Kashiwagi, H.; Tomiyama, Y.; Yatomi, Y.; Umemura, K.; et al. Essential in vivo roles of the C-type lectin receptor CLEC-2: Embryonic/neonatal lethality of CLEC-2-deficient mice by blood/lymphatic misconnections and impaired thrombus formation of CLEC-2-deficient platelets. J. Biol. Chem. 2010, 285, 24494–24507. [Google Scholar] [PubMed]
- Chippaux, J.P. Snake-bites: Appraisal of the global situation. Bull. WHO 1998, 76, 515–524. [Google Scholar]
- Watson, A.A.; Brown, J.; Harlos, K.; Eble, J.A.; Walter, T.S.; O’Callaghan, C.A. The crystal structure and mutational binding analysis of the extracellular domain of the platelet-activating receptor CLEC-2. J. Biol. Chem. 2007, 282, 3165–3172. [Google Scholar]
- Watson, A.A.; O’Callaghan, C.A. Crystallization and X-ray diffraction analysis of human CLEC-2. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2005, 61, 1094–1096. [Google Scholar]
- Watson, A.A.; Christou, C.M.; James, J.R.; Fenton-May, A.E.; Moncayo, G.E.; Mistry, A.R.; Davis, S.J.; Gilbert, R.J.; Chakera, A.; O’Callaghan, C.A. The platelet receptor CLEC-2 is active as a dimer. Biochemistry 2009, 48, 10988–10996. [Google Scholar]
- Watson, A.A.; Eble, J.A.; O’Callaghan, C.A. Crystal structure of rhodocytin, a ligand for the platelet-activating receptor CLEC-2. Protein Sci. 2008, 17, 1611–1616. [Google Scholar]
- Hughes, C.E.; Pollitt, A.Y.; Mori, J.; Eble, J.A.; Tomlinson, M.G.; Hartwig, J.H.; O’Callaghan, C.A.; Futterer, K.; Watson, S.P. CLEC-2 activates Syk through dimerization. Blood 2010, 115, 2947–2955. [Google Scholar] [Green Version]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol.Biol. 2007, 372, 774–797. [Google Scholar]
- Barrett, C.P.; Hall, B.A.; Noble, M.E. Dynamite: A simple way to gain insight into protein motions. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2280–2287. [Google Scholar]
- De Groot, B.L.; van Aalten, D.M.; Scheek, R.M.; Amadei, A.; Vriend, G.; Berendsen, H.J. Prediction of protein conformational freedom from distance constraints. Proteins 1997, 29, 240–251. [Google Scholar]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Watson, A.A.; O’Callaghan, C.A. Molecular Analysis of the Interaction of the Snake Venom Rhodocytin with the Platelet Receptor CLEC-2. Toxins 2011, 3, 991-1003. https://doi.org/10.3390/toxins3080991
Watson AA, O’Callaghan CA. Molecular Analysis of the Interaction of the Snake Venom Rhodocytin with the Platelet Receptor CLEC-2. Toxins. 2011; 3(8):991-1003. https://doi.org/10.3390/toxins3080991
Chicago/Turabian StyleWatson, Aleksandra A., and Christopher A. O’Callaghan. 2011. "Molecular Analysis of the Interaction of the Snake Venom Rhodocytin with the Platelet Receptor CLEC-2" Toxins 3, no. 8: 991-1003. https://doi.org/10.3390/toxins3080991
APA StyleWatson, A. A., & O’Callaghan, C. A. (2011). Molecular Analysis of the Interaction of the Snake Venom Rhodocytin with the Platelet Receptor CLEC-2. Toxins, 3(8), 991-1003. https://doi.org/10.3390/toxins3080991