Heparin-Binding Epidermal Growth Factor-like Growth Factor/Diphtheria Toxin Receptor in Normal and Neoplastic Hematopoiesis

Abstract
:1. HB-EGF


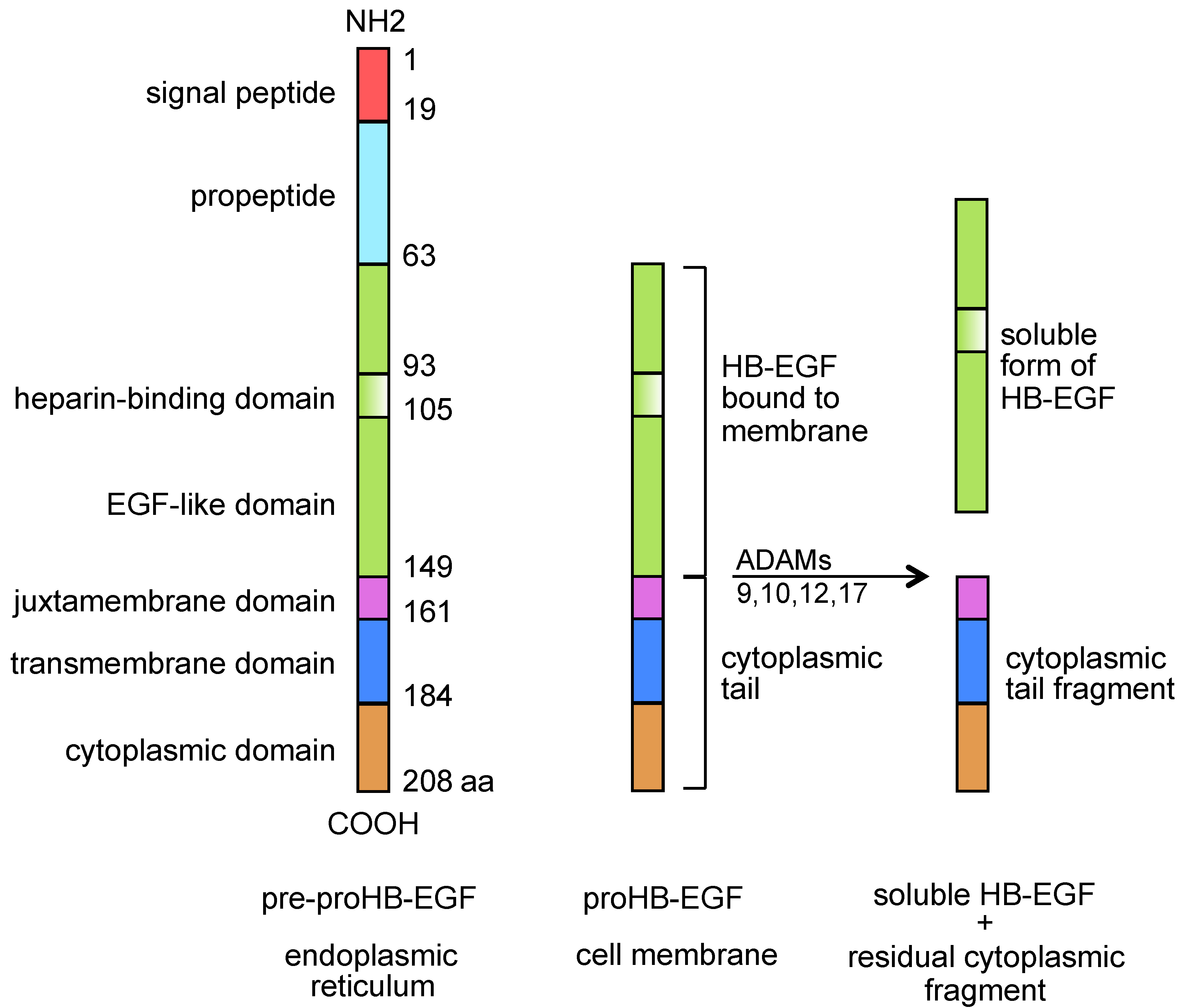{kind=link}
{kind=link}
| EGFR/ErbB1/HER1 | ErbB2/HER2 | ErbB3/HER3 | ErbB4/HER4 |
|---|---|---|---|
| HB-EGF | NRG1/2 | HB-EGF | |
| EGF | NRG1/2/3/4 | ||
| TGF-α | BTC | ||
| AR | EPR | ||
| BTC | |||
| EPR | |||
| ErbB/HER homo- and heterodimers activated by HB-EGF 1/1 - 1/2 - 1/3 - 1/4 - 2/4 - 3/4 - 4/4 | |||
2. EGF Family Members, Cognate Receptors, and Layers of Regulation
3. HB-EGF-Dependent EGFR Phosphorylation and Transduction Pathways
4. GPCR-Dependent Transactivation of EGFR
5. GPCR-Dependent Transinhibition of EGFR
6. Factors Inducing HB-EGF in Hematopoietic Cells
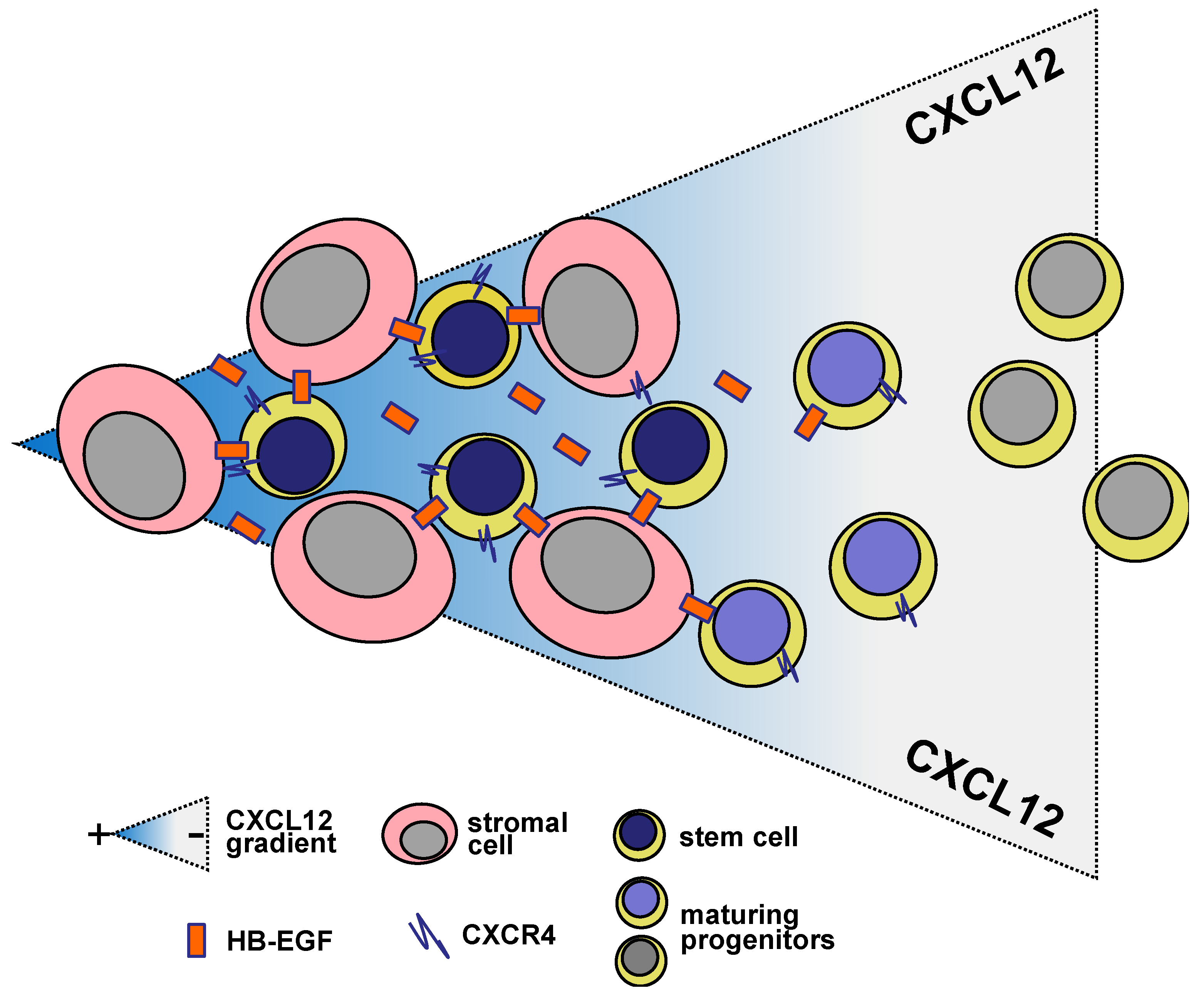
7. HB-EGF in Hematopoietic Microenvironment

8. CXCL12- and HB-EGF-Related Loops in Hematopoiesis
9. HB-EGF in Leukemia
10. HB-EGF in Multiple Myeloma
11. Final Remarks
Acknowledgments
Funding
Conflict of Interest
References
- Higashiyama, S.; Abraham, J.A.; Miller, J.; Fiddes, J.C.; Klagsbrun, M. A heparin-binding growth factor secreted by macrophage-like cells that is related to EGF. Science 1991, 251, 936–939. [Google Scholar] [CrossRef]
- Higashiyama, S.; Lau, K.; Besner, G.E.; Abraham, J.A.; Klagsbrun, M. Structure of heparin-binding EGF-like growth factor. Multiple forms, primary structure, and glycosylation of the mature protein. J. Biol. Chem. 1992, 267, 6205–6212. [Google Scholar]
- Naglich, J.G.; Metherall, J.E.; Russel, D.W.; Eidels, L. Expression cloning of a diphtheria toxin receptor: identity with a heparin-binding EGF-like growth factor precursor. Cell 1992, 69, 1051–1061. [Google Scholar] [CrossRef]
- Iwamoto, R.; Higashiyama, S.; Mitamura, T.; Taniguchi, N.; Klagsbrun, M.; Mekada, E. Heparin-binding EGF-like growth factor, which acts as the diphtheria toxin receptor, forms a complex with membrane protein DRAP27/CD9, which up-regulates functional receptors and diphtheria toxin sensitiviy. EMBO J. 1994, 13, 2322–2330. [Google Scholar]
- Mitamura, T.; Higashiyama, S.; Taniguchi, N.; Klagsbrun, M.; Mekada, E. Diphtheria toxin binds to the epidermal growth factor (EGF)-like domain of human heparin-binding EGF-like growth factor/diphtheria toxin receptor and inhibits specifically its mitogenic activity. J. Biol. Chem. 1995, 270, 1015–1019. [Google Scholar]
- Chang, M.P.; Bramhall, J.; Graves, S.; Bonavida, B.; Wisnieski, B.J. Internucleosomal DNA cleavage precedes diphtheria toxin-induced cytolysis. Evidence that cell lysis is not a simple consequence of translation inhibition. J. Biol. Chem. 1989, 264, 15261–15267. [Google Scholar]
- Blotnick, S.; Peoples, G.E.; Freeman, M.R.; Eberlein, T.J.; Klagsbrun, M. T lymphocytes synthesize and export heparin-binding epidermal growth factor-like growth factor and basic fibroblast growth factor, mitogens for vascular cells and fibroblasts: differential production and release by CD4+ and CD8+ T cells. Proc. Natl. Acad. Sci. USA 1994, 91, 2890–2894. [Google Scholar]
- Peoples, G.E.; Blotnick, S.; Takahashi, K.; Freeman, M.R.; Klagsbrun, M.; Eberlein, T.J. T lymphocytes that infiltrate tumors and atherosclerotic plaques produce heparin-binding epidermal growth factor-like growth factor and basic fibroblast growth factor: a potential pathologic role. Proc. Natl. Acad. Sci. USA 1995, 92, 6547–6551. [Google Scholar]
- Vinante, F.; Marchi, M.; Rigo, A.; Scapini, P.; Pizzolo, G.; Cassatella, M.A. Granulocyte-macrophage colony-stimulating factor induces expression of heparin-binding epidermal growth factor-like growth factor/diphtheria toxin receptor and sensitivity to diphtheria toxin in human neutrophils. Blood 1999, 94, 3169–3177. [Google Scholar]
- Powell, P.P.; Klagsbrun, M.; Abraham, J.A.; Jones, R.C. Eosinophils expressing heparin-binding EGF-like growth factor mRNA localize around lung microvessels in pulmonary hypertension. Am. J. Pathol. 1993, 143, 784–793. [Google Scholar]
- Vinante, F.; Rigo, A.; Papini, E.; Cassatella, M.A.; Pizzolo, G. Heparin-binding epidermal growth factor-like growth factor/diphtheria toxin receptor expression by acute myeloid leukemia cells. Blood 1999, 93, 1715–1723. [Google Scholar]
- De Vos, J.; Couderc, G.; Tarte, K.; Jourdan, M.; Requirand, G.; Delteil, M.C.; Rossi, J.F.; Mechti, N.; Klein, B. Identifying intercellular signaling genes expressed in malignant plasma cells by using complementary DNA arrays. Blood 2001, 98, 771–780. [Google Scholar]
- Dluz, S.; Higashiyama, S.; Damm, D.; Abraham, J.A.; Klagsbrun, M. Heparin-binding epidermal growth factor-like growth factor expression in cultured fetal human vascular smooth muscle cells. Induction of mRNA levels and secretion of active mitogen. J. Biol. Chem. 1993, 268, 18330–18334. [Google Scholar]
- Yoshizumi, M.; Kourembanas, S.; Temizer, D.H.; Cambria, R.P.; Quertermous, T.; Lee, M.E. Tumor necrosis factor increases transcription of the heparin-binding epidermal growth factor-like growth factor gene in vascular endothelial cells. J. Biol. Chem. 1992, 267, 9467–9469. [Google Scholar]
- Hashimoto, K.; Higashiyama, S.; Asada, H.; Hashimura, E.; Kobayashi, T.; Sudo, K.; Nakagawa, T.; Damm, D.; Yoshikawa, K.; Taniguchi, N. Heparin-binding epidermal growth factor-like growth factor is an autocrine growth factor for human keratinocytes. J. Biol. Chem. 1994, 269, 20060–20066. [Google Scholar]
- Mishima, K.; Higashiyama, S.; Asai, A.; Yamaoka, K.; Nagashima, Y.; Taniguchi, N.; Kitanaka, C.; Kirino, T.; Kuchino, Y. Heparin-binding epidermal growth factor-like growth factor stimulates mitogenic signaling and is highly expressed in human malignant gliomas. Acta Neuropathol. 1998, 96, 322–328. [Google Scholar]
- Raab, G.; Klagsbrun, M. Heparin-binding EGF-like growth factor. Biochem. Biophys. Acta 1997, 1333, 179–199. [Google Scholar]
- Rigo, A.; Gottardi, M.; Zamò, A.; Mauri, P.; Bonifacio, M.; Krampera, M.; Damiani, E.; Pizzolo, G.; Vinante, F. Macrophages may promote cancer growth via a GM-CSF/HB-EGF paracrine loop that is enhanced by CXCL12. Mol. Cancer 2010, 9, 273. [Google Scholar] [CrossRef]
- Elenius, K.; Paul, S.; Allison, G.; Sun, J.; Klagsbrun, M. Activation of HER4 by heparin-binding EGF-like growth factor stimulates chemotaxis but not proliferation. EMBO J. 1997, 16, 1268–1278. [Google Scholar] [CrossRef]
- Nakamura, K.; Iwamoto, R.; Mekada, E. Membrane-anchored heparin-binding EGF-like growth factor (HB-EGF) and diphtheria toxin receptor-associated protein (DRAP27)/CD9 form a complex with integrin α3β1 at cell-cell contact sites. J. Cell Biol. 1995, 129, 1691–1705. [Google Scholar] [CrossRef]
- Iwamoto, R.; Handa, K.; Mekada, E. Contact-dependent growth inhibition and apoptosis of epidermal growth factor (EGF) receptor-expressing cells by the membrane-anchored form of heparin-binding EGF-like growth factor. J. Biol. Chem. 1999, 274, 25906–25912. [Google Scholar] [CrossRef]
- Takemura, T.; Kondo, S.; Homma, T.; Sakai, M.; Harris, R.C. The membrane-bound form of heparin-binding epidermal growth factor-like growth factor promotes survival of cultured renal epithelial cells. J. Biol. Chem. 1997, 272, 31036–31042. [Google Scholar]
- Miyoshi, E.; Higashiyama, S.; Nakagawa, T.; Hayashi, N.; Taniguchi, N. Membrane-anchored heparin-binding EGF-like growth factor acts as a tumor survival factor in hepatoma cell line. J. Biol. Chem. 1997, 272, 14349–14355. [Google Scholar]
- Higashiyama, S. Metalloproteinase-mediated shedding of heparin-binding EGF-like growth factor and its pathophysiological roles. Protein. Pept. Lett. 2004, 11, 443–450. [Google Scholar] [CrossRef]
- Izumi, Y.; Hirata, M.; Hasuwa, H.; Iwamoto, R.; Umata, T.; Miyado, K.; Tamai, Y.; Kurisaki, T.; Sehara-Fujisawa, A.; Ohno, S.; Mekada, E. A metalloprotease-disintegrin, MDC9/meltrin-γ/ADAM9 and PKCδ are involved in TPA-induced ectodomain shedding of membrane-anchored heparin-binding EGF-like growth factor. EMBO J. 1998, 17, 7260–7272. [Google Scholar] [CrossRef]
- Lemjabbar, H.; Basbaum, C. Platelet-activating factor receptor and ADAM10 mediate responses to Staphylococcus aureus in epithelial cells. Nat. Med. 2002, 8, 41–46. [Google Scholar] [CrossRef]
- Asakura, M.; Kitakaze, M.; Takashima, S.; Liao, Y.; Ishikura, F.; Yoshinaka, T.; Ohmoto, H.; Node, K.; Yoshino, K.; Ishiguro, H.; Asanuma, H.; Sanada, S.; Matsumura, Y.; Takeda, H.; Beppu, S.; Tada, M.; Hori, M.; Higashiyama, S. Cardiac hypertrophy is inhibited by antagonism of ADAM12 processing of HB-EGF: metalloproteinase inhibitors as a new therapy. Nat. Med. 2002, 8, 35–40. [Google Scholar] [CrossRef]
- Sunnarborg, S.W.; Hinkle, C.L.; Stevenson, M.; Russell, W.E.; Raska, C.S.; Peschon, J.J.; Castner, B.J.; Gerhart, M.J.; Paxton, R.J.; Black, R.A.; Lee, D.C. Tumor necrosis factor-alpha converting enzyme (TACE) regulates epidermal growth factor receptor ligand availability. J. Biol. Chem. 2002, 277, 12838–12845. [Google Scholar]
- Yan, Y.; Shirakabe, K.; Werb, Z. The metalloprotease Kuzbanian (ADAM10) mediates the transactivation of EGF receptor by G protein-coupled receptors. J. Cell Biol. 2002, 158, 221–226. [Google Scholar] [CrossRef]
- Abramovitch, R.; Neeman, M.; Reich, R.; Stein, I.; Keshet, E.; Abraham, J.A.; Solomon, A.; Marikovsky, M. Intercellular communication between vascular smooth muscle and endothelial cells mediated by heparin-binding epidermal growth factor-like growth factor and vascular endothelial growth factor. FEBS Lett. 1998, 425, 441–447. [Google Scholar] [CrossRef]
- Higashiyama, S.; Abraham, J.A.; Klagsbrun, M. Heparin-binding EGF-like growth factor stimulation of smooth muscle cell migration: dependence on interactions with cell surface heparan sulfate. J. Cell Biol. 1993, 122, 933–940. [Google Scholar] [CrossRef]
- Faber-Elman, A.; Solomon, A.; Abraham, J.A.; Marikovsky, M.; Schwartz, M. Involvement of wound-associated factors in rat brain astrocyte migratory response to axonal injury: In vitro simulation. J. Clin. Invest. 1996, 97, 162–171. [Google Scholar]
- Hieda, M.; Isokane, M.; Koizumi, M.; Higashi, C.; Tachibana, T.; Shudou, M.; Taguchi, T.; Hieda, Y.; Higashiyama, S. Membrane-anchored growth factor, HB-EGF, on the cell surface targeted to the inner nuclear membrane. J. Cell Biol. 2008, 180, 763–769. [Google Scholar]
- Nanba, D.; Mammoto, A.; Hashimoto, K.; Higashiyama, S. Proteolytic release of the carboxy-terminal fragment of proHB-EGF causes nuclear export of PLZF. J. Cell Biol. 2003, 163, 489–502. [Google Scholar] [CrossRef]
- Nakano, T.; Raines, E.W.; Abraham, J.A.; Klagsbrun, M.; Ross, R. Lysophosphatidylcholine upregulates the level of heparin-binding epidermal growth factor-like growth factor mRNA in human monocytes. Proc. Natl. Acad. Sci. USA 1994, 91, 1069–1073. [Google Scholar]
- Marikovsky, M.; Breuing, K.; Liu, P.Y.; Eriksson, E.; Higashiyama, S.; Farber, P.; Abraham, J.; Klagsbrun, M. Appearance of heparin-binding EGF-like growth factor in wound fluid as a response to injury. Proc. Natl. Acad. Sci. USA 1993, 90, 3889–3893. [Google Scholar] [CrossRef]
- Zhang, Z.; Funk, C.; Glasser, S.R.; Mulholland, J. Progesterone regulation of heparin-binding epidermal growth factor-like growth factor gene expression during sensitization and decidualization in the rat uterus: Effects of the antiprogestin, ZK 98.299. Endocrinology 1994, 135, 1256–1263. [Google Scholar] [CrossRef]
- Freeman, M.R.; Yoo, J.J.; Raab, G.; Soker, S.; Adam, R.M.; Schneck, F.X.; Renshaw, A.A.; Klagsbrun, M.; Atala, A. Heparin-binding EGF-like growth factor is an autocrine growth factor for human urothelial cells and is synthesized by epithelial and smooth muscle cells in the human bladder. J. Clin. Invest. 1997, 99, 1028–1036. [Google Scholar] [CrossRef]
- Freeman, M.R.; Paul, S.; Kaefer, M.; Ishikawa, M.; Adam, R.M.; Renshaw, A.A.; Elenius, K.; Klagsbrun, M. Heparin-binding EGF-like growth factor in the human prostate: synthesis predominantly by interstitial and vascular smooth muscle cells and action as a carcinoma cell mitogen. J. Cell. Biochem. 1998, 68, 328–338. [Google Scholar]
- Arbiser, J.L.; Raab, G.; Rohan, R.M.; Paul, S.; Hirschi, K.; Flynn, E.; Price, E.R.; Fisher, D.E.; Cohen, C.; Klagsbrun, M. Isolation of mouse stromal cells associated with a human tumor using differential diphtheria toxin sensitivity. Am. J. Pathol. 1999, 155, 723–729. [Google Scholar]
- Mehta, V.B.; Besner, G.E. HB-EGF promotes angiogenesis in endothelial cells via PI3-kinase and MAPK signaling pathways. Growth Factors 2007, 25, 253–263. [Google Scholar] [CrossRef]
- Nishi, E.; Prat, A.; Hospital, V.; Elenius, K.; Klagsbrun, M. N-arginine dibasic convertase is a specific receptor for heparin-binding EGF-like growth factor that mediates cell migration. EMBO J. 2001, 20, 3342–3350. [Google Scholar]
- Plowman, G.D.; Culouscou, J.M.; Whitney, G.S.; Green, J.M.; Carlton, G.W.; Foy, L.; Neubauer, M.G.; Shoyab, M. Ligand-specific activation of HER4/p180erbB4, a fourth member of the epidermal growth factor receptor family. Proc. Natl. Acad. Sci. USA 1993, 90, 1746–1750. [Google Scholar]
- Elenius, K.; Corfas, G.; Paul, S.; Choi, C.J.; Rio, C.; Plowman, G.D.; Klagsbrun, M. A novel juxtamembrane domain isoform of HER4/ErbB4. Isoform-specific tissue distribution and differential processing in response to phorbol ester. J. Biol. Chem. 1997, 272, 26761–26768. [Google Scholar]
- Krampera, M.; Pasini, A.; Rigo, A.; Scupoli, M.T.; Tecchio, C.; Malpeli, G.; Scarpa, A.; Dazzi, F.; Pizzolo, G.; Vinante, F. HB-EGF/HER-1 signaling in bone marrow mesenchymal stem cells: inducing cell expansion and reversibly preventing multilineage differentiation. Blood 2005, 106, 59–66. [Google Scholar] [CrossRef]
- Lue, H-W.; Yang, X.; Wang, R.; Qian, W.; Xu, R.Z.H.; Lyles, R.; Osunkoya, A.O.; Zhou, B.P.; Vessella, R.L.; Zayzafoon, M.; Liu, Z-R.; Zhau, H.E.; Chung, L.W.K. LIV-1 Promotes Prostate Cancer Epithelial-to-Mesenchymal Transition and Metastasis through HB-EGF Shedding and EGFR-Mediated ERK Signaling. PLoS One 2011, 6, e27720. [Google Scholar]
- Umeda, Y.; Miyazaki, Y.; Shiinoki, H.; Higashiyama, S.; Nakanishi, Y.; Hieda, Y. Involvement of heparin-binding EGF-like growth factor and its processing by metalloproteinases in early epithelial morphogenesis of the submandibular gland. Dev. Biol. 2001, 237, 202–211. [Google Scholar]
- Chen, X.; Raab, G.; Deutsch, U.; Zhang, J.; Ezzell, R.M.; Klagsbrun, M. Induction of heparin-binding EGF-like growth factor expression during myogenesis. Activation of the gene by MyoD and localization of the transmembrane form of the protein on the myotube surface. J. Biol. Chem. 1995, 270, 18285–18294. [Google Scholar]
- Oyagi, A.; Oida, Y.; Kakefuda, K.; Shimazawa, M.; Shioda, N.; Moriguchi, S.; Kitaichi, K.; Nanba, D.; Yamaguchi, K.; Furuta, Y.; Fukunaga, K.; Higashiyama, S.; Hara, H. Generation and characterization of conditional heparin-binding EGF-Like growth factor knockout mice. PLoS One 2009, 4, e7461. [Google Scholar] [CrossRef]
- Iwamoto, R.; Mekada, E. ErbB and HB-EGF signaling in heart development and function. Cell Struct. Funct. 2006, 31, 1–14. [Google Scholar] [CrossRef]
- Kaneto, H.; Miyagawa, J.; Kajimoto, Y.; Yamamoto, K.; Watada, H.; Umayahara, Y.; Hanafusa, T.; Matsuzawa, Y.; Yamasaki, Y.; Higashiyama, S.; Taniguchi, N. Expression of heparin-binding epidermal growth factor-like growth factor during pancreas development. A potential role of PDX-1 in transcriptional activation. J. Biol. Chem. 1997, 272, 29137–29143. [Google Scholar] [CrossRef]
- Riese, D.J., 2nd; Stern, D.F. Specificity within the EGF family/ErbB receptor family signaling network. Bioessays 1998, 20, 41–48. [Google Scholar] [CrossRef]
- Yarden, Y.; Sliwkowski, M.X. Untangling the ErbB signalling network. Nat. Rev. Mol. Cell Biol. 2001, 2, 127–137. [Google Scholar] [CrossRef]
- Schlessinger, J. Common and distinct elements in cellular signaling via EGF and FGF receptors. Science 2004, 306, 1506–1507. [Google Scholar] [CrossRef]
- Paria, B.C.; Elenius, K.; Klagsbrun, M.; Dey, S.K. Heparin-binding EGF-like growth factor interacts with mouse blastocysts independently of ErbB1: a possible role for heparan sulfate proteoglycans and ErbB4 in blastocyst implantation. Development 1999, 126, 1997–2005. [Google Scholar]
- Iwamoto, R.; Yamazaki, S.; Asakura, M.; Takashima, S.; Hasuwa, H.; Miyado, K.; Adachi, S.; Kitakaze, M.; Hashimoto, K.; Raab, G.; Nanba, D.; Higashiyama, S.; Hori, M.; Klagsbrun, M.; Mekada, E. Heparin-binding EGF-like growth factor and ErbB signaling is essential for heart function. Proc. Natl. Acad. Sci. USA 2003, 100, 3221–3226. [Google Scholar]
- Shishido, Y.; Sharma, K.D.; Higashiyama, S.; Klagsbrun, M.; Mekada, E. Heparin-like molecules on the cell surface potentiate binding of diphtheria toxin to the diphtheria toxin receptor/membrane-anchored heparin-binding epidermal growth factor-like growth factor. J. Biol. Chem. 1995, 270, 29578–29585. [Google Scholar]
- Iwamoto, R.; Mekada, E. Heparin-binding EGF-like growth factor: a juxtacrine growth factor. Cytokine Growth Factor Rev. 2000, 11, 335–344. [Google Scholar] [CrossRef]
- Hieda, M.; Koizumi, M.; Higashi, C.; Tachibana, T.; Taguchi, T.; Higashiyama, S. The cytoplasmic tail of heparin-binding EGF-like growth factor regulates bidirectional intracellular trafficking between the plasma membrane and ER. FEBS Open Bio. 2012, 2, 339–344. [Google Scholar] [CrossRef]
- Kinugasa, Y.; Hieda, M.; Hori, M.; Higashiyama, S. The carboxyl-terminal fragment of pro-HB-EGF reverses Bcl6-mediated gene repression. J. Biol. Chem. 2007, 282, 14797–14806. [Google Scholar] [CrossRef]
- Wang, X.; Mizushima, H.; Adachi, S.; Ohishi, M.; Iwamoto, R.; Makada, E. Cytoplasmic domain phosphorylation of heparin-binding EGF-like growth factor. Cell. Struct. Funct. 2006, 31, 15–27. [Google Scholar] [CrossRef]
- Hynes, N.E.; Lane, H.A. ErbB receptors and cancer: the complexity of targeted inhibitors. Nat. Rev. Cancer 2005, 5, 341–354. [Google Scholar] [CrossRef]
- Kunami, N.; Yotsumoto, F.; Ishitsuka, K.; Fukami, T.; Odawara, T.; Manabe, S.; Ishikawa, T.; Tamura, K.; Kuroki, M.; Miyamoto, S. Antitumor effects of CRM197, a specific inhibitor of HB-EGF, in T-cell acute lymphoblastic leukemia. Anticancer Res. 2011, 31, 2483–2488. [Google Scholar]
- Miyamoto, S.; Yagi, H.; Yotsumoto, F.; Kawarabayashi, T.; Mekada, E. Heparin-binding epidermal growth factor-like growth factor as a novel targeting molecule for cancer therapy. Cancer Sci. 2006, 97, 341–347. [Google Scholar] [CrossRef]
- Yotsumoto, F.; Yagi, H.; Suzuki, S.O.; Oki, E.; Tsujioka, H.; Hachisuga, T.; Sonoda, K.; Kawarabayashi, T.; Mekada, E.; Miyamoto, S. Validation of HB-EGF and amphiregulin as targets for human cancer therapy. Biochem. Biophys. Res. Commun. 2008, 365, 555–561. [Google Scholar]
- Hackel, P.O.; Zwick, E.; Prenzel, N.; Ullrich, A. Epidermal growth factors: critical mediators of multiple receptor pathways. Curr. Opin. Cell Biol. 1999, 11, 184–189. [Google Scholar] [CrossRef]
- Rigo, A.; Gottardi, M.; Damiani, E.; Bonifacio, M.; Ferrarini, I.; Mauri, P.; Vinante, F. CXCL12 and [N33A]CXCL12 in 5637 and HeLa cells: regulating HER1 phosphorylation via calmodulin/calcineurin. PLoS One 2012, 7, e34432. [Google Scholar]
- Saito, T.; Okada, S.; Ohshima, K.; Yamada, E.; Sato, M.; Uehara, Y.; Shimizu, H.; Pessin, J.E.; Mori, M. Differential Activation of Epidermal Growth Factor (EGF) Receptor Downstream Signaling Pathways by Betacellulin and EGF. Endocrinology 2004, 145, 4232–4243. [Google Scholar] [CrossRef]
- Sahin, U.; Weskamp, G.; Kelly, K.; Zhou, H.M.; Higashiyama, S.; Peschon, J.; Hartmann, D.; Saftig, P.; Blobel, C.P. Distinct roles for ADAM10 and ADAM17 in ectodomain shedding of six EGFR ligands. J. Cell Biol. 2004, 164, 769–779. [Google Scholar] [CrossRef]
- Dethlefsen, S.M.; Raab, G.; Moses, M.A.; Adam, R.M.; Klagsbrun, M.; Freeman, M.R. Extracellular calcium influx stimulates metalloproteinase cleavage and secretion of heparin-binding EGF-like growth factor independently of protein kinase C. J. Cell. Biochem. 1998, 69, 143–153. [Google Scholar]
- Prenzel, N.; Zwick, E.; Daub, H.; Leserer, M.; Abraham, R.; Wallasch, C.; Ullrich, A. EGF receptor transactivation by G-protein-coupled receptors requires metalloproteinase cleavage of proHB-EGF. Nature 1999, 402, 884–888. [Google Scholar]
- Daub, H.; Wallasch, C.; Lankenau, A.; Herrlich, A.; Ullrich, A. Signal characteristics of G-protein-transactivated EGF receptor. EMBO J. 1997, 16, 7032–7044. [Google Scholar] [CrossRef]
- Eguchi, S.; Numaguchi, K.; Iwasaki, H.; Matsumoto, T.; Yamakawa, T.; Utsunomiya, H.; Motley, E.D.; Kawakatsu, H.; Owada, K.M.; Hirata, Y.; Marumo, F.; Inagami, T. Calcium-dependent epidermal growth factor receptor transactivation mediates the angiotensin II-induced mitogen-activated protein kinase activation in vascular smooth muscle cells. J. Biol. Chem. 1998, 273, 8890–8896. [Google Scholar] [CrossRef]
- Rajagopal, S.; Rajagopal, K.; Lefkowitz, R.J. Teaching old receptors new tricks: biasing seven-transmembrane receptors. Nat. Rev. Drug Discov. 2010, 9, 373–386. [Google Scholar] [CrossRef]
- Luttrel, L.M.; Daaka, Y.; Lefkowitz, R.J. Regulation of tyrosine cascades by G-protein-coupled receptors. Curr. Opin. Cell Biol. 1999, 11, 177–183. [Google Scholar] [CrossRef]
- Lefkowitz, R.J. Seven transmembrane receptors: something old, something new. Acta Physiol. (Oxf.) 2007, 190, 9–19. [Google Scholar] [CrossRef]
- Daub, H.; Weiss, F.U.; Wallasch, C.; Ullrich, A. Role of transactivation of the EGF receptor in signalling by G-protein-coupled receptors. Nature 1996, 379, 557–560. [Google Scholar] [CrossRef]
- Yamazaki, S.; Iwamoto, R.; Saeki, K.; Asakura, M.; Takashima, S.; Yamazaki, A.; Kimura, R.; Mizushima, H.; Moribe, H.; Higashiyama, S.; Endoh, M.; Kaneda, Y.; Takagi, S.; Itami, S.; Takeda, N.; Yamada, G.; Mekada, E. Mice with defects in HB-EGF ectodomain shedding show severe developmental abnormalities. J. Cell Biol. 2003, 163, 469–475. [Google Scholar] [CrossRef]
- Ohtsu, H.; Dempsey, P.J.; Satoru Eguchi, S. ADAMs as mediators of EGF receptor transactivation by G protein-coupled receptors. Am. J. Physiol. Cell Physiol. 2006, 291, C1–C10. [Google Scholar] [CrossRef]
- Thelen, M.; Thelen, S. CXCR7, CXCR4 and CXCL12: An eccentric trio? J. Neuroimmunol. 2008, 198, 9–13. [Google Scholar] [CrossRef]
- Singh, R.K.; Lokeshwar, B.L. The IL-8-regulated chemokine receptor CXCR7 stimulates EGFR signaling to promote prostate cancer growth. Cancer Res. 2011, 71, 3268–3277. [Google Scholar] [CrossRef]
- Kasina, S.; Scherle, P.A.; Hall, C.L.; Macoska, J.A. ADAM-mediated amphiregulin shedding and EGFR transactivation. Cell Prolif. 2009, 42, 799–812. [Google Scholar] [CrossRef]
- Yan, Y.; Shirakabe, K.; Werb, Z. The metalloprotease Kuzbanian (ADAM10) mediates the transactivation of EGF receptor by G protein–coupled receptors. J. Cell Biol. 2002, 158, 221–226. [Google Scholar] [CrossRef]
- Cook, A.; Hippensteel, R.; Shimizu, S.; Nicolai, J.; Fatatis, A.; Meucci, O. Interactions between chemokines: regulation of fractalkine/CX3CL1 homeostasis by SDF/CXCL12 in cortical neurons. J. Biol. Chem. 2010, 285, 10563–10571. [Google Scholar]
- Pan, D.; Rubin, G.M. Kuzbanian controls proteolytic processing of Notch and mediates lateral inhibition during Drosophila and vertebrate neurogenesis. Cell 1997, 90, 271–280. [Google Scholar] [CrossRef]
- Moll, N.M.; Ransohoff, R.M. CXCL12 and CXCR4 in bone marrow physiology. Expert. Rev. Hematol. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- McGrath, K.E.; Koniski, A.D.; Maltby, K.M.; McGann, J.K.; Palis, J. Embryonic expression and function of the chemokine SDF-1 and its receptor, CXCR4. Dev. Biol. 1999, 213, 442–456. [Google Scholar] [CrossRef]
- Bleul, C.C.; Fuhlbrigge, R.C.; Casasnovas, J.M.; Aiuti, A.; Springer, T.A. A highly efficacious lymphocyte chemoattractant, stromal cell-derived factor 1 (SDF-1). J. Exp. Med. 1996, 184, 1101–1109. [Google Scholar] [CrossRef]
- Vinante, F.; Rigo, A.; Scupoli, MT.; Pizzolo, G. CD30 triggering by agonistic antibodies regulates CXCR4 expression and CXCL12 chemotactic activity in the cell line L540. Blood 2002, 99, 52–60. [Google Scholar] [CrossRef]
- Montresor, A.; Bolomini-Vittori, M.; Simon, S.I.; Rigo, A.; Vinante, F.; Laudanna, C. Comparative analysis of normal versus CLL B-lymphocytes reveals patient-specific variability in signaling mechanisms controlling LFA-1 activation by chemokines. Cancer Res. 2009, 69, 9281–9290. [Google Scholar] [CrossRef]
- Gupta, S.K.; Pillarisetti, K.; Lysko, P.G. Modulation of CXCR4 expression and SDF-1α functional activity during differentiation of human monocytes and macrophages. J. Leukoc. Biol. 1999, 66, 135–143. [Google Scholar]
- McCormick, P.J.; Segarra, M.; Gasperini, P.; Gulino, A.V.; Tosato, G. Impaired recruitment of Grk6 and beta-Arrestin 2 causes delayed internalization and desensitization of a WHIM syndrome-associated CXCR4 mutant receptor. PLoS One 2009, 4, e8102. [Google Scholar]
- Sánchez-Martín, L.; Estecha, A.; Samaniego, R.; Sánchez-Ramón, S.; Vega, M.Á.; Sánchez-Mateos, P. The chemokine CXCL12 regulates monocyte-macrophage differentiation and RUNX3 expression. Blood 2011, 117, 88–97. [Google Scholar] [CrossRef]
- Müller, A.; Homey, B.; Soto, H.; Ge, N.; Catron, D.; Buchanan, M.E.; McClanahan, T.; Murphy, E.; Yuan, W.; Wagner, S.N.; Barrera, J.L.; Mohar, A.; Verástegui, E.; Zlotnik, A. Involvement of chemokine receptors in breast cancer metastasis. Nature 2001, 410, 50–56. [Google Scholar] [CrossRef]
- Kryczek, I.; Wei, S.; Keller, E.; Liu, R.; Zou, W. Stroma-derived factor (SDF-1/CXCL12) and human tumor pathogenesis. Am. J. Physiol. Cell Physiol. 2007, 292, C987–C995. [Google Scholar] [CrossRef]
- Andreev, J.; Galisteo, M.L.; Kranenburg, O.; Logan, S.K.; Chiu, E.S.; Okigaki, M.; Cary, L.A.; Moolenaar, W.H.; Schlessinger, J. Src and Pyk2 mediate G-protein-coupled receptor activation of epidermal growth factor receptor (EGFR) but are not required for coupling to the mitogen-activated Protein (MAP) kinase signaling cascade. J. Biol. Chem. 2001, 276, 20130–20135. [Google Scholar]
- Cabioglu, N.; Summy, J.; Miller, C.; Parikh, N.U.; Sahin, A.A.; Tuzlali, S.; Pumiglia, K.; Gallick, G.E.; Price, J.E. CXCL-12/stromal cell-derived factor-1alpha transactivates HER2-neu in breast cancer cells by a novel pathway involving Src kinase activation. Cancer Res. 2005, 65, 6493–6497. [Google Scholar] [CrossRef]
- Mahtouk, K.; Hose, D.; Rème, T.; De Vos, J.; Jourdan, M.; Moreaux, J.; Fiol, G.; Raab, M.; Jourdan, E.; Grau, V.; Moos, M.; Goldschmidt, H.; Baudard, M.; Rossi, J.F.; Cremer, F.W.; Klein, B. Expression of EGF-family receptors and amphiregulin in multiple myeloma. Amphiregulin is a growth factor for myeloma cells. Oncogene 2005, 24, 3512–3524. [Google Scholar] [CrossRef]
- Lefkowitz, R.J.; Shenoy, S.K. Transduction of Receptor Signals by β-Arrestins. Science 2005, 308, 512–517. [Google Scholar] [CrossRef]
- Whalen, E.J.; Rajagopal, S.; Lefkowitz, R.J. Therapeutic potential of beta-arrestin- and G protein-biased agonists. Trends Mol. Med. 2011, 17, 126–139. [Google Scholar] [CrossRef]
- Shenoy, S.K.; Lefkowitz, R.J. β-Arrestin-mediated receptor trafficking and signal transduction. Trends Pharmacol. Sci. 2011, 32, 521–533. [Google Scholar] [CrossRef]
- Dealwis, C.; Fernandez, E.J.; Thompson, D.A.; Simon, R.J.; Siani, M.A.; Lolis, E. Crystal structure of chemically synthesized [N33A] stromal cell-derived factor 1alpha, a potent ligand for the HIV-1 "fusin" coreceptor. Proc. Natl. Acad. Sci. USA 1998, 95, 6941–6946. [Google Scholar]
- Fen, Z.; Dhadly, M.S.; Yoshizumi, M.; Hilkert, R.J.; Quertermous, T.; Eddy, R.L.; Shows, T.B.; Lee, M.E. Structural organization and chromosomal assignment of the gene encoding the human heparin-binding epidermal growth factor-like growth factor/diphtheria toxin receptor. Biochemistry 1993, 32, 7932–7938. [Google Scholar] [CrossRef]
- McCarthy, S.A.; Samuels, M.L.; Pritchard, C.A.; Abraham, J.A.; McMahon, M. Rapid induction of heparin-binding epidermal growth factor/ diphtheria toxin receptor expression by Raf and Ras oncogenes. Genes Dev. 1995, 9, 1953–1964. [Google Scholar] [CrossRef]
- Kerkhoff, E.; Rapp, U.R. Induction of cell proliferation in quiescent NIH 3T3 cells by oncogenic c-Raf-1. Mol. Cell Biol. 1997, 17, 2576–2586. [Google Scholar]
- Hsu, H.; Xiong, J.; Goeddel, D.V. The TNF receptor 1-associated protein TRADD signals cell death and NF-kappaB activation. Cell 1995, 81, 495–504. [Google Scholar] [CrossRef]
- Schüle, R.; Umesono, K.; Mangelsdorf, D.J.; Bolado, J.; Pike, J.W.; Evans, R.M. Jun-Fos and receptors for vitamins A and D recognize a common response element in the human osteocalcin gene. Cell 1990, 61, 497–504. [Google Scholar] [CrossRef]
- Kwon, E.M.; Sakamoto, K.M. The molecular mechanism of action of granulocyte-macrophage colony-stimulating factor. J. Investig. Med. 1996, 44, 442–446. [Google Scholar]
- Kaneto, H.; Miyagawa, J.; Kajimoto, Y.; Yamamoto, K.; Watada, H.; Umayahara, Y.; Hanafusa, T.; Matsuzawa, Y.; Yamasaki, Y.; Higashiyama, S.; Taniguchi, N. Expression of heparin-binding epidermal growth factor-like growth factor during pancreas development. A potential role of PDX-1 in transcriptional activation. J. Biol. Chem. 1997, 272, 29137–29143. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, M.S.; Hancock, A.L.; Harper, J.C.; Park, J.Y.; Poy, G.; Perantoni, A.O.; Cam, M.; Malik, K.; Lee, S.B. Identification of novel Wilms' tumor suppressor gene target genes implicated in kidney development. J. Biol. Chem. 2007, 282, 16278–16287. [Google Scholar] [CrossRef]
- Morrison, S.J.; Spradling, A.C. Stem cells and niches: mechanisms that promote stem cell maintenance throughout life. Cell 2008, 132, 598–611. [Google Scholar] [CrossRef]
- Wilson, A.; Trumpp, A. Bone-marrow haematopoietic-stem-cell niches. Nat. Rev. Immunol. 2006, 6, 93–106. [Google Scholar] [CrossRef]
- Raaijmakers, M.H.G.P. Niche contributions to oncogenesis: emerging concepts and implications for the hematopoietic system. Haematologica 2011, 96, 1041–1048. [Google Scholar] [CrossRef]
- Sugiyama, T.; Kohara, H.; Noda, M.; Nagasawa, T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 2006, 25, 977–988. [Google Scholar] [CrossRef]
- Adams, G.B.; Martin, R.P.; Alley, I.R.; Chabner, K.T.; Cohen, K.S.; Calvi, L.M.; Kronenberg, H.M.; Scadden, D.T. Therapeutic targeting of a stem cell niche. Nat. Biotechnol. 2007, 25, 238–243. [Google Scholar] [CrossRef]
- Kohara, H.; Omatsu, Y.; Sugiyama, T.; Noda, M.; Fujii, N.; Nagasawa, T. Development of plasmacytoid dendritic cells in bone marrow stromal cell niches requires CXCL12-CXCR4 chemokine signaling. Blood 2007, 110, 4153–4160. [Google Scholar] [CrossRef]
- Greenbaum, A.; Hsu, Y.M.; Day, R.B.; Schuettpelz, L.G.; Christopher, M.J.; Borgerding, J.N.; Nagasawa, T.; Link, D.C. CXCL12 in early mesenchymal progenitors is required for haematopoietic stem-cell maintenance. Nature 2013, 495, 227–230. [Google Scholar] [CrossRef]
- Eminli, S.; Foudi, A.; Stadtfeld, M.; Maherali, N.; Ahfeldt, T.; Mostoslavsky, G.; Hock, H.; Hochedlinger, K. Differentiation stage determines potential of hematopoietic cells for reprogramming into induced pluripotent stem cells. Nat. Genet. 2009, 41, 968–976. [Google Scholar] [CrossRef]
- Foudi, A.; Hochedlinger, K.; Van Buren, D.; Schindler, J.W.; Jaenisch, R.; Carey, V.; Hock, H. Analysis of histone 2B-GFP retention reveals slowly cycling hematopoietic stem cells. Nat. Biotechnol. 2009, 27, 84–90. [Google Scholar]
- Yamazaki, S.; Iwama, A.; Takayanagi, S.; Morita, Y.; Eto, K.; Ema, H.; Nakauchi, H. Cytokine signals modulated via lipid rafts mimic niche signals and induce hibernation in hematopoietic stem cells. EMBO J. 2006, 25, 3515–3523. [Google Scholar] [CrossRef]
- Santaguida, M.; Schepers, K.; King, B.; Sabnis, A.J.; Forsberg, E.C.; Attema, J.L.; Braun, B.S.; Passegué, E. JunB protects against myeloid malignancies by limiting hematopoietic stem cell proliferation and differentiation without affecting self-renewal. Cancer Cell 2009, 15, 341–352. [Google Scholar] [CrossRef]
- Yamazaki, S.; Iwama, A.; Takayanagi, S.; Eto, K.; Ema, H.; Nakauchi, H. TGF-beta as a candidate bone marrow niche signal to induce hematopoietic stem cell hibernation. Blood 2009, 113, 1250–1256. [Google Scholar] [CrossRef]
- Arai, F.; Hirao, A.; Ohmura, M.; Sato, H.; Matsuoka, S.; Takubo, K.; Ito, K.; Koh, G.Y.; Suda, T. Tie2/angiopoietin-1 signaling regulates hematopoietic stem cell quiescence in the bone marrow niche. Cell 2004, 118, 149–161. [Google Scholar]
- Fleming, H.E.; Janzen, V.; Lo Celso, C.; Guo, J.; Leahy, K.M.; Kronenberg, H.M.; Scadden, D.T. Wnt signaling in the niche enforces hematopoietic stem cell quiescence and is necessary to preserve self-renewal in vivo. Cell Stem Cell 2008, 2, 274–283. [Google Scholar] [CrossRef]
- Davies, C.C.; Chakraborty, A.; Cipriani, F.; Haigh, K.; Haigh, J.J.; Behrens, A. Identification of a co-activator that links growth factor signalling to c-Jun/AP-1 activation. Nat. Cell Biol. 2010, 12, 963–972. [Google Scholar] [CrossRef]
- Butler, J.M.; Nolan, D.J.; Vertes, E.L.; Varnum-Finney, B.; Kobayashi, H.; Hooper, A.T.; Seandel, M.; Shido, K.; White, I.A.; Kobayashi, M.; Witte, L.; May, C.; Shawber, C.; Kimura, Y.; Kitajewski, J.; Rosenwaks, Z.; Bernstein, I.D.; Rafii, S. Endothelial cells are essential for the selfrenewal and repopulation of Notch-dependent hematopoietic stem cells. Cell Stem Cell 2010, 6, 251–264. [Google Scholar]
- Omatsu, Y.; Sugiyama, T.; Kohara, H.; Kondoh, G.; Fujii, N.; Kohno, K.; Nagasawa, T. The essential functions of adipo-osteogenic progenitors as the hematopoietic stem and progenitor cell niche. Immunity 2010, 33, 387–399. [Google Scholar] [CrossRef]
- Jenkinson, W.E.; Bacon, A.; White, A.J.; Anderson, G.; Jenkinson, E.J. An epithelial progenitor pool regulates thymus growth. J. Immunol. 2008, 181, 6101–6108. [Google Scholar]
- Petrie, H.T.; Zúñiga-Pflücker, J.C. Zoned out: functional mapping of stromal signaling microenvironments in the thymus. Annu. Rev. Immunol. 2007, 25, 649–679. [Google Scholar] [CrossRef]
- Rode, I.; Boehm, T. Regenerative capacity of adult cortical thymic epithelial cells. Proc. Natl. Acad. Sci. USA 2012, 109, 3463–3468. [Google Scholar] [CrossRef]
- Zandstra, P.W.; Lauffenburger, D.A.; Eaves, C.J. A ligand-receptor signaling threshold model of stem cell differentiation control: a biologically conserved mechanism applicable to hematopoiesis. Blood 2000, 96, 1215–1222. [Google Scholar]
- Burger, J.A.; Kipps, T.J. CXCR4: a key receptor in the crosstalk between tumor cells and their microenvironment. Blood 2006, 107, 1761–1767. [Google Scholar] [CrossRef]
- Corcoran, K.E.; Trzaska, K.A.; Fernandes, H.; Bryan, M.; Taborga, M.; Srinivas, V.; Packman, K.; Patel, P.S.; Rameshwar, P. Mesenchymal stem cells in early entry of breast cancer into bone marrow. PLoS One 2008, 3, e2563. [Google Scholar]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Hachet-Haas, M.; Balabanian, K.; Rohmer, F.; Pons, F.; Franchet, C.; Lecat, S.; Chow, K.Y.; Dagher, R.; Gizzi, P.; Didier, B.; Lagane, B.; Kellenberger, E.; Bonnet, D.; Baleux, F.; Haiech, J.; Parmentier, M.; Frossard, N.; Arenzana-Seisdedos, F.; Hibert, M.; Galzi, J.L. Small neutralizing molecules to inhibit actions of the chemokine CXCL12. J. Biol. Chem. 2008, 283, 23189–23199. [Google Scholar] [CrossRef]
- Sison, E.A.; Brown, P. The bone marrow microenvironment and leukemia: biology and therapeutic targeting. Expert. Rev. Hematol. 2011, 4, 271–283. [Google Scholar] [CrossRef]
- Tallman, M.S.; Gilliland, D.G.; Rowe, J.M. Drug therapy for acute myeloid leukemia. Blood 2005, 106, 1154–1163. [Google Scholar] [CrossRef]
- Appelbaum, F.R.; Rosenblum, D.; Arceci, R.J.; Carroll, W.L.; Breitfeld, P.P.; Forman, S.J.; Larson, R.A.; Lee, S.J.; Murphy, S.B.; O'Brien, S.; Radich, J.; Scher, N.S.; Smith, F.O.; Stone, R.M.; Tallman, M.S. End points to establish the efficacy of new agents in the treatment of acute leukemia. Blood 2007, 109, 1810–1816. [Google Scholar] [CrossRef]
- Jabbour, E.; Kantarjian, H. Chronic myeloid leukemia: 2012 update on diagnosis, monitoring, and management. Am. J. Hematol. 2012, 87, 1037–1045. [Google Scholar] [CrossRef]
- Gaidano, G.; Foà, R.; Dalla-Favera, R. Molecular pathogenesis of chronic lymphocytic leukemia. J. Clin. Invest. 2012, 122, 3432–3438. [Google Scholar] [CrossRef]
- Nanba, D.; Higashiyama, S. Dual intracellular signaling by proteolytic cleavage of membrane-anchored heparin-binding EGF-like growth factor. Cytokine Growth Factor Rev. 2004, 15, 13–19. [Google Scholar] [CrossRef]
- Hatfield, K.J.; Reikvam, H.; Bruserud, Ø. The crosstalk between the matrix metalloprotease system and the chemokine network in acute myeloid leukemia. Curr. Med. Chem. 2010, 17, 4448–4461. [Google Scholar] [CrossRef]
- Kim, H.Y.; Oh, Y.S.; Song, I.C.; Kim, S.W.; Lee, H.J.; Yun, H.J.; Kim, S.; Jo, D.Y. Endogenous stromal cell-derived factor-1 (CXCL12) supports autonomous growth of acute myeloid leukemia cells. Leuk. Res. 2013. [Google Scholar] [CrossRef]
- Vinante, F.; Rigo, A.; Tecchio, C.; Morosato, L.; Nadali, G.; Ricetti, M.M.; Krampera, M.; Zanolin, E.; Locatelli, F.; Gallati, H.; Chilosi, M.; Pizzolo, G. Serum levels of p55 and p75 soluble TNF receptors in adult acute leukaemia at diagnosis: correlation with clinical and biological features and outcome. Br. J. Haematol. 1998, 102, 1025–1034. [Google Scholar] [CrossRef]
- Kornblau, S.M.; McCue, D.; Singh, N.; Chen, W.; Estrov, Z.; Coombes, K.R. Recurrent expression signatures of cytokines and chemokines are present and are independently prognostic in acute myelogenous leukemia and myelodysplasia. Blood 2010, 116, 4251–4261. [Google Scholar] [CrossRef]
- Bonifacio, M.; Rigo, A.; Guardalben, E.; Bergamini, C.; Cavalieri, E.; Fato, R.; Pizzolo, G.; Suzuki, H.; Vinante, F. α-bisabolol is an effective proapoptotic agent against BCR-ABL+ cells in synergism with Imatinib and Nilotinib. PLoS One 2012, 7, e46674. [Google Scholar]
- Andhavarapu, S.; Roy, V. Immunomodulatory drugs in multiple myeloma. Expert Rev. Hematol. 2013, 6, 69–82. [Google Scholar] [CrossRef]
- Kumar, A.; Loughran, T.; Alsina, M.; Durie, B.G.; Djulbegovic, B. Management of multiple myeloma: a systematic review and critical appraisal of published studies. Lancet Oncol. 2003, 4, 293–304. [Google Scholar] [CrossRef]
- Reijmers, R.M.; Spaargaren, M.; Pals, S.T. Heparan sulfate proteoglycans in the control of B cell development and the pathogenesis of multiple myeloma. FEBS J. 2013. [Google Scholar] [CrossRef]
- Klein, B.; Tarte, K.; Jourdan, M.; Mathouk, K.; Moreaux, J.; Jourdan, E.; Legouffe, E.; De Vos, J.; Rossi, J.F. Survival and proliferation factors of normal and malignant plasma cells. Int. J. Hematol. 2003, 78, 106–113. [Google Scholar] [CrossRef]
- Blotta, S.; Jakubikova, J.; Calimeri, T.; Roccaro, A.M.; Amodio, N.; Azab, A.K.; Foresta, U.; Mitsiades, C.S.; Rossi, M.; Todoerti, K.; Molica, S.; Morabito, F.; Neri, A.; Tagliaferri, P.; Tassone, P.; Anderson, K.C.; Munshi, N.C. Canonical and noncanonical Hedgehog pathway in the pathogenesis of multiple myeloma. Blood 2012, 120, 5002–5013. [Google Scholar] [CrossRef]
- Mirandola, L.; Apicella, L.; Colombo, M.; Yu, Y.; Berta, D.G.; Platonova, N.; Lazzari, E.; Lancellotti, M.; Bulfamante, G.; Cobos, E.; Chiriva-Internati, M.; Chiaramonte, R. Anti-Notch treatment prevents multiple myeloma cells localization to the bone marrow via the chemokine system CXCR4/SDF-1. Leukemia 2013. [Google Scholar] [CrossRef]
- Badr, G.; Lefevre, E.A.; Mohany, M. Thymoquinone inhibits the CXCL12-induced chemotaxis of multiple myeloma cells and increases their susceptibility to Fas-mediated apoptosis. PLoS One 2011, 6, e23741. [Google Scholar] [CrossRef]
- Udi, J.; Schüler, J.; Wider, D.; Ihorst, G.; Catusse, J.; Waldschmidt, J.; Schnerch, D.; Follo, M.; Wäsch, R.; Engelhardt, M. Potent in vitro and in vivo activity of sorafenib in multiple myeloma: induction of cell death, CD138-downregulation and inhibition of migration through actin depolymerization. Br. J. Haematol. 2013, 161, 104–116. [Google Scholar] [CrossRef]
- Mahtouk, K.; Cremer, F.W.; Rème, T.; Jourdan, M.; Baudard, M.; Moreaux, J.; Requirand, G.; Fiol, G.; De Vos, J.; Moos, M.; Quittet, P.; Goldschmidt, H.; Rossi, J.-F.; Hose, D.; Klein, B. Heparan sulphate proteoglycans are essential for the myeloma cell growth activity of EGF-family ligands in multiple myeloma. Oncogene 2006, 25, 7180–7191. [Google Scholar] [CrossRef]
- Yoon, S.O.; Zhang, X.; Lee, I.Y.; Spencer, N.; Vo, P.; Choi, Y.S. CD9 is a novel marker for plasma cell precursors in human germinal centers. Biochem. Biophys. Res. Commun. 2013, 431, 41–46. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Vinante, F.; Rigo, A. Heparin-Binding Epidermal Growth Factor-like Growth Factor/Diphtheria Toxin Receptor in Normal and Neoplastic Hematopoiesis. Toxins 2013, 5, 1180-1201. https://doi.org/10.3390/toxins5061180
Vinante F, Rigo A. Heparin-Binding Epidermal Growth Factor-like Growth Factor/Diphtheria Toxin Receptor in Normal and Neoplastic Hematopoiesis. Toxins. 2013; 5(6):1180-1201. https://doi.org/10.3390/toxins5061180
Chicago/Turabian StyleVinante, Fabrizio, and Antonella Rigo. 2013. "Heparin-Binding Epidermal Growth Factor-like Growth Factor/Diphtheria Toxin Receptor in Normal and Neoplastic Hematopoiesis" Toxins 5, no. 6: 1180-1201. https://doi.org/10.3390/toxins5061180
APA StyleVinante, F., & Rigo, A. (2013). Heparin-Binding Epidermal Growth Factor-like Growth Factor/Diphtheria Toxin Receptor in Normal and Neoplastic Hematopoiesis. Toxins, 5(6), 1180-1201. https://doi.org/10.3390/toxins5061180