The Effects of a Chactoid Scorpion Venom and Its Purified Toxins on Rat Blood Pressure and Mast Cells Histamine Release



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
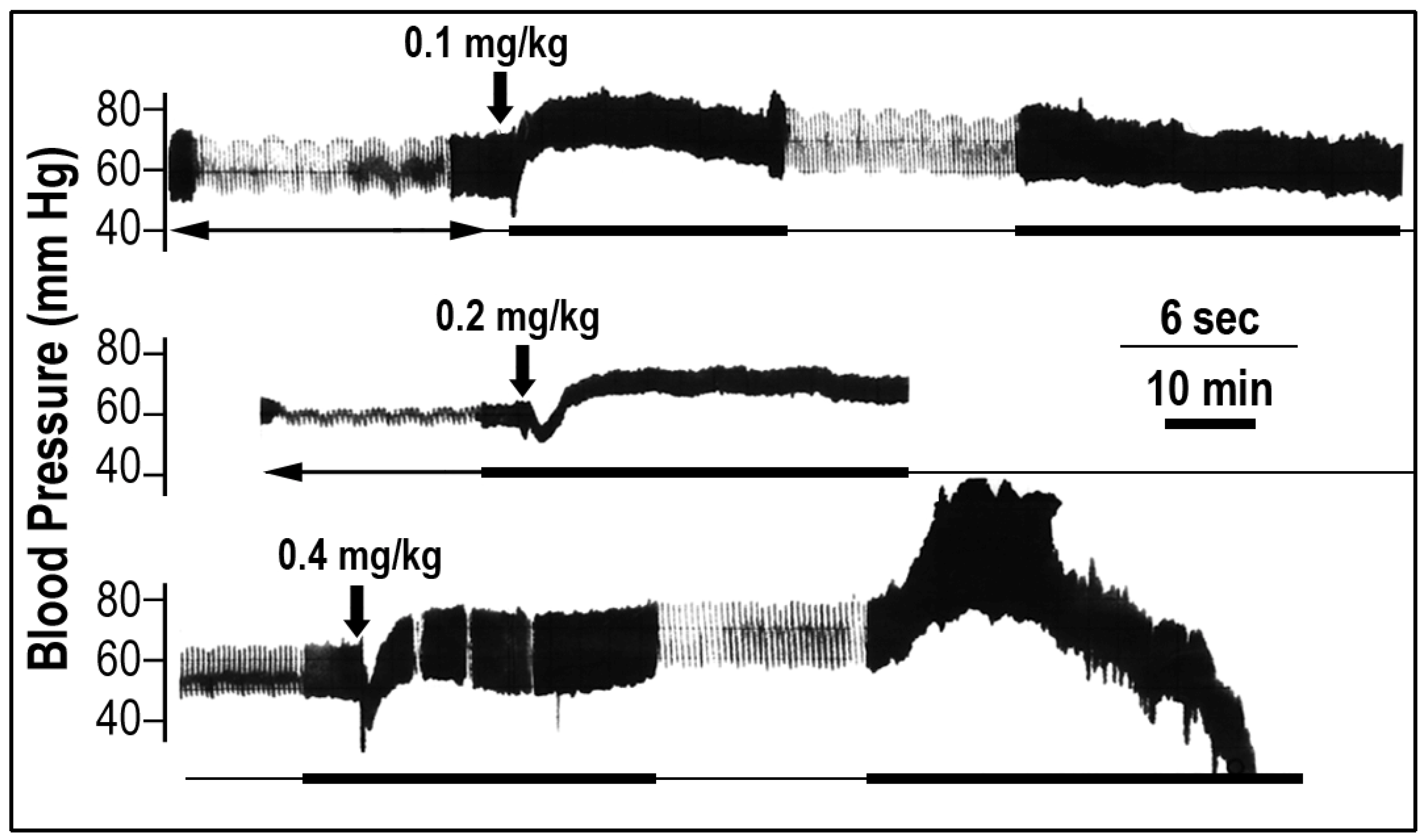
2.1. The Effect of Scorpio maurus palmatus Venom on Blood Pressure
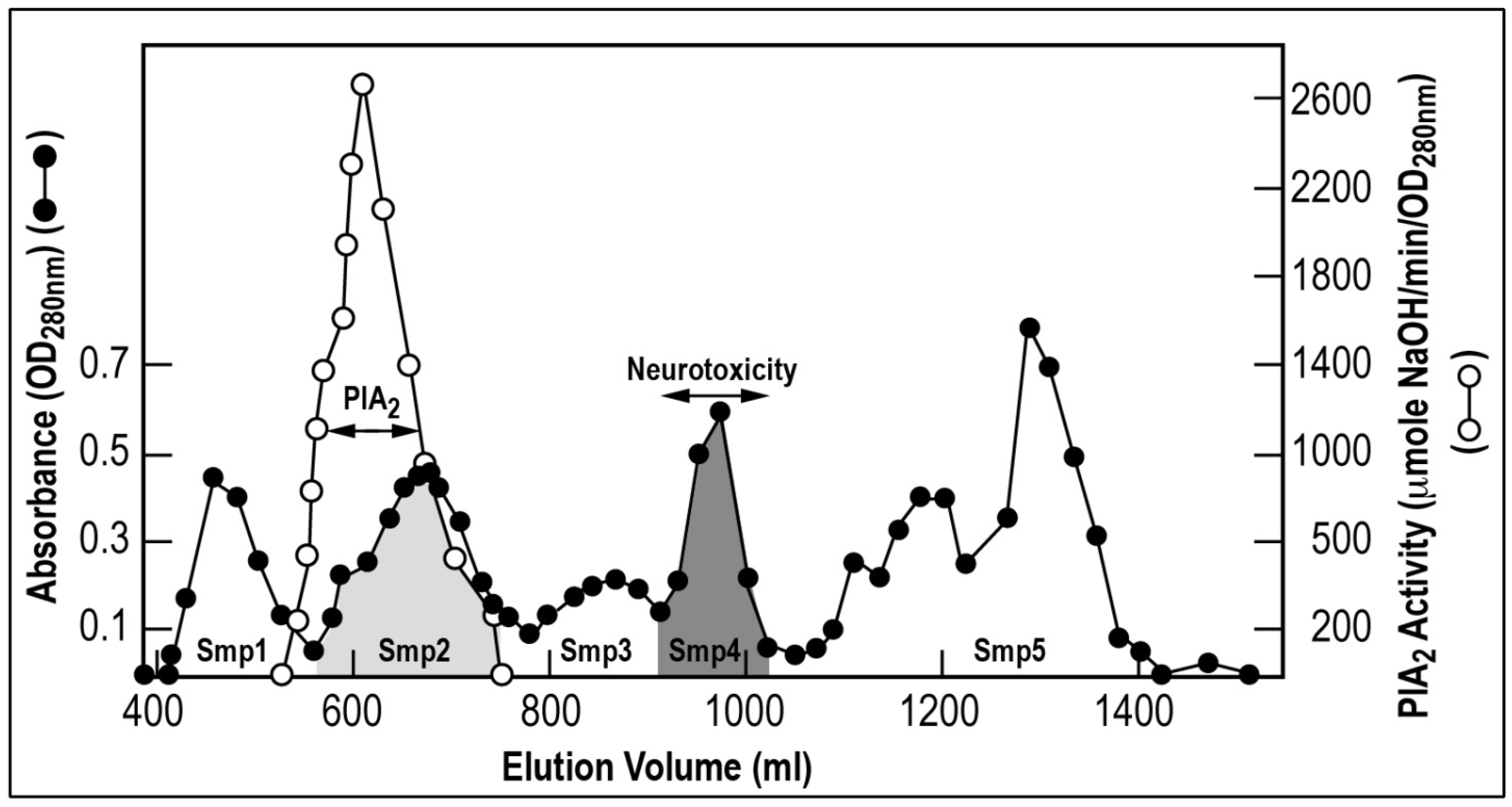
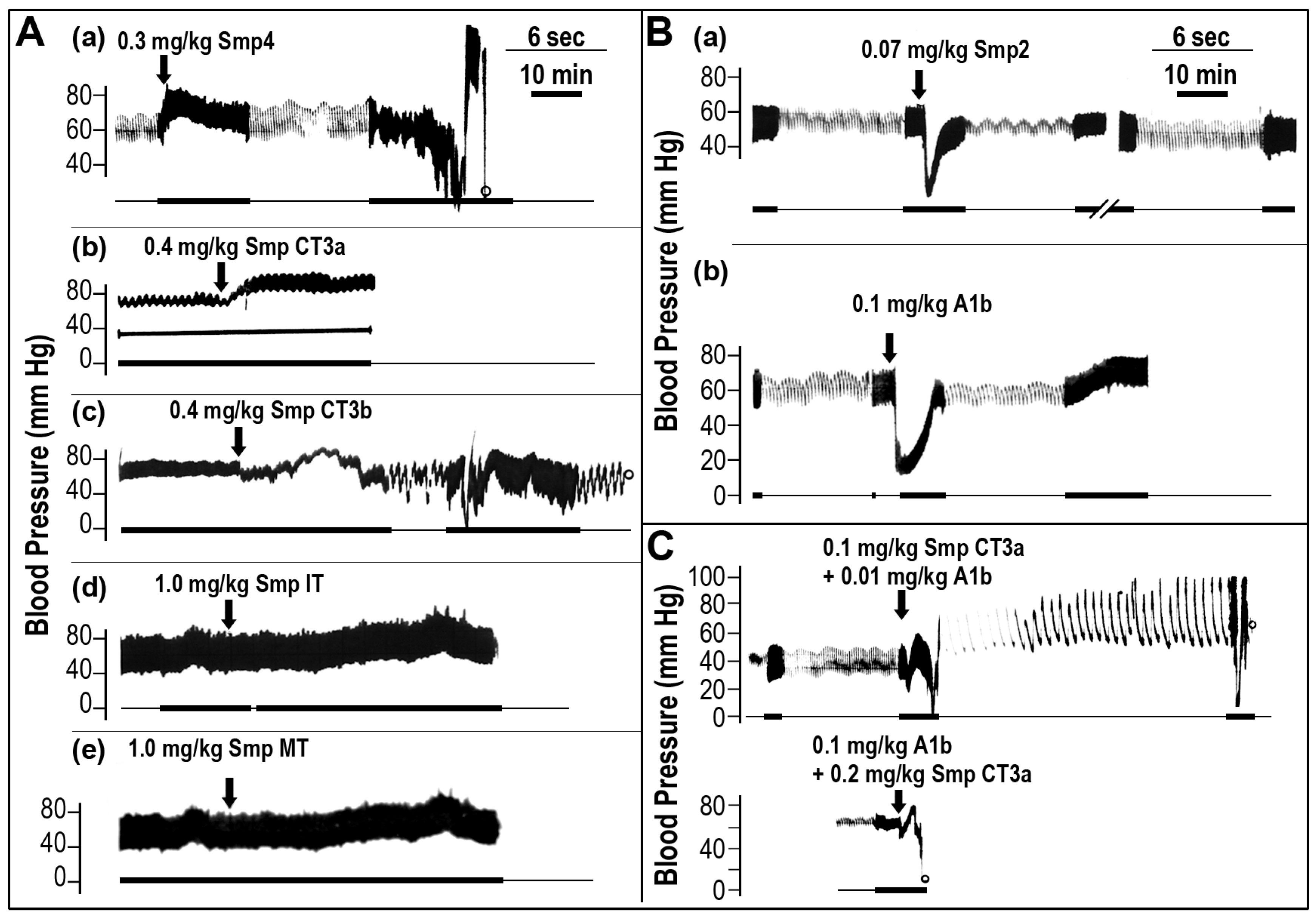
2.2. Chromatographic Separation of the Venom and Identification of Fractions Responsible for Blood Pressure Perturbation



2.3. The Effect of Venom and Isolated Protein Fractions on Histamine Release from Mast Cells

2.4. Discussion
3. Experimental Section
3.1. Materials
3.2. Venom Separation by Gel Filtration
3.3. Blood Pressure Measurement
3.4. Histamine Release Assay
3.5. Phospholipase A2 Activity Measurement
3.6. Toxicity
3.7. Statistics
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Chippaux, J.P.; Goyffon, M. Epidemiology of scorpionism: A global appraisal. Acta Trop. 2008, 107, 71–79. [Google Scholar] [CrossRef]
- Petricevich, V.L. Scorpion venom and the inflammatory response. Mediat. Inflamm. 2010, 2010, 903295. [Google Scholar] [CrossRef]
- Abdoon, N.A.; Fatani, A.J. Correlation between blood pressure, cytokines and nitric oxide in conscious rabbits injected with Leiurus quinquestriatus quinquestriatus scorpion venom. Toxicon 2009, 54, 471–480. [Google Scholar] [CrossRef]
- Hakim, A.; Sahnoun, Z.; Kallel, H.; Kassis, M.; Bouaziz, M.; Zeghal, K.M. Beneficial effects of high dobutamine doses on hemodynamic perturbations induced by Buthus occitanus tunetanus venom in rats. Med. Sci. Monit. 2009, 15, BR71–BR74. [Google Scholar]
- Nouira, S.; Elatrous, S.; Besbes, L.; Boukef, R.; Devaux, C.; Aubrey, N.; Elayeb, M.; Abroug, F. Neurohormonal activation in severe scorpion envenomation: Correlation with hemodynamics and circulating toxin. Toxicol. Appl. Pharmacol. 2005, 208, 111–116. [Google Scholar] [CrossRef]
- Tarasiuk, A.; Khvatskin, S.; Sofer, S. Effects of antivenom serotherapy on hemodynamic pathophysiology in dogs injected with L. quinquestriatus scorpion venom. Toxicon 1998, 36, 963–971. [Google Scholar] [CrossRef]
- Lazarovici, P.; Yanai, P.; Pelhate, M.; Zlotkin, E. Insect toxic component from the venom of a chactoid scorpion, Scorpio maurus palmatus (Scorpionidae). J. Biol. Chem. 1982, 257, 8397–8404. [Google Scholar]
- Lazarovici, P.; Zlotkin, E. A mammal toxin derived from the venom of a chactoid scorpion. Comp. Biochem. Physiol. C 1982, 71, 177–181. [Google Scholar] [CrossRef]
- Lazarovici, P.; Menashe, M.; Zlotkin, E. Toxicity to crustacea due to polypeptide-phospholipase interaction in the venom of a chactoid scorpion. Arch. Biochem. Biophys. 1984, 229, 270–286. [Google Scholar] [CrossRef]
- Lazarovici, P.; Menashe, M.; Primor, N.; Hochman, J.; Zlotkin, E. The cytotoxicity of a cobra venom phospholipase to mouse lymphoma cells. Arch. Toxicol. 1982, 51, 167–173. [Google Scholar] [CrossRef]
- Lee, C.Y. Mode of Action of Cobra Venom and Its Purified Toxins; Plenum Press: New York, NY, USA, 1971; Volume 1, p. 21. [Google Scholar]
- Huang, H.C. Release of slow reacting substance from the guinea-pig lung by phospholipases A2 of Vipera russelli snake venom. Toxicon 1984, 22, 359–372. [Google Scholar] [CrossRef]
- Huang, H.C.; Lee, C.Y. Isolation and pharmacological properties of phospholipases A2 from Vipera russelli (russell's viper) snake venom. Toxicon 1984, 22, 207–217. [Google Scholar] [CrossRef]
- Dutta, A.; Deshpande, S.B. Indian red scorpion venom-induced augmentation of cardio-respiratory reflexes and pulmonary edema involve the release of histamine. Toxicon 2011, 57, 193–198. [Google Scholar] [CrossRef]
- Liu, T.; Bai, Z.T.; Pang, X.Y.; Chai, Z.F.; Jiang, F.; Ji, Y.H. Degranulation of mast cells and histamine release involved in rat pain-related behaviors and edema induced by scorpion Buthus martensi karch venom. Eur. J. Pharmacol. 2007, 575, 46–56. [Google Scholar] [CrossRef]
- Galajda, Z.; Balla, J.; Szentmiklosi, A.J.; Biro, T.; Czifra, G.; Dobrosi, N.; Cseppento, A.; Patonay, L.; Roszer, T.; Balla, G.; et al. Histamine and H1-histamine receptors faster venous circulation. J. Cell. Mol. Med. 2011, 15, 2614–2623. [Google Scholar] [CrossRef]
- Roy, P.D.; Moran, D.M.; Bryant, V.; Stevenson, R.; Stanworth, D.R. Further studies on histamine release from rat mast cells in vitro induced by peptides. Characteristics of a synthetic intermediate with potent releasing activity. Biochem J. 1980, 191, 233–237. [Google Scholar]
- Gwee, M.C.; Nirthanan, S.; Khoo, H.E.; Gopalakrishnakone, P.; Kini, R.M.; Cheah, L.S. Autonomic effects of some scorpion venoms and toxins. Clin. Exp. Pharmacol. Physiol. 2002, 29, 795–801. [Google Scholar] [CrossRef]
- Vasconcelos, F.; Lanchote, V.L.; Bendhack, L.M.; Giglio, J.R.; Sampaio, S.V.; Arantes, E.C. Effects of voltage-gated Na+ channel toxins from Tityus serrulatus venom on rat arterial blood pressure and plasma catecholamines. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005, 141, 85–92. [Google Scholar] [CrossRef]
- Rodriguez, A.; Zerpa, H.; Ruiz, A.; Bermudez, V.; Garcia, F.; Silva, A.; Gutierrez, L.; Villasmil, S. Effect of clonidine in mice injected with Tityus discrepans scorpion venom. Toxicon 2013, 63, 70–77. [Google Scholar] [CrossRef]
- Bawaskar, H.S.; Bawaskar, P.H. Efficacy and safety of scorpion antivenom plus prazosin compared with prazosin alone for venomous scorpion (Mesobuthus tamulus) sting: Randomised open label clinical trial. BMJ 2011, 342, c7136. [Google Scholar] [CrossRef]
- Shore, P.A.; Burkhalter, A.; Cohn, V.H., Jr. A method for the fluorometric assay of histamine in tissues. J. Pharmacol. Exp. Ther. 1959, 127, 182–186. [Google Scholar]
- Sagi-Eisenberg, R.; Ben-Neriah, Z.; Pecht, I.; Terry, S.; Blumberg, S. Structure-activity relationship in the mast cell degranulating capacity of neurotensin fragments. Neuropharmacology 1983, 22, 197–201. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ettinger, K.; Cohen, G.; Momic, T.; Lazarovici, P. The Effects of a Chactoid Scorpion Venom and Its Purified Toxins on Rat Blood Pressure and Mast Cells Histamine Release. Toxins 2013, 5, 1332-1342. https://doi.org/10.3390/toxins5081332
Ettinger K, Cohen G, Momic T, Lazarovici P. The Effects of a Chactoid Scorpion Venom and Its Purified Toxins on Rat Blood Pressure and Mast Cells Histamine Release. Toxins. 2013; 5(8):1332-1342. https://doi.org/10.3390/toxins5081332
Chicago/Turabian StyleEttinger, Keren, Gadi Cohen, Tatjana Momic, and Philip Lazarovici. 2013. "The Effects of a Chactoid Scorpion Venom and Its Purified Toxins on Rat Blood Pressure and Mast Cells Histamine Release" Toxins 5, no. 8: 1332-1342. https://doi.org/10.3390/toxins5081332
APA StyleEttinger, K., Cohen, G., Momic, T., & Lazarovici, P. (2013). The Effects of a Chactoid Scorpion Venom and Its Purified Toxins on Rat Blood Pressure and Mast Cells Histamine Release. Toxins, 5(8), 1332-1342. https://doi.org/10.3390/toxins5081332