1. Introduction
Members of the fungal family of
Clavicipitaceae infest grass species, including cereal grains, and are capable of producing a number of so-called ergot alkaloids, which might exert adverse effects on livestock [
1]. For monogastric animals, such as pigs, especially the species
Claviceps purpurea is of special interest, as it infects mainly rye, wheat and triticale and develops mycelium instead of kernels on the ears. These mycelia appear as discolored hardened sclerotia and might contain a total alkaloid (TA) content between 0.01% and 0.21% composed of individual alkaloids. These alkaloids potentially interact with adrenergic, serotoninergic and dopaminergic receptors, depending on their chemical nature, concentration and proportion to each other [
1]. Besides their pharmacological potential, ergot alkaloids are also capable of influencing the liver, the organ exposed to these substances via the portal vein directly after absorption from the gastro-intestinal tract. The liver is involved in xenobiotic metabolism and was shown to respond to ergocryptine and ergometrine maleate with an increased liver weight and an altered glycogen metabolism of rats [
2,
3,
4]. Moreover, the xenobiotic metabolizing enzymes of the cytochrome P450 (CYP) family were demonstrated to be involved in ergot metabolism [
5] and to be induced [
6]. Moreover, hepatic mitochondrial Ca
2+ ATPase activity and, consequently, ATP formation was found to be inhibited by ergotamine and ergonovine [
7]. It becomes clear that the hepatocyte might be affected, at least at the cytosolic, microsomal and mitochondrial level. Each of these hepatic compartment functions can be tested by non-invasive breath tests using specific carbon-labeled compounds (for reviews, see [
8,
9,
10,
11,
12,
13,
14]). These compartment-specific substrates are primarily metabolized to CO
2 and include, for example,
13C-α-ketoisocaproic acid (KICA),
N-[4-Methoxyphenyl]acetamid (
13C-Methacetin, MC) and
13C-phenylalanine for testing mitochondrial, microsomal and cytosolic compartments, respectively (e.g., [
12]). Effects of ergot on pigs are rather inconsistent [
15] and might be due to the mentioned variation in ergot alkaloid content and pattern [
16]. So far, effects on porcine livers were only evaluated by blood clinical-chemical measures, indicative of hepatocyte function and integrity [
17,
18]. However, these parameters are static and cannot be regarded as reliable markers for global liver function or as a tool to quantify functional hepatic reserve [
12]. Thus, the aim of the present experiment was to test the effects of two ergot batches differing in alkaloid content and pattern on feed intake, weight gain and serum clinical-chemical measures of rearing piglets, along with measurement of the hepatic mitochondrial and microsomal function using the KICA and MC breath test, respectively.
3. Discussion
It has been repeatedly pointed out that an efficient protection of farm animals from ergot, which might be present in feed, can only be achieved when the toxic compounds, i.e., the ergot alkaloids, are analyzed in feed and related to the health and performance of animals. Only by collecting such corresponding data will it be possible to replace the current regulation on the upper limit of 1000 mg ergot per kg unground cereal grains. Thus we aimed at this issue with a special focus on the possible effects of ergot alkaloids on the porcine liver.
Although the TA content of the ergot-containing diets varied from 9 to 22 mg/kg the effects on performance appeared to be independent of both the ergot source and the TA content. However, it needs to be stressed that especially the piglets fed the Ergot 5-high diet, which contained 22 mg TA/kg, were characterized by the lowest feed intake compared to piglets fed all the other ergot-containing diets (9–14 mg TA/kg). Live weight gain over the whole experiment of the latter three groups was 28% to 35% lower than that of the control group, while group Ergot 5-high gained 46% less live weight than the control group, which is equivalent to an additional drop of 11% to 18% compared to the other three ergot fed groups. Due to the variation and the limited number of replications, the differences failed to reach significance. However, putting the data of the present experiment together within the frame of literature findings, a clear linear negative relationship between the dietary TA content and performance of rearing piglets becomes obvious [
20]. Based on this literature compilation, it can be deduced that feed intake, live weight gain and gain-to-feed ratio decrease by 0.9%, 1.24% and 0.9%, respectively, per incremental increase of 1 mg TA/kg diet.
Besides the effects of ergot alkaloids on the gross performance of the piglets, a second aim of the present experiment was to evaluate possible liver effects. Earlier findings revealed that ergot alkaloids affected the nutrient status of the liver [
2,
3,
4]. Moreover, hepatic CYP metabolism of and induction by ergot alkaloids have been demonstrated [
5,
6]. Additionally, the hepatic mitochondrial Ca
2+ ATPase activity was shown to be influenced by ergot alkaloids [
7]. Thus, we hypothesized that ergot alkaloids fed to piglets affect the liver at different subcellular compartments. In order to test this hypothesis, we used KICA and MC as breath test substrates to evaluate the effects of ergot alkaloids on hepatic branched-chain α-ketoacid dehydrogenase (BCKDH, EC 1.2.4.4) and on CYP1A2 (EC 1.14.14.1), respectively.
A literature review about advantages and pitfalls of breath tests for evaluation of the liver functional capacity in general, and of hepatic mitochondrial function in particular, suggested that KICA and methionine are promising substrates [
12]. While the complex metabolic pathways of methionine still have to be studied, with regard to the most appropriate label position, the metabolism of KICA undergoes two main enzymatic pathways, which include the transamination into the corresponding branched-chain amino acid leucine and the oxidative decarboxylation by BCKDH [
12]. The latter enzyme is located in mitochondria, and the main activity is confined to the liver of humans (for reviews see [
11,
12]; rats [
21] and pigs [
22]). The dysfunction of hepatocyte mitochondria is regarded as one of the earliest signs of a dysfunction of the whole organ, as damage to this cellular structure occurs at an earlier stage than in the whole hepatocyte [
23]. Thus, testing this decarboxylase using
13C breath tests is thought to be a tool for detecting effects of xenobiotics [
12,
23]. Among other factors, the liver specificity of the KICA breath test depends on the quantitative proportion of the hepatic BCKDH activity of the total body activity. For pigs, there is experimental evidence about this proportion. It could be demonstrated that liver was characterized by an approximately forty-times higher total activity than muscle when expressed per gram tissue, and it was still eighteen times higher when related to hepatic protein. However, when the total masses of liver and muscles were considered, the contribution of the liver was just 2.3-times higher than that of the muscles [
22]. Interestingly, excessive dietary leucine supply (approximately 50% higher than the requirement) resulted in a significant stimulation of the BCKDH activity both in the liver and in the muscle. For a human KICA breath test, it is generally recommended to supply 20 mg/kg of leucine per person orally in order to prevent or to minimize the transamination pathway of KICA and to force the label to the terminal and irreversible degradation pathway through the BCKDH activity with the intended CO
2 formation. Whether the leucine stimulatory effect on the BCKDH activity interferes with the hepatic effects to be evaluated with the KICA breath test has not been addressed thus far. Originally, the KICA breath test was performed without the supplemental leucine [
21,
24] and could be demonstrated to be sensitive for detection of xenobiotic effects. Later, the KICA was given to humans in the absence and presence of increasing amounts of leucine, and it could be shown that the label recovery (cPDR) was higher in the presence of leucine. However, in spite of the different label recovery, there was a clear correlation of both tests within subjects [
25]. Thus, the detection of treatment effects on BCKDH activity is also possible without supplemental leucine.
In contrast to KICA, the substrate MC is metabolized by the microsomal mixed functional oxidase P4501A2, which exhibits its main activity in the liver, and the clinical relevance of this test lies in the assessment of the hepatic residual functional microsomal mass (for review see [
11]). With regard to the relevance of this P450 isoform, it was deduced from literature findings that all main activities of human CYP isoforms were also found in porcine liver microsomes [
26,
27]. Although no information is available on the total proportion of porcine hepatic P4501A2 of total body activity, the total P450 activity in liver microsomes is considered to be comparable for humans and pigs [
28]. Based on these assumptions, the applicability of the MC breath test to evaluate toxin effects on the porcine liver has been proposed [
29].
The liver specificity of both breath tests also depends on the hepatic extraction rate (difference between hepatic substance inflow and outflow concentration divided by the inflow concentration) of the test substrates in relation to blood flow [
11]. Ideal substrates should be characterized by low hepatic extraction rates, which make the test less dependent on hepatic blood flow. Both test substrates used in the present experiment are efficiently extracted by the liver. MC is regarded as a high extraction rate substance (>0.8) in humans [
11], while the KICA hepatic extraction rate amounts to approximately 0.6 in rats [
30]. Thus, differences in blood flow, caused either by individual variation or by treatment effects, need to be considered with regard to overall variation in discussing the results. General effects of ergot alkaloids on blood circulation could be expected based on their potential interactions with adrenergic, serotoninergic and dopaminergic receptors [
1]. However, in the view that breath test results did not significantly differ in ergot-exposed piglets from the control piglets, there were obviously no adverse effects both on the tested mitochondrial and microsomal functions and on hepatic blood flow.
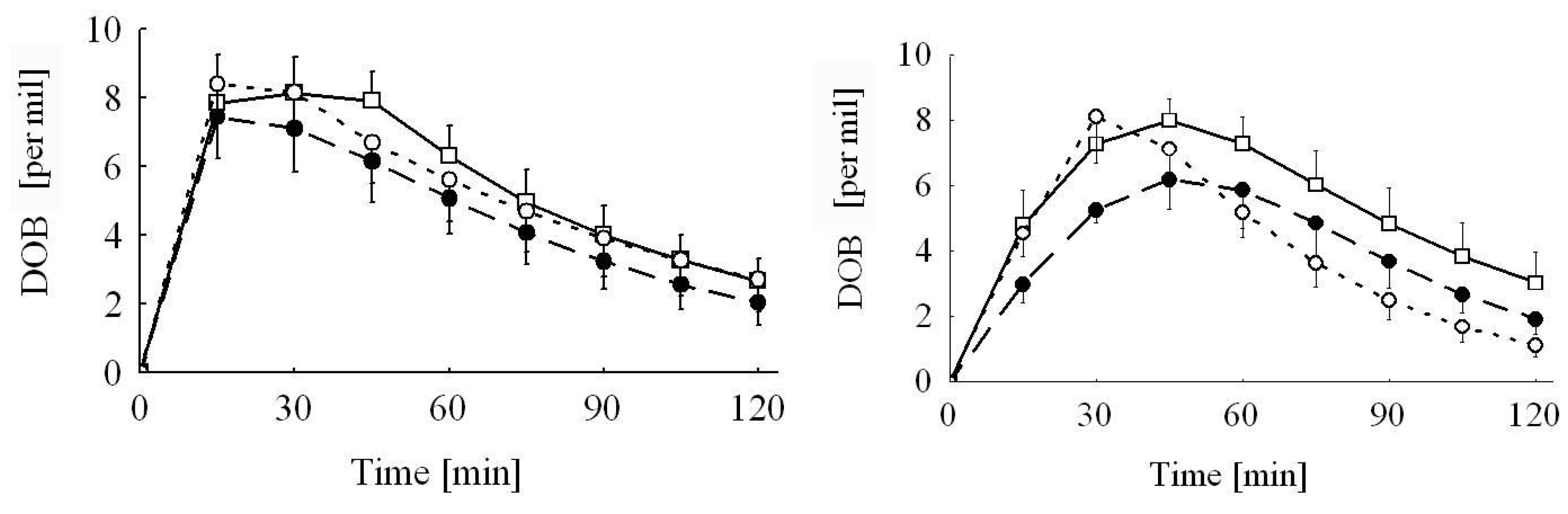
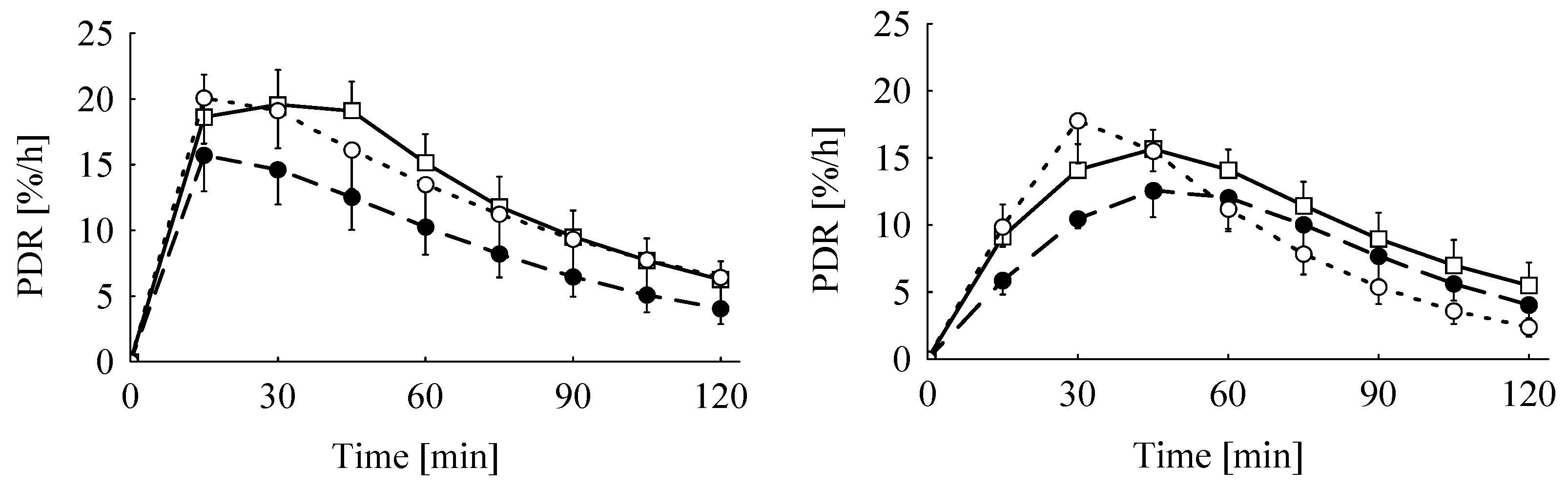
The significant interactions between breath test substrate and sex were mainly caused by the significantly prolonged elimination half-life (
t0.5) in male piglets fed the Ergot 5-high diet when tested with KICA compared to all other sub-groups independent of test substrate, treatment group or sex. The reasons for this isolated prolongation cannot be explained by other parameters recorded in the experiment. As a consequence of the prolonged tracer elimination, these piglets exhibited the lowest cPDR. Other piglets also tested with KICA similarly showed a lower cPDR, particularly during the first hour after the administration of the substrate. This initially retarded tracer recovery is also reflected by the approximately 18 min delayed tracer climax (
tmax) estimated for the KICA breath test. These marked differences were probably not caused by differences in gastric emptying, as both test substrates were administered via a probe directly into the stomach. The smoother increase in KICA oxidation (see
Figure 1) and the later
tmax might be caused by at least two reasons. Firstly, the mitochondrial and microsomal kinetics of KICA and MC oxidation, respectively, might be different due to the underlying differences in metabolic pathways and biochemical events at these subcellular fractions. Secondly, a part of the labeled KICA might temporarily be retained as labeled leucine via the transamination pathway and a subsequent leucine oxidation, which again occurs via KICA. Evidence for the latter explanation comes from human and rat breath test studies employing both
13C-KICA and
13C-leucine as test substrates [
25]. Leucine oxidation climaxed approximately 15 to 20 min later than KICA (55
vs. 35 min in males and 50
vs. 35 min in females). A similar delay in
tmax for oxidized leucine relative to KICA has been observed in rats [
31]. Moreover, hypothyroid rats used in these experiments showed decreased energy expenditure and an increased KICA and leucine oxidation as measured by
13C-breath tests with substrates administered intravenously. These results further indicate that the thyroid status obviously influences the leucine pools, the protein turnover and, finally, the degree of KICA transamination and decarboxylation rate and might consequently contribute to the overall variation as observed with the KICA breath test.
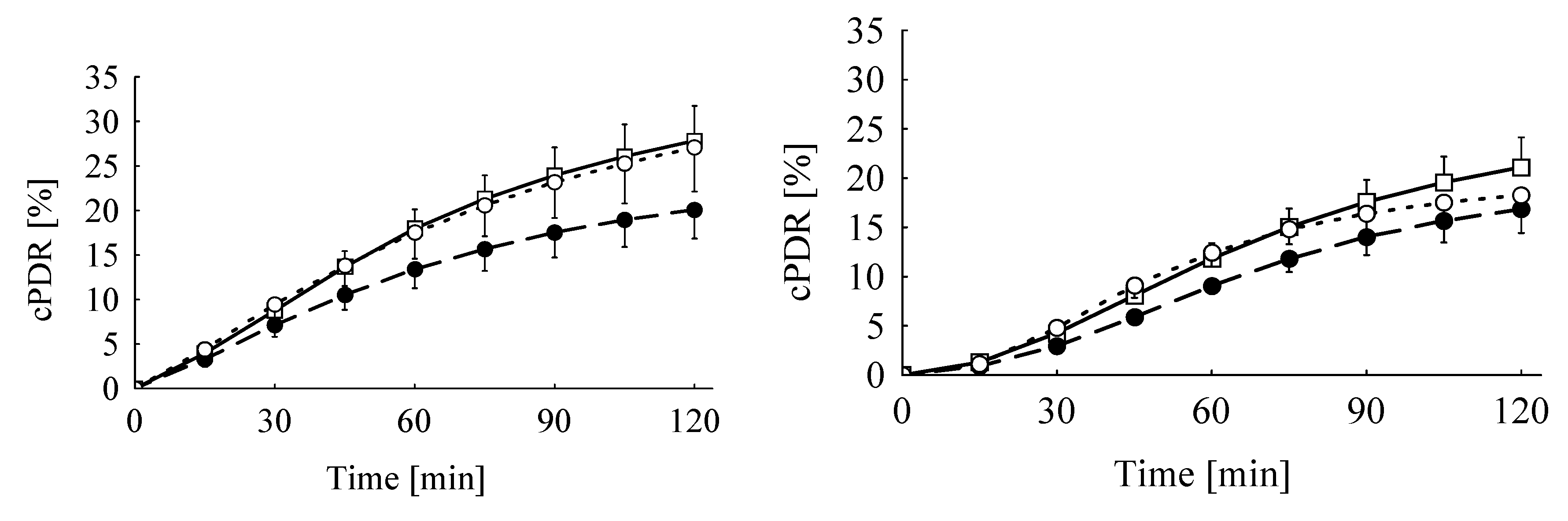
Although feeding the diet Ergot 5-high with the higher TA content of 22 mg/kg tended to decrease the cPDR60 by 26% and 28% in the MC and KICA breath test, respectively, this difference to the control groups failed to reach significance. Furthermore, feeding the Ergot 15-high diet with a TA concentration of 13.5 mg/kg apparently did not differ from the corresponding control groups, both according to the MC and KICA breath test, respectively (−1% vs. +5%). Therefore, both tests discriminated between treatments to the same extent, and the missing significance of the interactions between dietary treatment and test substrate underlines this conclusion.
In reviewing the usefulness of
13C breath tests for diagnosing liver fibrosis, it was concluded that a significant proportion of patients suffering from chronic viral hepatitis and a few suffering from non-alcoholic fatty liver disease were characterized by normal aminotransferase levels despite significant hepatic lesions [
32]. Therefore, it was also of interest for the present experiment to correlate the serum-clinical parameters more or less indicative for liver health with the results of the breath tests (
Table 6 and
Table 7). With the exception of serum albumin concentration in the KICA breath test, none of the other parameters correlated with the results of the breath test. Therefore, the variation observed in the breath test results cannot be explained by variation in the GGT, ALAT, ASAT and GLDH activities in serum. As no correlation was observed between the serum albumin concentration and the breath test results according to the MC breath test, the significant medium positive correlation in the KICA breath test is difficult to explain.
Significant treatment effects were observed for the serum bilirubin concentration. Among others, the total bilirubin level in serum is determined by the balance between the degree of hemoglobin degradation and its elimination with the bile via the liver. Therefore, increased total bilirubin concentrations might result from an increased hemolysis and/or a compromised hepatic bile acid formation, conjugation and elimination. However, neither a cholestatic condition nor other hepatocellular damages as indicated by the unaltered GGT and the GLDH and ALAT activity, respectively, were detected. In addition, the results of the breath tests further substantiate the view that the increased total bilirubin concentrations were probably not related to the liver bile formation process, but might be associated to the overnight starvation of the piglets before blood was collected and the breath test was performed. This view is supported by the negative correlation of −0.5 between total bilirubin and glucose concentration, which partially caused the significant group differences for the serum glucose concentrations (
Table 6 and
Table 7). As a group feeding system was used for piglet keeping, the individual starvation times could only partially be controlled. The higher feed intake level of the control group might explain why these piglets responded more sensitively than ergot-fed piglets, which were generally characterized by a lower feed intake (
Table 3).



{kind=link}
{kind=link}
{kind=link}



