Biocontrol of Penicillium nordicum Growth and Ochratoxin A Production by Native Yeasts of Dry Cured Ham

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection and Identification of Yeast Species
2.2. Preparation of Yeast Inocula
2.3. Preparation of P. nordicum Inocula
2.4. Inhibition Assay of P. nordicum by Yeast Strains

2.5. OTA Production by P. nordicum
2.6. OTA Recovery in Yeast Treated Plates
2.7. Statistical Analysis
3. Results
3.1. Yeast Inhibition Activity Against P. nordicum


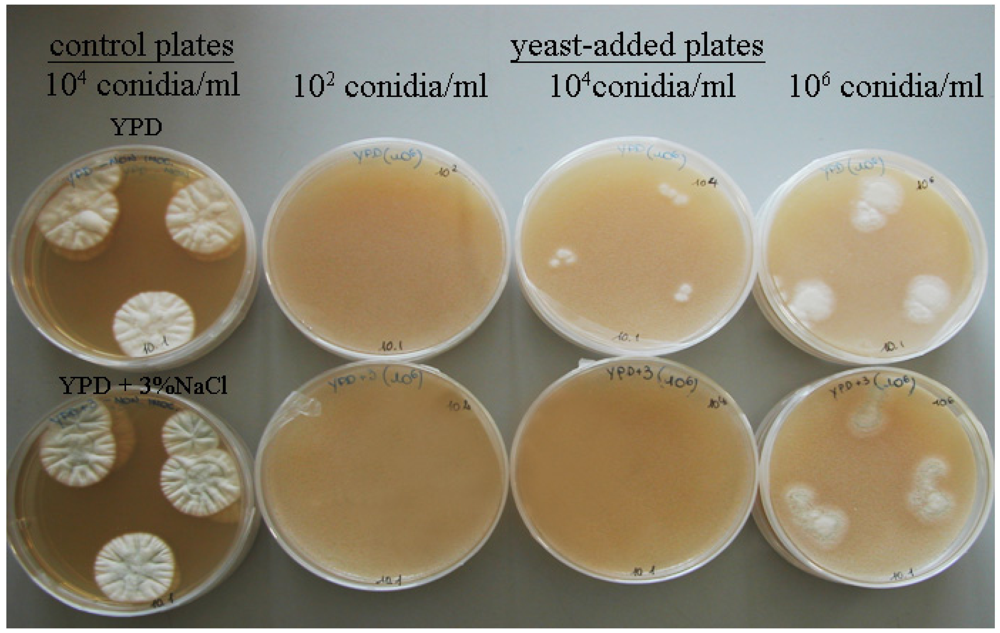{kind=link}
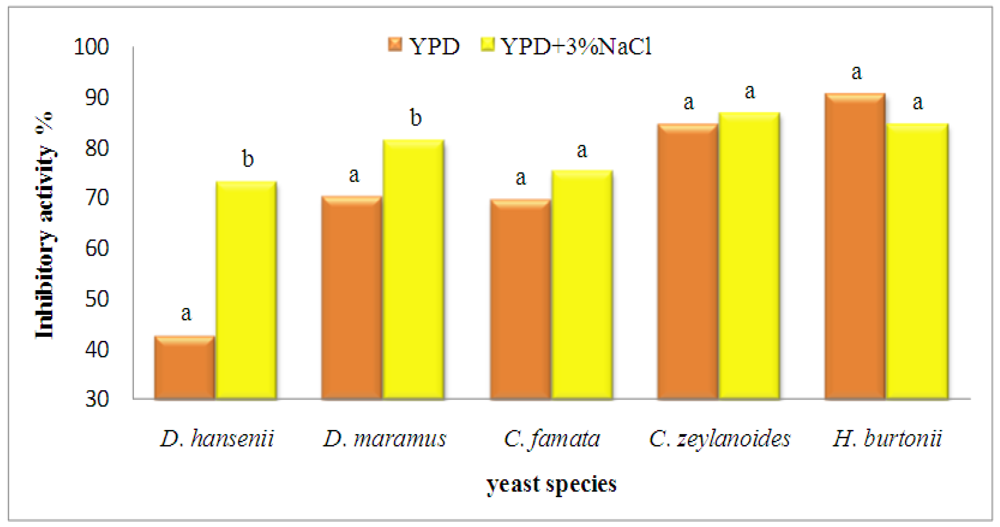{kind=link}
{kind=link}
| Yeast species | Test | P. nordicum a | ||
|---|---|---|---|---|
| 102 | 104 | 106 | ||
| D. hansenii | C | 35.5 ± 2.0 | 41.6 ± 2.2 | 42.4 ± 2.4 |
| Y | 12.5 ± 3.5 | 20.1 ± 3.2 | 23.7 ± 3.5 | |
| D. maramus | C | 37.1 ± 2.0 | 41.7 ± 2.2 | 46.2 ± 1.4 |
| Y | 3.6 ± 1.3 | 11.4 ± 1.9 | 18.0 ± 1.8 | |
| C. famata | C | 40.4 ± 1.4 | 44.4 ±1.4 | 46.6 ± 1.4 |
| Y | 7.1 ± 1.4 | 14.0 ± 2.8 | 15.9 ± 2.1 | |
| C. zeylanoides | C | 34.4 ± 1.9 | 39.6 ± 2.0 | 41.7 ± 2.1 |
| Y | 2.1 ± 1.0 | 5.7 ± 2.2 | 8.8 ± 2.3 | |
| H. burtonii | C | 35.4 ± 2.2 | 39.5 ± 2.0 | 41.9 ± 2.1 |
| Y | 0.7 ± 0.4 | 5.6 ± 1.8 | 8.8 ± 1.8 | |
| Item | % Inhibitory activity | P-value | |
|---|---|---|---|
| YS | 0.000 | ||
| D. hansenii | 58.2 c | ||
| D. maramus | 75.9 b | ||
| C. famata | 72.6 b | ||
| C. zeylanoides | 85.9 a | ||
| H. burtonii | 87.9 a | ||
| PN | 0.000 | ||
| 102 conidia/mL | 87.9 a | ||
| 104 conidia/mL | 73.2 b | ||
| 106 conidia/mL | 67.1 b | ||
| S | 0.000 | ||
| 0% NaCl | 71.7 b | ||
| 3% NaCl | 80.5 a | ||
| Interaction YS × S | 0.000 | ||
| Interaction YS × PN | 0.724 | ||
| Interaction S × PN | 0.803 | ||

3.2. OTA Production in Control and Yeast-Added Plates
| OTA (ppb) | ||||
|---|---|---|---|---|
| Mean | Median | 95th percentile | ||
| Control plates three levels of P. nordicum inoculum as conidia/mL | 102 | 2353 | 2518 | 4523 |
| 104 | 1652 | 1587 | 3429 | |
| 106 | 1064 | 1000 | 2432 | |
| Yeast-added plates three levels of P. nordicum inoculum as conidia/mL + 108 cfu/mL yeasts (Y) | 102 + Y | 33.3 | 4.0 | 187 |
| 104 + Y | 35.2 | 4.0 | 138 | |
| 106 + Y | 42.7 | 7.5 | 179 | |
| Yeast species | OTA | C | Y |
|---|---|---|---|
| D. maramus | L | 0 | 69 |
| M | 29 | 31 | |
| H | 71 | 0 | |
| D. hansenii | L | 0 | 47 |
| M | 35 | 53 | |
| H | 65 | 0 | |
| C. famata | L | 0 | 58 |
| M | 40 | 42 | |
| H | 60 | 0 | |
| C. zeylanoides | L | 0 | 75 |
| M | 35 | 25 | |
| H | 65 | 0 | |
| H. burtonii | L | 0 | 86 |
| M | 33 | 14 | |
| H | 67 | 0 |
| D. maramus | C. famata | C. zeylanoides | H. burtonii | |
|---|---|---|---|---|
| D. hansenii | 0.000 | 0.310 | 0.000 | 0.000 |
| D. maramus | 0.092 | 0.260 | 0.032 | |
| C. famata | 0.006 | 0.000 | ||
| C. zeylanoides | 0.135 |

| medium | Yeasts (cfu/mL) | OTA (ppb) | Recovery (%) |
|---|---|---|---|
| YPD | N | 405 ± 25 | 81 |
| Y | 375 ± 17 | 75 | |
| YPD +3% NaCl | N | 390 ± 15 | 78 |
| Y | 425 ± 13 | 85 |
4. Discussion
5. Conclusions
References
- Martin, M.; Cordoba, J.J.; Aranda, E.; Cordoba, M.G.; Asensio, M.A. Contribution of selected fungal population to the volatile compounds of dry -cured ham. Int. J. Food Microbiol. 2006, 110, 8–13. [Google Scholar]
- Andrade, M.J.; Cordoba, J.J.; Sánchez, B.; Casado, E.M.; Rodríguez, M. Evaluation and selection of yeasts isolated from dry-cured Iberian ham by their volatile compound production. Food Chem. 2009, 113, 457–463. [Google Scholar]
- Spotti, E.; Berni, E.; Cacchioli, C. Characteristics and Applications of Molds. In Meat Biotechnology, 1st; Toldrá, F., Ed.; Springer: New York, NY, USA, 2008; pp. 181–195. [Google Scholar]
- Asefa, D.T.; Kure, C.F.; Gjerde, R.O.; Omer, M.K.; Langsrud, S.; Nesbakken, T.; Skaar, I. Fungal growth pattern, sources and factors of mould contamination in dry-cured meat production process. Int. J. Food Microbiol. 2010, 140, 131–135. [Google Scholar]
- Battilani, P.; Pietri, A.; Giorni, P.; Formenti, S.; Bertuzzi, T.; Toscani, T.; Virgili, R.; Kozakiewicz, Z. Penicillium population in dry-cured ham manufacturing plants. J. Food Prot. 2007, 70, 975–980. [Google Scholar]
- Nunez, F.; Rodriguez, M.M.; Bermudez, M.E.; Cordoba, J.J.; Asensio, M.A. Composition and toxigenic potential of the mould population on dry-cured Iberian ham. Int. J. Food Microbiol. 1996, 32, 185–197. [Google Scholar]
- Comi, G.; Orlic, S.; Redzepovic, S.; Urso, R.; Iacumin, L. Moulds isolated from Istrian dried ham at the pre-ripening and ripening level. Int. J. Food Microbiol. 2004, 96, 29–34. [Google Scholar]
- Wang, X.; Ma, P.; Jiang, D.; Peng, Q.; Yang, H. The natural microflora of Xuanwei ham and the no-mouldy ham production. J. Food Eng. 2006, 77, 103–111. [Google Scholar]
- Castellari, C.; Quadrelli, A.M.; Laich, F. Surface mycobiota on Argentinean dry fermented sausages. Int. J. Food Microbiol. 2010, 142, 149–155. [Google Scholar]
- Larsen, T.O.; Svendsen, A.; Smedsgaard, J. Biochemical characterization of ochratoxin A-producing strains of the genus Penicillium. Appl. Environ. Microbiol. 2001, 67, 3630–3635. [Google Scholar] [CrossRef]
- Pfohl-Leszkowicz, A. Ochratoxin A and aristolochic acid involvement in nephropathies and associated urothelial tract tumors. Arh. Hig. Rada Toksikol. 2009, 60, 465–483. [Google Scholar]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium subgenus Penicillium: A guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 49, 1–173. [Google Scholar]
- Pietri, A.; Bertuzzi, T.; Gualla, A.; Piva, G. Occurrence of ochratoxin A in raw ham muscles and in pork products from northern Italy. Ital. J. Food Sci. 2006, 18, 1–8. [Google Scholar]
- Battilani, P.; Formenti, S.; Toscani, T.; Virgili, R. Influence of abiotic parameters on ochratoxin A production by Penicillium nordicum strain in dry-cured meat model systems. Food Control 2010, 21, 1739–1744. [Google Scholar] [CrossRef]
- Simoncini, N.; Rotelli, D.; Virgili, R.; Quintavalla, S. Dynamics and characterization of yeasts during ripening of typical Italian dry-cured ham. Food Microbiol. 2007, 24, 577–584. [Google Scholar]
- Asefa, D.T.; Møretrø, T.; Gjerde, R.O.; Langsrud, S.; Kure, C.F.; Sidhu, M.S.; Nesbakken, T.; Skaar, I. Yeast diversity and dynamics in the production processes of Norwegian dry-cured meat products. Int. J. Food Microbiol. 2009, 133, 135–140. [Google Scholar]
- Nunez, F.; Rodriguez, M.M.; Cordoba, J.J.; Bermudez, M.E.; Asensio, M.A. Yeast population during ripening of dry-cured Iberian ham. Int. J. Food Microbiol. 1996, 29, 271–280. [Google Scholar]
- Levenson, D.; Pfaller, M.A.; Smith, M.A.; Hollis, R.; Gerarden, T.; Tucci, C.B.; Isenberg, H.D. Candida zeylanoides: an opportunistic yeast. J. Clin. Microbiol. 1991, 29, 1689–1692. [Google Scholar]
- Simoncini, N.; Virgili, R.; Quintavalla, S.; Formenti, S.; Battilani, P. Biotypization of autochthonous yeasts of dry-cured meat products. Ind. Conserve 2009, 84, 107–114. [Google Scholar]
- Pinna, A.; Quintavalla, S.; Simoncini, N.; Toscani, T.; Virgili, R. Volatile organic compounds of a ham-like model system inoculated with authoctonous yeasts isolated from typical hams. Ind. Conserve 2009, 84, 91–103. [Google Scholar]
- Petersson, S.; Hansen, M.W.; Axberg, K.; Hult, K.; Schnürer, J. Ochratoxin A accumulation in cultures of Penicillium verrucosum with the antagonistic yeast Pichia anomala and Saccharomyces cerevisiae. Mycol. Res. 1998, 102, 1003–1008. [Google Scholar] [CrossRef]
- Druvefors, U.A.; Schnürer, J. Mold-inhibitory activity of different yeast species during airtight storage of wheat grain. FEMS Yeast Res. 2005, 5, 373–378. [Google Scholar]
- Masoud, W.; Kaltoft, C.H. The effects of yeasts involved in the fermentation of Coffea arabica in East Africa on growth and ochratoxin A (OTA) production by Aspergillus ochraceus. Int. J. Food Microbiol. 2006, 106, 229–234. [Google Scholar] [CrossRef]
- Guinebretiere, M.H.; Nguyen-The, C.; Morrison, N.; Reich, M.; Nicot, P. Isolation and characterization of antagonists for the biocontrol of the postharvest wound pathogen Botrytis cinerea on strawberry fruits. J. Food Prot. 2000, 3, 386–394. [Google Scholar]
- Suzzi, G.; Romano, P.; Ponti, I.; Montuschi, C. Natural wine yeasts as biocontrol agents. J. Appl. Bacteriol. 1995, 78, 304–308. [Google Scholar]
- Liu, S.Q.; Tsao, M. Biocontrol of dairy moulds by antagonistic dairy yeast Debaryomyces hansenii in yoghurt and cheese at elevated temperatures. Food Control 2009, 20, 852–855. [Google Scholar] [CrossRef]
- Sánchez-Molinero, F.; Arnau, J. Effect of the inoculation of a starter culture and vacuum packaging (during resting stage) on the appearance and some microbiological and physicochemical parameters of dry-cured ham. Meat Sci. 2008, 79, 29–38. [Google Scholar]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large-subunit (26S) ribosomal DNA partial sequences. Antoine van Leeuwenhoek 1997, 73, 331–371. [Google Scholar]
- Bogs, C.; Battilani, P.; Geisen, R. Development of a molecular detection and differentiation system for ochratoxin A producing Penicillium species and its application to analyse the occurrence of P. nordicum in cured meats. Int. J. Food Microbiol. 2006, 107, 39–47. [Google Scholar] [CrossRef]
- Bleve, G.; Grieco, F.; Logrieco, A.; Visconti, A. Isolation of epiphytic yeasts with potential for biocontrol of Aspergillus carbonarius and A. niger on grape. Int. J. Food Microbiol. 2006, 108, 204–209. [Google Scholar] [CrossRef]
- Lima, G.; Arru, S.; De Curtis, F.; Arras, G. Influence of antagonistic, host fruit and pathogen on the biological control of postharvest fungal diseases by yeasts. J. Ind. Microbiol. Biotechnol. 1999, 23, 223–229. [Google Scholar]
- Bragulat, M.R.; Abarca, M.L.; Cabañes, F.J. An easy screening method for fungi producing ochratoxin A in pure culture. Int. J. Food Microbiol. 2001, 71, 139–144. [Google Scholar]
- Toscani, T.; Moseriti, A.; Dossena, A.; Dall’Asta, C.; Simoncini, N.; Virgili, R. Determination of ochratoxin A in dry-cured meat products by HPLC-FLD quantitative methods. J. Chromatogr. B 2007, 855, 242–248. [Google Scholar]
- Asefa, D.T.; Gjerde, R.O.; Sidhu, M.S.; Langsrud, S.; Kure, C.F.; Nesbakken, T.; Skaar, I. Moulds contaminants on Norwegian dry-cured meat products. Int. J. Food Microbiol. 2009, 128, 435–439. [Google Scholar]
- Spotti, E.; Berni, E.; Cacchioli, C.; Simoncini, N.; Quintavalla, S. Growth and antagonistic activity of Hyphopichia burtonii against other fungal species frequently found on meat products during maturation. Ind. Conserve 2009, 84, 11–22. [Google Scholar]
- Björnberg, A.; Schnürer, J. Inhibition of the growth of the grain-storage molds in vitro by the yeast Pichia anomala (Hansen) Kurtzman. Can. J. Microbiol. 1993, 39, 623–628. [Google Scholar] [CrossRef]
- Breuer, U.; Harms, H. Debaryomyces hansenii—an extremophilic yeast with biotechnological potential. Yeast 2006, 23, 415–437. [Google Scholar] [CrossRef]
- Ramakrishna, N.; Lacey, J.; Smith, J.E. Colonization of barley grain by Penicillium verrucosumand Ochratoxin A formation in the presence of competing fungi. J. Food Prot. 1996, 59, 1311–1317. [Google Scholar]
- Metaxopoulos, J.; Stavropoulos, S.; Kakouri, A.; Samelis, J. Yeasts isolated from traditional Greek dry salami. Ital. J. Food Sci. 1996, 1, 25–32. [Google Scholar]
- Monte, E.; Villanueva, J.R.; Domìnguez, A. Fungal profiles of Spanish country-cured hams. Int. J. Food Microbiol. 1986, 3, 355–359. [Google Scholar]
- Arras, G. Mode of action of an isolate of Candida famata in biological control of Penicillium digitatum in orange fruits. Postharvest Biol. Technol. 1996, 8, 191–198. [Google Scholar] [CrossRef]
- Sonjak, S.; Ličen, M.; Frisvad, J.C.; Gunde-Cimerman, N. The mycobiota of three dry-cured meat products from Slovenia. Food Microbiol. 2011, 28, 373–376. [Google Scholar]
- Asefa, D.; Kure, C.F.; Gjerde, R.O.; Langsrud, S.; Omer, M.O.; Nesbakken, T.; Skaar, I. A HACCP plan for mycotoxigenic hazards associated with dry-cured meat production processes. Food Control 2010, 22, 831–837. [Google Scholar]
- Llorente, P.; Marquina, D.; Santos, A.; Peinado, J.M.; Spencer-Martins, I. Effect of salt on the killer phenotype of yeasts from olive brines. Appl. Environ. Microbiol. 1997, 63, 1165–1167. [Google Scholar]
- Marquinia, D.; Barroso, J.; Santos, A.; Peinado, J.M. Production and characteristics of Debaryomyces hansenii killer toxin. Microbiol. Res. 2001, 156, 387–391. [Google Scholar] [CrossRef]
- Patharajan, S.; Reddy, K.R.N.; Spadaro, D.; Lore, A.; Gullino, M.L.; Garibaldi, A.; Karthikeyan, V. Potential of yeast antagonists on in vitro biodegradation of ochratoxin A. Food Control 2011, 22, 290–296. [Google Scholar] [CrossRef]
- Caridi, A. New perspectives in safety and quality enhancement of wine through selection of yeasts based on the parietal adsorption activity. Int. J. Food Microbiol. 2007, 120, 167–172. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Virgili, R.; Simoncini, N.; Toscani, T.; Camardo Leggieri, M.; Formenti, S.; Battilani, P. Biocontrol of Penicillium nordicum Growth and Ochratoxin A Production by Native Yeasts of Dry Cured Ham. Toxins 2012, 4, 68-82. https://doi.org/10.3390/toxins4020068
Virgili R, Simoncini N, Toscani T, Camardo Leggieri M, Formenti S, Battilani P. Biocontrol of Penicillium nordicum Growth and Ochratoxin A Production by Native Yeasts of Dry Cured Ham. Toxins. 2012; 4(2):68-82. https://doi.org/10.3390/toxins4020068
Chicago/Turabian StyleVirgili, Roberta, Nicoletta Simoncini, Tania Toscani, Marco Camardo Leggieri, Silvia Formenti, and Paola Battilani. 2012. "Biocontrol of Penicillium nordicum Growth and Ochratoxin A Production by Native Yeasts of Dry Cured Ham" Toxins 4, no. 2: 68-82. https://doi.org/10.3390/toxins4020068
APA StyleVirgili, R., Simoncini, N., Toscani, T., Camardo Leggieri, M., Formenti, S., & Battilani, P. (2012). Biocontrol of Penicillium nordicum Growth and Ochratoxin A Production by Native Yeasts of Dry Cured Ham. Toxins, 4(2), 68-82. https://doi.org/10.3390/toxins4020068