Host-Defense Activities of Cyclotides

Abstract
:1. Introduction


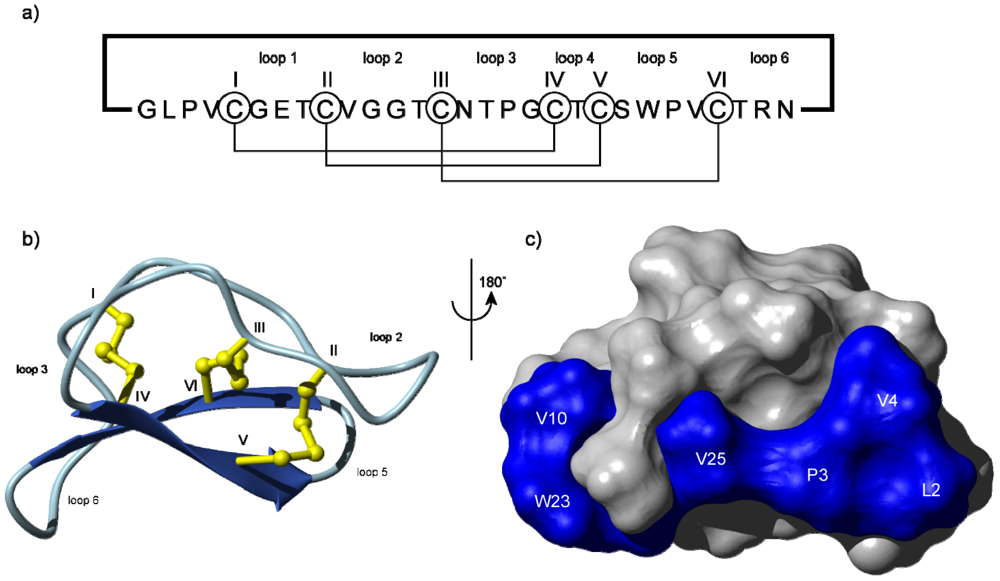{kind=link}
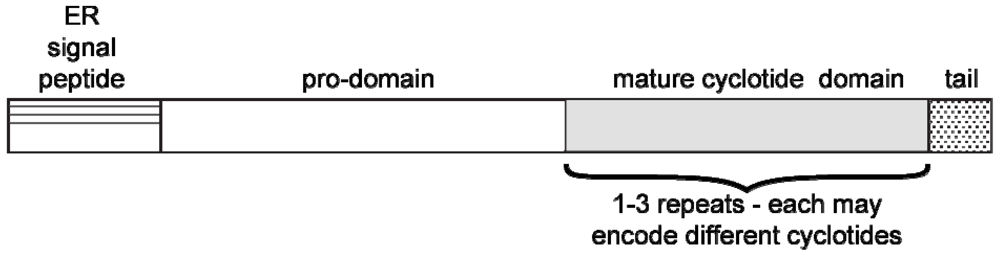{kind=link}
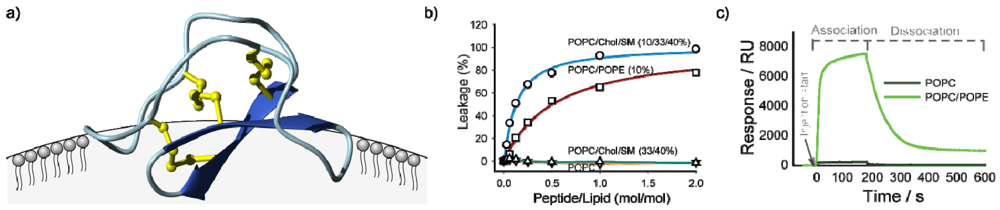{kind=link}
{kind=link}
| Cyclotide a | AA b | Ch c | Sequence d | PDB e | Ref. |
|---|---|---|---|---|---|
| Möbius subfamily | |||||
| kalata B1 | 30 | +2 | GLPVCGETCVGGTCNTPGCTCSWPVCTRN | 1NB1 | [13] |
| kalata B2 | 29 | −1 | GLPVCGETCFGGTCNTPGCSCTWPICTRD | 1PT4 | [22,23] |
| kalata B7 | 29 | +1 | GLPVCGETCTLGTCYTQGCTCSWPICKRN | 2JWM | [24] |
| kalata B12 | 28 | −2 | GSLCGDTCFVLGCNDSSCSCNYPICVKD | 2KVX | [25] |
| cycloviolacin O14 | 31 | +3 | GSIPACGESCFKGKCYTPGCSCSKYPLCAKN | 2GJ0 | [26] |
| varv F | 29 | 0 | GVPICGETCTLGTCYTAGCSCSWPVCTRN | 3E4H | [27] |
| Bracelet subfamily | |||||
| circulin A | 30 | +2 | GIPCGESCVWIPCISAALGCSCKNKVCYRN | 1BH4 | [28,29] |
| circulin B | 31 | +2 | GVIPCGESCVFIPCISTLLGCSCKNKVCYRN | 2ERI | [29] |
| cycloviolacin O1 | 30 | 0 | GIPCAESCVYIPCTVTALLGCSCSNRVCYN | 1NBJ | [26] |
| cycloviolacin O2 | 30 | +2 | GIPCGESCVWIPCISSAIGCSCKSKVCYRN | 2KNM | [26] |
| kalata B5 | 30 | −1 | GTPCGESCVYIPCISGVIGCSCTDKVCYLN | 2KUX | [30,31] |
| kalata B8 | 31 | +1 | GSVLNCGETCLLGTCYTTGCTCNKYRVCTKD | 2B38 | [32] |
| tricyclon A | 33 | −1 | GGTIFDCGESCFLGTCYTKGCSCGEWKLCYGTN | 1YP8 | [33] |
| palicourein | 37 | −1 | GDPTFCGETCRVIPVCTYSAALGCTCDDRSDGLCKRN | 1R1F | [34,35] |
| vhl-1 | 31 | 0 | SISCGESCAMISFCFTEVIGCSCKNKVCYLN | 1ZA8 | [36] |
| vhl-2 | 30 | −1 | GLPVCGETCFTGTCYTNGCTCDPWPVCTRN | 2KUK | [36,37] |
| vhr1 | 30 | 0 | GIPCAESCVWIPCTVTALLGCSCSNKVCYN | 1VB8 | [3] |
| Trypsin inhibitor subfamily | |||||
| MCoTI-II | 34 | +3 | GGVCPKILKKCRRDSDCPGACICRGNGYCGSGSD | 1IB9 | [38,39] |

2. Pesticidal and/or Toxic Activities
| Cyclotide | Activity | Ref. |
|---|---|---|
| Möbius subfamily | ||
| kalata B1 | Insecticidal, molluscicidal, hemolytic, nematocidal, antibacterial, anti-HIV | [47,53,54,55,56] |
| kalata B2 | Insecticidal, molluscicidal, nematocidal, antibacterial | [22,56,57] |
| kalata B5 | Molluscicidal | [56] |
| cycloviolacin O14 | Nematocidal, anti-HIV, hemolytic | [26] |
| varv A & F | Cytotoxic | [27,58] |
| Bracelet subfamily | ||
| circulin A & B | Hemolytic, anti-bacterial, anti-HIV | [29] |
| cycloviolacin O1 | Nematocidal, molluscicidal | [26,56,59,60] |
| cycloviolacin O2 | Nematocidal, cytotoxicity, hemolytic, anti-barnacle, antibacterial | [57,61] |
| Cycloviolacin Y1 | Hemolytic, anti-HIV | [62] |
| vhl-1 | Nematocidal, anti-HIV | [36] |
| Trypsin inhibitor subfamily | ||
| MCoTI-II | Protease inhibition | [63,64,65] |
2.1. Hemolytic and Cytotoxic Activities
2.2. Antimicrobial Activity
2.3. Insecticidal Activity
2.4. Anti-Fouling Activities
2.5. Molluscicidal Activities
2.6. Nematocidal Activities
3. Mechanisms of Action

4. Methods of Production of Cyclotides

5. Safety and Breakdown
6. Conclusions
Acknowledgments
Conflict of Interest
References
- Craik, D.J.; Daly, N.L.; Bond, T.; Waine, C. Plant cyclotides: A unique family of cyclic and knotted proteins that defines the cyclic cystine knot structural motif. J. Mol. Biol. 1999, 294, 1327–1336. [Google Scholar]
- Gruber, C.W.; Elliott, A.G.; Ireland, D.C.; Delprete, P.G.; Dessein, S.; Goransson, U.; Trabi, M.; Wang, C.K.; Kinghorn, A.B.; Robbrecht, E.; Craik, D.J. Distribution and evolution of circular miniproteins in flowering plants. Plant Cell 2008, 20, 2471–2483. [Google Scholar]
- Trabi, M.; Craik, D.J. Tissue-specific expression of head-to-tail cyclized miniproteins in Violaceae and structure determination of the root cyclotide Viola hederacea root cyclotide1. Plant Cell 2004, 16, 2204–2216. [Google Scholar]
- Craik, D.J.; Daly, N.L.; Mulvenna, J.; Plan, M.R.; Trabi, M. Discovery, structure and biological activities of the cyclotides. Curr. Protein Pept. Sci. 2004, 5, 297–315. [Google Scholar]
- Göransson, U.; Svangard, E.; Claeson, P.; Bohlin, L. Novel strategies for isolation and characterization of cyclotides: The discovery of bioactive macrocyclic plant polypeptides in the Violaceae. Curr. Protein Pept. Sci. 2004, 5, 317–329. [Google Scholar]
- Gran, L.; Sandberg, F.; Sletten, K. Oldenlandia affinis (R&S) DC. A plant containing uteroactive peptides used in African traditional medicine. J. Ethnopharmacol. 2000, 70, 197–203. [Google Scholar] [CrossRef]
- Daly, N.L.; Rosengren, K.J.; Craik, D.J. Discovery, structure and biological activities of cyclotides. Adv.Drug Deliv. Rev. 2009, 61, 918–930. [Google Scholar]
- Pelegrini, P.B.; Quirino, B.F.; Franco, O.L. Plant cyclotides: An unusual class of defense compounds. Peptides 2007, 28, 1475–1481. [Google Scholar]
- Craik, D.J.; Cĕmažar, M.; Daly, N.L. The cyclotides and related macrocyclic peptides as scaffolds in drug design. Curr. Opin. Drug Discov. Dev. 2006, 9, 251–260. [Google Scholar]
- Craik, D.J.; Clark, R.J.; Daly, N.L. Potential therapeutic applications of the cyclotides and related cystine knot mini-proteins. Expert Opin. Investig. Drugs 2007, 16, 595–604. [Google Scholar]
- Sancheti, H.; Camarero, J.A. “Splicing up” drug discovery. Cell-based expression and screening of genetically-encoded libraries of backbone-cyclized polypeptides. Adv. Drug Deliv. Rev. 2009, 61, 908–917. [Google Scholar] [CrossRef]
- Gruber, C.W.; Cemazar, M.; Anderson, M.A.; Craik, D.J. Insecticidal plant cyclotides and related cystine knot toxins. Toxicon 2007, 49, 561–575. [Google Scholar]
- Saether, O.; Craik, D.J.; Campbell, I.D.; Sletten, K.; Juul, J.; Norman, D.G. Elucidation of the primary and three-dimensional structure of the uterotonic polypeptide kalata B1. Biochemistry 1995, 34, 4147–4158. [Google Scholar]
- Colgrave, M.L.; Craik, D.J. Thermal, chemical, and enzymatic stability of the cyclotide kalata B1: The importance of the cyclic cystine knot. Biochemistry 2004, 43, 5965–5975. [Google Scholar] [CrossRef]
- Craik, D.J.; Daly, N.L.; Waine, C. The cystine knot motif in toxins and implications for drug design. Toxicon 2001, 39, 43–60. [Google Scholar]
- Göransson, U.; Craik, D.J. Disulfide mapping of the cyclotide kalata B1. Chemical proof of the cyclic cystine knot motif. J. Biol. Chem. 2003, 278, 48188–48196. [Google Scholar]
- Rosengren, K.J.; Daly, N.L.; Plan, M.R.; Waine, C.; Craik, D.J. Twists, knots, and rings in protein. Structural definition of the cyclotide framework. J. Biol. Chem. 2003, 278, 8606–8616. [Google Scholar]
- Craik, D.J.; Cemazar, M.; Wang, C.K.; Daly, N.L. The cyclotide family of circular miniproteins: Nature’s combinatorial peptide template. Biopolymers 2006, 84, 250–266. [Google Scholar]
- Poth, A.G.; Colgrave, M.L.; Philip, R.; Kerenga, B.; Daly, N.L.; Anderson, M.A.; Craik, D.J. Discovery of cyclotides in the fabaceae plant family provides new insights into the cyclization, evolution, and distribution of circular proteins. ACS Chem. Biol. 2011, 6, 345–355. [Google Scholar] [CrossRef]
- Nguyen, G.K.; Zhang, S.; Nguyen, N.T.; Nguyen, P.Q.; Chiu, M.S.; Hardjojo, A.; Tam, J.P. Discovery and characterization of novel cyclotides originated from chimeric precursors consisting of albumin-1 chain a and cyclotide domains in the fabaceae family. J. Biol. Chem. 2011, 286, 24275–24287. [Google Scholar]
- Chiche, L.; Heitz, A.; Gelly, J.C.; Gracy, J.; Chau, P.T.; Ha, P.T.; Hernandez, J.F.; Le-Nguyen, D. Squash inhibitors: From structural motifs to macrocyclic knottins. Curr. Protein Pept. Sci. 2004, 5, 341–349. [Google Scholar]
- Jennings, C.V.; Rosengren, K.J.; Daly, N.L.; Plan, M.; Stevens, J.; Scanlon, M.J.; Waine, C.; Norman, D.G.; Anderson, M.A.; Craik, D.J. Isolation, solution structure, and insecticidal activity of kalata B2, a circular protein with a twist: Do Mobius strips exist in nature? Biochemistry 2005, 44, 851–860. [Google Scholar]
- Nair, S.S.; Romanuka, J.; Billeter, M.; Skjeldal, L.; Emmett, M.R.; Nilsson, C.L.; Marshall, A.G. Structural characterization of an unusually stable cyclic peptide, kalata B2 from Oldenlandia affinis. Biochim. Biophys.Acta 2006, 1764, 1568–1576. [Google Scholar]
- Craik, D.J. Plant cyclotides: Circular, knotted peptide toxins. Toxicon 2001, 39, 1809–1813. [Google Scholar]
- Wang, C.K.L.; Clark, R.J.; Cemazar, M.; Craik, D.J. Role of conserved Glu in cyclotide stability and activity: A structural and functional study of kalata B12, a naturally occurring Glu to Asp mutant. Biochemistry 2011, 50, 4077–4086. [Google Scholar]
- Ireland, D.C.; Colgrave, M.L.; Craik, D.J. A novel suite of cyclotides from Viola odorata: Sequence variation and the implications for structure, function and stability. Biochem. J. 2006, 400, 1–12. [Google Scholar]
- Wang, C.K.; Hu, S.H.; Martin, J.L.; Sjogren, T.; Hajdu, J.; Bohlin, L.; Claeson, P.; Goransson, U.; Rosengren, K.J.; Tang, J.; Tan, N.H.; Craik, D.J. Combined X-ray and NMR analysis of the stability of the cyclotide cystine knot fold that underpins its insecticidal activity and potential use as drug scaffold. J. Biol. Chem. 2009, 284, 10672–10683. [Google Scholar]
- Daly, N.L.; Koltay, A.; Gustafson, K.R.; Boyd, M.R.; Casas-Finet, J.R.; Craik, D.J. Solution structure by NMR of circulin A: A macrocyclic knotted peptide having anti-HIV activity. J. Mol. Biol. 1999, 285, 333–345. [Google Scholar]
- Gustafson, K.R.; Sowder, R.C.I.; Henderson, L.E.; Parsons, I.C.; Kashman, Y.; Cardellina, J.H.I.; McMahon, J.B.; Buckheit, R.W.J.; Pannell, L.K.; Boyd, M.R. Circulins A and B: Novel HIV-inhibitory macrocyclic peptides from the tropical tree Chassalia parvifolia. J. Am. Chem. Soc. 1994, 116, 9337–9338. [Google Scholar]
- Plan, M.R.; Rosengren, K.J.; Sando, L.; Daly, N.L.; Craik, D.J. Structural and biochemical characteristics of the cyclotide kalata B5 from Oldenlandia affinis. Biopolym. Pept. Sci. 2010, 94, 647–658. [Google Scholar]
- Plan, M.R.R.; Göransson, U.; Clark, R.J.; Daly, N.L.; Colgrave, M.L.; Craik, D.J. The cyclotide fingerprint in Oldenlandia affinis: Elucidation of chemically modified, linear and novel macrocyclic peptides. ChemBioChem 2007, 8, 1001–1011. [Google Scholar]
- Daly, N.L.; Clark, R.J.; Plan, M.R.; Craik, D.J. Kalata B8, a novel antiviral circular protein, exhibits conformational flexibility in the cystine knot motif. Biochem. J. 2006, 393, 619–626. [Google Scholar] [CrossRef]
- Mulvenna, J.P.; Sando, L.; Craik, D.J. Processing of a 22 kDa precursor protein to produce the circular protein tricyclon A. Structure 2005, 13, 691–701. [Google Scholar]
- Barry, D.G.; Daly, N.L.; Bokesch, H.R.; Gustafson, K.R.; Craik, D.J. Solution structure of the cyclotide palicourein: Implications for the development of a pharmaceutical framework. Structure 2004, 12, 85–94. [Google Scholar]
- Bokesch, H.R.; Pannell, L.K.; Cochran, P.K.; Sowder, R.C., II; McKee, T.C.; Boyd, M.R. A novel anti-HIV macrocyclic peptide from Palicourea condensata. J. Nat. Prod. 2001, 64, 249–250. [Google Scholar] [CrossRef]
- Chen, B.; Colgrave, M.L.; Daly, N.L.; Rosengren, K.J.; Gustafson, K.R.; Craik, D.J. Isolation and characterization of novel cyclotides from Viola hederaceae: Solution structure and anti-HIV activity of vhl-1, a leaf-specific expressed cyclotide. J. Biol. Chem. 2005, 280, 22395–22405. [Google Scholar]
- Chen, B.; Colgrave, M.L.; Wang, C.; Craik, D.J. Cycloviolacin H4, a hydrophobic cyclotide from Viola hederaceae. J. Nat. Prod. 2006, 69, 23–28. [Google Scholar]
- Felizmenio-Quimio, M.E.; Daly, N.L.; Craik, D.J. Circular proteins in plants: Solution structure of a novel macrocyclic trypsin inhibitor from Momordica cochinchinensis. J. Biol. Chem. 2001, 276, 22875–22882. [Google Scholar]
- Heitz, A.; Hernandez, J.F.; Gagnon, J.; Hong, T.T.; Pham, T.T.; Nguyen, T.M.; Le-Nguyen, D.; Chiche, L. Solution structure of the squash trypsin inhibitor MCoTI-II. A new family for cyclic knottins. Biochemistry 2001, 40, 7973–7983. [Google Scholar]
- Wang, C.K.; Kaas, Q.; Chiche, L.; Craik, D.J. CyBase: A database of cyclic protein sequences and structures, with applications in protein discovery and engineering. Nucleic Acids Res. 2008, 36, D206–D210. [Google Scholar]
- Gran, L. An oxytocic principle found in Oldenlandia affinis DC. Medd. Nor. Farm. Selsk. 1970, 12, 173–180. [Google Scholar]
- Schöpke, T.; Hasan Agha, M.I.; Kraft, R.; Otto, A.; Hiller, K. Hämolytisch aktive komponenten aus Viola tricolor L. und Viola arvensis Murray. Sci. Pharm. 1993, 61, 145–153. [Google Scholar]
- Gustafson, K.R.; McKee, T.C.; Bokesch, H.R. Anti-HIV cyclotides. Curr. Protein Pept. Sci. 2004, 5, 331–340. [Google Scholar]
- Gustafson, K.R.; Walton, L.K.; Sowder, R.C.I.; Johnson, D.G.; Pannell, L.K.; Cardellina, J.H.I.; Boyd, M.R. New circulin macrocyclic polypeptides from Chassalia parvifolia. J. Nat. Prod. 2000, 63, 176–178. [Google Scholar]
- Witherup, K.M.; Bogusky, M.J.; Anderson, P.S.; Ramjit, H.; Ransom, R.W.; Wood, T.; Sardana, M. Cyclopsychotride A, A biologically active, 31-residue cyclic peptide isolated from Psychotria longipes. J. Nat. Prod. 1994, 57, 1619–1625. [Google Scholar] [CrossRef]
- Tam, J.P.; Lu, Y.A.; Yang, J.L.; Chiu, K.W. An unusual structural motif of antimicrobial peptides containing end-to-end macrocycle and cystine-knot disulfides. Proc. Natl. Acad. Sci. USA 1999, 96, 8913–8918. [Google Scholar]
- Jennings, C.; West, J.; Waine, C.; Craik, D.; Anderson, M. Biosynthesis and insecticidal properties of plant cyclotides: The cyclic knotted proteins from Oldenlandia affinis. Proc. Natl. Acad. Sci. USA 2001, 98, 10614–10619. [Google Scholar]
- Craik, D.J.; Anderson, M.A.; Barry, D.G.; Clark, R.J.; Daly, N.L.; Jennings, C.V.; Mulvenna, J. Discovery and structures of the cyclotides: Novel macrocyclic peptides from plants. Lett. Pep. Sci. 2002, 8, 119–128. [Google Scholar]
- Dutton, J.L.; Renda, R.F.; Waine, C.; Clark, R.J.; Daly, N.L.; Jennings, C.V.; Anderson, M.A.; Craik, D.J. Conserved structural and sequence elements implicated in the processing of gene-encoded circular proteins. J. Biol. Chem. 2004, 279, 46858–46867. [Google Scholar]
- Saska, I.; Gillon, A.D.; Hatsugai, N.; Dietzgen, R.G.; Hara-Nishimura, I.; Anderson, M.A.; Craik, D.J. An asparaginyl endopeptidase mediates in vivo protein backbone cyclisation. J. Biol. Chem. 2007, 282, 29721–29728. [Google Scholar]
- Gillon, A.D.; Saska, I.; Jennings, C.V.; Guarino, R.F.; Craik, D.J.; Anderson, M.A. Biosynthesis of circular proteins in plants. Plant J. 2008, 53, 505–515. [Google Scholar]
- Poth, A.G.; Colgrave, M.L.; Lyons, R.E.; Daly, N.L.; Craik, D.J. From the cover: Discovery of an unusual biosynthetic origin for circular proteins in legumes. Proc. Natl. Acad. Sci. USA 2011, 108, 10127–10132. [Google Scholar]
- Daly, N.L.; Gustafson, K.R.; Craik, D.J. The role of the cyclic peptide backbone in the anti-HIV activity of the cyclotide kalata B1. FEBS Lett. 2004, 574, 69–72. [Google Scholar]
- Gran, L. Oxytocic principles of Oldenlandia affinis. Lloydia 1973, 36, 174–178. [Google Scholar]
- Colgrave, M.L.; Kotze, A.C.; Huang, Y.H.; O’Grady, J.; Simonsen, S.M.; Craik, D.J. Cyclotides: Natural, circular plant peptides that possess significant activity against gastrointestinal nematode parasites of sheep. Biochemistry 2008, 47, 5581–5589. [Google Scholar]
- Plan, M.R.; Saska, I.; Cagauan, A.G.; Craik, D.J. Backbone cyclised peptides from plants show molluscicidal activity against the rice pest Pomacea canaliculata (golden apple snail). J. Agric. Food Chem. 2008, 56, 5237–5241. [Google Scholar]
- Ovesen, R.G.; Brandt, K.K.; Göransson, U.; Nilesen, J.; Hansen, H.C.B.; Cedergreen, N. Biomedicine in the environment: Cyclotides constitute potent natural toxins in plants and soil bacteria. Environ. Toxinol. Chem. 2011, 30, 119–1196. [Google Scholar]
- Lindholm, P.; Göransson, U.; Johansson, S.; Claeson, P.; Gullbo, J.; Larsson, R.; Bohlin, L.; Backlund, A. Cyclotides: A novel type of cytotoxic agents. Mol. Cancer Ther. 2002, 1, 365–369. [Google Scholar]
- Colgrave, M.L.; Kotze, A.C.; Ireland, D.C.; Wang, C.K.; Craik, D.J. The anthelmintic activity of the cyclotides: Natural variants with enhanced activity. ChemBioChem 2008, 9, 1939–1945. [Google Scholar]
- Colgrave, M.L.; Kotze, A.C.; Kopp, S.; McCarthy, J.S.; Coleman, G.T.; Craik, D.J. Anthelmintic activity of cyclotides: In vitro studies with canine and human hookworms. Acta Trop. 2009, 109, 163–166. [Google Scholar]
- Göransson, U.; Sjogren, M.; Svangard, E.; Claeson, P.; Bohlin, L. Reversible antifouling effect of the cyclotide cycloviolacin O2 against barnacles. J. Nat. Prod. 2004, 67, 1287–1290. [Google Scholar]
- Wang, C.K.; Colgrave, M.L.; Gustafson, K.R.; Ireland, D.C.; Göransson, U.; Craik, D.J. Anti-HIV cyclotides from the Chinese medicinal herb Viola yedoensis. J. Nat. Prod. 2008, 71, 47–52. [Google Scholar]
- Hernandez, J.F.; Gagnon, J.; Chiche, L.; Nguyen, T.M.; Andrieu, J.P.; Heitz, A.; Trinh Hong, T.; Pham, T.T.; le Nguyen, D. Squash trypsin inhibitors from Momordica cochinchinensis exhibit an atypical macrocyclic structure. Biochemistry 2000, 39, 5722–5730. [Google Scholar]
- Thongyoo, P.; Bonomelli, C.; Leatherbarrow, R.J.; Tate, E.W. Potent inhibitors of beta-tryptase and human leukocyte elastase based on the MCoTI-II scaffold. J. Med. Chem. 2009, 52, 6197–6200. [Google Scholar]
- Thongyoo, P.; Roque-Rosell, N.; Leatherbarrow, R.J.; Tate, E.W. Chemical and biomimetic total syntheses of natural and engineered MCoTI cyclotides. Org. Biomol. Chem. 2008, 6, 1462–1470. [Google Scholar]
- Daly, N.L.; Craik, D.J. Acyclic permutants of naturally occurring cyclic proteins. Characterization of cystine knot and beta -sheet formation in the macrocyclic polypeptide kalata B1. J. Biol. Chem. 2000, 275, 19068–19075. [Google Scholar]
- Barry, D.G.; Daly, N.L.; Clark, R.J.; Sando, L.; Craik, D.J. Linearization of a naturally occurring circular protein maintains structure but eliminates hemolytic activity. Biochemistry 2003, 42, 6688–6695. [Google Scholar]
- Simonsen, S.M.; Sando, L.; Rosengren, K.J.; Wang, C.K.; Colgrave, M.L.; Daly, N.L.; Craik, D.J. Alanine scanning mutagenesis of the prototypic cyclotide reveals a cluster of residues essential for bioactivity. J. Biol. Chem. 2008, 283, 9805–9813. [Google Scholar]
- Svangard, E.; Burman, R.; Gunasekera, S.; Lovborg, H.; Gullbo, J.; Goransson, U. Mechanism of action of cytotoxic cyclotides: Cycloviolacin O2 disrupts lipid membranes. J. Nat. Prod. 2007, 70, 643–647. [Google Scholar]
- Svangard, E.; Göransson, U.; Hocaoglu, Z.; Gullbo, J.; Larsson, R.; Claeson, P.; Bohlin, L. Cytotoxic cyclotides from Viola tricolor. J. Nat. Prod. 2004, 67, 144–147. [Google Scholar]
- Svangard, E.; Göransson, U.; Smith, D.; Verma, C.; Backlund, A.; Bohlin, L.; Claeson, P. Primary and 3-D modelled structures of two cyclotides from Viola odorata. Phytochemistry 2003, 64, 135–142. [Google Scholar]
- Burman, R.; Svedlund, E.; Felth, J.; Hassan, S.; Herrmann, A.; Clark, R.J.; Craik, D.J.; Bohlin, L.; Claeson, P.; Goransson, U.; Gullbo, J. Evaluation of toxicity and anti-tumour activity of cycloviolacin O2 in mice. Biopolym. Pept. Sci. 2010, 94, 626–634. [Google Scholar]
- Gran, L.; Sletten, K.; Skjeldal, L. Cyclic peptides from Oldenlandia affinis DC. Molecular and biological properties. Chem. Biodivers. 2008, 5, 2014–2022. [Google Scholar] [CrossRef]
- Pranting, M.; Loov, C.; Burman, R.; Goransson, U.; Andersson, D.I. The cyclotide cycloviolacin O2 from Viola odorata has potent bactericidal activity against Gram-negative bacteria. J. Antimicrob. Chemoth. 2010, 65, 1964–1971. [Google Scholar]
- Barbeta, B.L.; Marshall, A.T.; Gillon, A.D.; Craik, D.J.; Anderson, M.A. Plant cyclotides disrupt epithelial cells in the midgut of lepidopteran larvae. Proc. Natl. Acad. Sci. USA 2008, 105, 1221–1225. [Google Scholar]
- Craik, D.J. Circling the enemy: Cyclic proteins in plant defence. Trends Plant Sci. 2009, 14, 328–335. [Google Scholar]
- Huang, Y.H.; Colgrave, M.L.; Clark, R.J.; Kotze, A.C.; Craik, D.J. Lysine-scanning mutagenesis reveals an amendable face of the cyclotide kalata B1 for the optimization of nematocidal activity. J. Biol. Chem. 2010, 285, 10797–10805. [Google Scholar]
- Shenkarev, Z.O.; Nadezhdin, K.D.; Sobol, V.A.; Sobol, A.G.; Skjeldal, L.; Arseniev, A.S. Conformation and mode of membrane interaction in cyclotides. Spatial structure of kalata B1 bound to a dodecylphosphocholine micelle. FEBS J. 2006, 273, 2658–2672. [Google Scholar] [CrossRef]
- Shenkarev, Z.O.; Nadezhdin, K.D.; Lyukmanova, E.N.; Sobol, V.A.; Skjeldal, L.; Arseniev, A.S. Divalent cation coordination and mode of membrane interaction in cyclotides: NMR spatial structure of ternary complex Kalata B7/Mn2+/DPC micelle. J. Inorg. Biochem. 2008, 102, 1246–1256. [Google Scholar]
- Wang, C.K.; Colgrave, M.L.; Ireland, D.C.; Kaas, Q.; Craik, D.J. Despite a conserved cystine knot motif, different cyclotides have different membrane binding modes. Biophys. J. 2009, 97, 1471–1481. [Google Scholar]
- Huang, Y.H.; Colgrave, M.L.; Daly, N.L.; Keleshian, A.; Martinac, B.; Craik, D.J. The biological activity of the prototypic cyclotide Kalata B1 is modulated by the formation of multimeric pores. J. Biol. Chem. 2009, 284, 20699–20707. [Google Scholar]
- Kamimori, H.; Hall, K.; Craik, D.J.; Aguilar, M.I. Studies on the membrane interactions of the cyclotides kalata B1 and kalata B6 on model membrane systems by surface plasmon resonance. Anal. Biochem. 2005, 337, 149–153. [Google Scholar]
- Henriques, S.T.; Huang, Y.-H.; Rosengren, K.J.; Franquelim, H.G.; Carvalho, F.A.; Johnson, A.; Sonza, S.; Tachedjian, G.; Castanha, M.A.R.B.; Daly, N.L.; Craik, D.J. Decoding the membrane activity of the cyclotide kalata B1: The importance of phosphatidylethanolamine phospholipids and lipid organization hemolytic and anti-HIV activities. J. Biol. Chem. 201, 286, 24231–24241. [Google Scholar]
- Herrmann, A.; Svangard, E.; Claeson, P.; Gullbo, J.; Bohlin, L.; Goransson, U. Key role of glutamic acid for the cytotoxic activity of the cyclotide cycloviolacin O2. Cell. Mol. Life Sci. 2006, 63, 235–245. [Google Scholar]
- Tam, J.P.; Lu, Y.-A. Synthesis of large cyclic cystine-knot peptide by orthogonal coupling strategy using unprotected peptide precursors. Tetrahedron Lett. 1997, 38, 5599–5602. [Google Scholar]
- Tam, J.P.; Lu, Y.-A. A biomimetic strategy in the synthesis and fragmentation of cyclic protein. Protein Sci. 1998, 7, 1583–1592. [Google Scholar]
- Clark, R.J.; Daly, N.L.; Craik, D.J. Structural plasticity of the cyclic-cystine-knot framework: Implications for biological activity and drug design. Biochem. J. 2006, 394, 85–93. [Google Scholar]
- Daly, N.L.; Love, S.; Alewood, P.F.; Craik, D.J. Chemical synthesis and folding pathways of large cyclic polypeptides: Studies of the cystine knot polypeptide kalata B1. Biochemistry 1999, 38, 10606–10614. [Google Scholar]
- Thongyoo, P.; Jaulent, A.M.; Tate, E.W.; Leatherbarrow, R.J. Immobilized protease-assisted synthesis of engineered cysteine-knot microproteins. ChemBioChem 2007, 8, 1107–1109. [Google Scholar]
- Thongyoo, P.; Tate, E.W.; Leatherbarrow, R.J. Total synthesis of the macrocyclic cysteine knot microprotein MCoTI-II. Chem. Commun. (Camb) 2006, 2848–2850. [Google Scholar]
- Kimura, R.H.; Tran, A.-T.; Camarero, J.A. Biosynthesis of the cyclotide kalata B1 by using protein splicing. Angew. Chem. Int. Ed. 2006, 118, 987–990. [Google Scholar]
- Austin, J.; Wang, W.; Puttamadappa, S.; Shekhtman, A.; Camarero, J.A. Biosynthesis and biological screening of a genetically encoded library based on the cyclotide MCoTI-I. ChemBioChem 2009, 10, 2663–2670. [Google Scholar]
- Camarero, J.A.; Kimura, R.H.; Woo, Y.-H.; Shekhtman, A.; Cantor, J. Biosynthesis of a fully functional cyclotide inside living bacterial cells. ChemBioChem 2007, 8, 1363–1366. [Google Scholar]
- Dawson, P.E.; Muir, T.W.; Clark-Lewis, I.; Kent, S.B. Synthesis of proteins by native chemical ligation. Science 1994, 266, 776–779. [Google Scholar]
- Kent, S.B. Total chemical synthesis of proteins. Chem. Soc. Rev. 2009, 38, 338–351. [Google Scholar]
- Gunasekera, S.; Daly, N.L.; Anderson, M.A.; Craik, D.J. Chemical synthesis and biosynthesis of the cyclotide family of circular proteins. IUBMB Life 2006, 58, 515–524. [Google Scholar]
- Leta Aboye, T.; Clark, R.J.; Craik, D.J.; Goransson, U. Ultra-stable peptide scaffolds for protein engineering-synthesis and folding of the circular cystine knotted cyclotide cycloviolacin O2. ChemBioChem 2008, 9, 103–113. [Google Scholar]
- Gunasekera, S.; Daly, N.L.; Clark, R.J.; Craik, D.J. Dissecting the oxidative folding of circular cystine knot miniproteins. Antioxid. Redox Signal. 2009, 11, 971–980. [Google Scholar]
- Marx, U.C.; Korsinczky, M.L.; Schirra, H.J.; Jones, A.; Condie, B.; Otvos, L., Jr.; Craik, D.J. Enzymatic cyclization of a potent Bowman-Birk protease inhibitor, sunflower trypsin inhibitor-1, and solution structure of an acyclic precursor peptide. J. Biol. Chem. 2003, 278, 21782–21789. [Google Scholar]
- Austin, J.; Kimura, R.H.; Woo, Y.H.; Camarero, J.A. In vivo biosynthesis of an Ala-scan library based on the cyclic peptide SFTI-1. Amino Acids 2010, 38, 1313–1322. [Google Scholar]
- Jagadish, K.; Camarero, J.A. Cyclotides, a promising molecular scaffold for peptide-based therapeutics. Biopolymers 2010, 94, 611–616. [Google Scholar]
- Dörnenburg, H. Plant cell culture technology-harnessing a biological approach for competitive cyclotides production. Biotechnol. Lett. 2008, 30, 1311–1321. [Google Scholar]
- Dörnenburg, H. Progress in kalata peptide production via plant cell bioprocessing. Biotechnol. J. 2009, 4, 632–645. [Google Scholar]
- Dörnenburg, H.; Frickinger, P.; Seydel, P. Plant cell-based processes for cyclotides production. J. Biotechnol. 2008, 135, 123–126. [Google Scholar]
- Seydel, P.; Dörnenburg, H. Establishment of in vitro plants, cell and tissue cultures from Oldenlandia affinis for the production of cyclic peptides. Plant Cell Tissue Organ Cult. 2006, 85, 247–255. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Craik, D.J. Host-Defense Activities of Cyclotides. Toxins 2012, 4, 139-156. https://doi.org/10.3390/toxins4020139
Craik DJ. Host-Defense Activities of Cyclotides. Toxins. 2012; 4(2):139-156. https://doi.org/10.3390/toxins4020139
Chicago/Turabian StyleCraik, David J. 2012. "Host-Defense Activities of Cyclotides" Toxins 4, no. 2: 139-156. https://doi.org/10.3390/toxins4020139
APA StyleCraik, D. J. (2012). Host-Defense Activities of Cyclotides. Toxins, 4(2), 139-156. https://doi.org/10.3390/toxins4020139