Effects of the Amino Acid Constituents of Microcystin Variants on Cytotoxicity to Primary Cultured Rat Hepatocytes

Abstract
:1. Introduction
2. Results





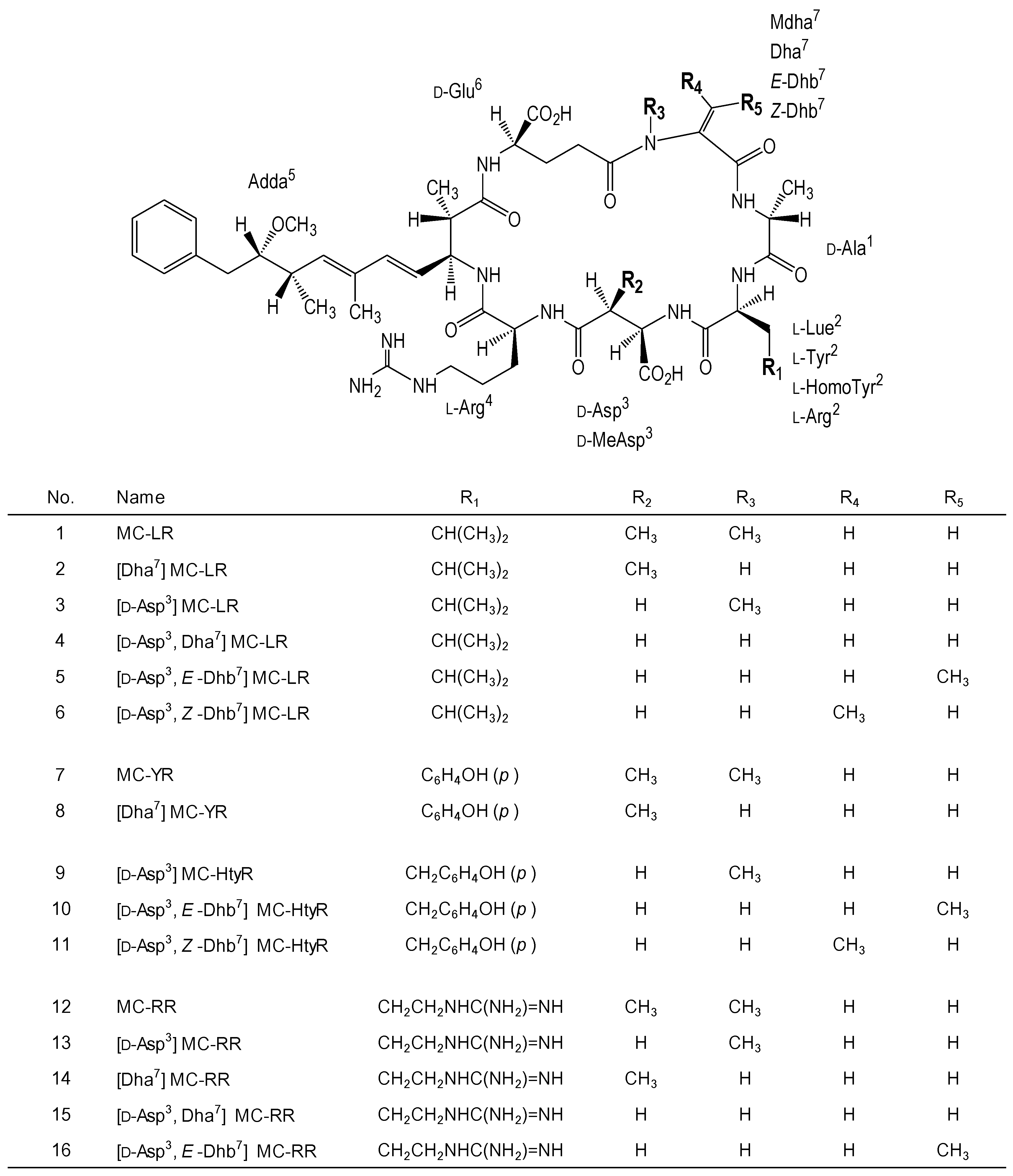{kind=link}
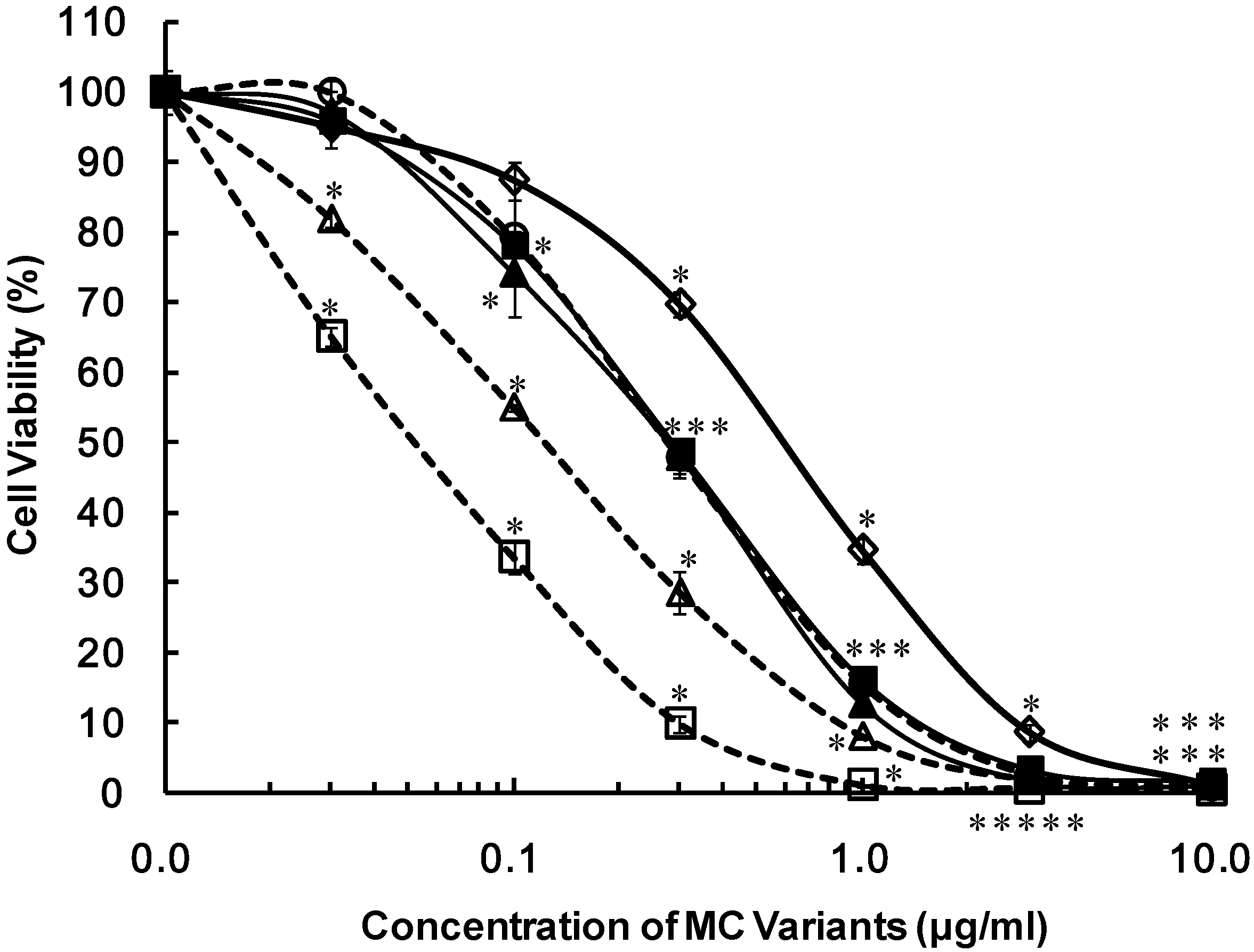{kind=link}
{kind=link}
{kind=link}
| No. | MC variants name | IC50 (μg/mL) |
|---|---|---|
| 6 | [d-Asp3, Z-Dhb7] MC-LR | 0.053 |
| 11 | [d-Asp3, Z-Dhb7] MC-HtyR | 0.120 |
| 5 | [d-Asp3, E-Dhb7] MC-LR | 0.133 |
| 4 | [d-Asp3, Dha7] MC-LR | 0.217 |
| 3 | [d-Asp3] MC-LR | 0.217 |
| 2 | [Dha7] MC-LR | 0.217 |
| 10 | [d-Asp3, E-Dhb7] MC-HtyR | 0.327 |
| 9 | [d-Asp3] MC-HtyR | 0.347 |
| 8 | [dha7] MC-YR | 0.418 |
| 1 | MC-LR | 0.800 |
| 7 | MC-YR | 1.48 |
| 15 | [d-Asp3, Dha7] MC-RR | 4.11 |
| 16 | [d-Asp3, E-Dhb7] MC-RR | 4.95 |
| 14 | [Dha7] MC-RR | 5.33 |
| 13 | [d-Asp3] MC-RR | >10 |
| 12 | MC-RR | >10 |
3. Discussion
4. Experimental Section
4.1. MC Variants
4.2. Cells
4.3. Cytotoxicity Assay
4.4. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Jochimsen, E.M.; Carmichael, W.W.; An, J.; Cardo, D.M.; Cookson, S.T.; Holmes, C.E.; Antunes, M.B.C.; de Melo Filho, D.A.; Lyra, T.M.; Barreto, V.S.T.; et al. Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. N. Engl. J. Med. 1998, 338, 873–878. [Google Scholar] [CrossRef]
- Codd, G.A.; Bell, S.G.; Kaya, K.; Ward, C.J.; Beattle, K.A.; Metcalf, J.S. Cyanobacterial toxins, exposure routes and human health. Eur. J. Physiol. 1999, 34, 405–415. [Google Scholar]
- Krishnamurthy, T.; Carmichael, W.W.; Sarver, E.W. Toxic peptides from freshwater cyanobacteria (blue-green algae). I. Isolation, purification and characterization of peptides from Microcysis aeruginosa and Anabaena flos-aquae. Toxicon 1986, 24, 865–873. [Google Scholar] [CrossRef]
- Watanabe, M.F.; Oishi, S.; Harada, K.; Matsuura, K.; Kawai, H.; Suzuki, M. Toxins contained in Microcystis species of cyanobacteria (blue-green algae). Toxicon 1988, 26, 1017–1025. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef]
- Sukenik, A.; Hadas, O.; Kaplan, A.; Quesada, A. Invasion of nostocales (cyanobacteria) to subtropical and temperate freshwater lakes—Physiological, regional, and global driving forces. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef]
- Botes, D.P.; Tuinman, A.A.; Wessels, P.L.; Viljoen, C.C.; Kruger, H.; Williams, D.H.; Santikarn, S.; Smith, R.J.; Hammond, S. The structure of cyanoginosin-LA, a cyclic heptapeptide toxin from the cyanobacterium Microcysis aeruginosa. J. Chem. Soc. Perkin Trans. 1984, 1, 2311–2318. [Google Scholar]
- Krishnamurthy, T.; Szafraniec, L.; Hunt, D.F.; Shabanowitz, J.; Yates, J.R., III; Hauer, C.R.; Carmichael, W.W.; Skulberg, O.; Codd, G.A.; Missler, S. Structural characterization of toxic cyclic peptides from blue-green algae by tandem mass spectrometry. Proc. Natl. Acad. Sci. USA 1989, 86, 770–774. [Google Scholar] [CrossRef]
- Rudolph-Böhner, S.; Mierke, D.F.; Moroder, L. Molecular structure of the cyanobacterial tumor-promoting microcystins. FEBS Lett. 1994, 349, 319–323. [Google Scholar] [CrossRef]
- Van Apeldoorn, M.E.; van Egmond, H.P.; Speijers, G.J.A.; Bakker, G.J.I. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef]
- Welker, M.; Brunke, M.; Preussel, K.; Lippert, I.; van Döhren, H. Diversity and distribution of Mycrocystis (Cyanobacteria) oligopeptide chemotypes from natural communities studied by single-colony mass spectrometry. Microbiology 2004, 150, 1785–1796. [Google Scholar] [CrossRef]
- Harada, K.; Matsuura, K.; Suzuki, M.; Watanabe, M.F.; Oishi, S.; Dahlem, A.M.; Beasley, V.R.; Carmicheal, W.W. Isolation and characterization of the minor components associated with microcystins LR and RR in the cyanobacterium (blue-green algae). Toxicon 1990, 28, 55–64. [Google Scholar] [CrossRef]
- Harada, K.; Ogawa, K.; Matsuura, K.; Murata, H.; Suzuki, M.; Watanabe, M.F.; Itezono, Y.; Nakayama, N. Structural determination of geometrical isomers of microcystins LR and RR from cyanobacteria by two-dimensional NMR spectroscopic techniques. Chem. Res. Toxicol. 1990, 3, 473–481. [Google Scholar] [CrossRef]
- An, J.; Carmichael, W.W. Use of a colorimetric protein phosphatase inhibition assay and enzyme linked immunosorbent assay for the study of microcystins and nodularins. Toxicon 1994, 32, 1495–1507. [Google Scholar] [CrossRef]
- Eriksson, J.E.; Grönberg, L.; Nygård, S.; Slotte, J.P.; Meriluoto, J.A.O. Hepatocellular uptake of 3H-dihydromicrocystin-LR, a cyclic peptide toxin. Biochim. Biophys. Acta 1990, 1025, 60–66. [Google Scholar]
- Fischer, W.J.; Altheimer, S.; Cattori, J.; Meier, P.J.; Dietrich, D.R.; Hagenbuch, B. Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin. Toxicol. Appl. Pharmacol. 2005, 203, 257–263. [Google Scholar] [CrossRef]
- Runnegar, M.T.; Kong, S.; Berndt, N. Protein phosphatase inhibition and in vivo hepatotoxicity of microcystins. Am. J. Physiol. 1993, 265, G224–G230. [Google Scholar]
- Yoshida, T.; Makita, Y.; Nagata, S.; Tsutsumi, T.; Yoshida, F.; Sekijima, M.; Tamura, S.; Ueno, Y. Acute oral toxicity of microcystin-LR, a cyanobacterial hepatotoxin, in mice. Nat. Toxins 1997, 5, 91–95. [Google Scholar] [CrossRef]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Choen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef]
- Žegura, B.; Gajski, G.; Štraser, A.; Garaj-Vrhovac, V.; Filipič, M. Microcystin-LR induced DNA damage in human peripheral blood lymphocytes. Mutat. Res. 2011, 726, 116–122. [Google Scholar] [CrossRef]
- Soares, R.M.; Cagido, V.R.; Ferraro, R.B.; Meyer-Fernandes, J.R.; Rocco, P.R.; Zin, W.A.; Azevedo, S.M. Effects of microcystin-LR on mouse lungs. Toxicon 2007, 50, 330–338. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Maatouk, I.; Plessis, M.J.; Périn, F. Genotoxic potential of microcystin-LR and in vitro in primary cultured rat hepatocytes and in vivo in rat liver. Environ. Toxicol. 2005, 20, 341–347. [Google Scholar] [CrossRef]
- Šuput, D.; Zorc-Pleskovič, R.; Petrovič, D.; Milutinovič, A. Cardiotoxic injury caused by chronic administration of microcystin-YR. Folia Biol. 2010, 56, 14–18. [Google Scholar]
- Nobre, A.C.; Jorge, M.C.; Menezes, D.B.; Fonteles, M.C.; Monteiro, H.S. Effects of microcystin-LR in isolated perfused rat kidney. Braz. J. Med. Biol. Res. 1999, 32, 985–988. [Google Scholar] [CrossRef]
- Dias, E.; Andrade, M.; Alverca, E.; Pereria, P.; Batoréu, M.C.; Jordan, P.; Silva, M.J. Comparative study of the cytotoxic effect of microcystin-LR and purified extracts from Microcystis aeruginosa on a kidney cell line. Toxicon 2009, 53, 487–495. [Google Scholar] [CrossRef]
- Li, T.; Ying, L.; Wang, H.; Li, N.; Fu, W.; Guo, Z.; Xu, L. Microcystin-LR induces ceramide to regulate PP2A and destabilize cytoskeleton in HEK293 cells. Toxicol. Sci. 2012, 128, 147–157. [Google Scholar]
- Gaudin, J.; Le Hegarat, L.; Nesslany, F.; Marzin, D.; Fessard, V. In vivo genotoxic potential of microcystin-LR: A cyanobacterial toxin, in investigated both by the unscheduled DNA synthesis (UDS) and the comet assays after intravenous administration. Environ. Toxicol. 2008, 24, 200–209. [Google Scholar]
- Hooser, S.B.; Beasley, V.R.; Lovell, R.A.; Carmichael, W.W.; Haschek, W.M. Toxicity of microcystin LR, a cyclic heptapeptide hepatotoxin from Microcystis aeruginosa, to rats and mice. Vet. Pathol. 1989, 26, 246–252. [Google Scholar] [CrossRef]
- Hooser, S.B. Fulminant hepatocyte apoptosis in vivo following microcystin-LR administration to rats. Toxicol. Pathol. 2000, 28, 726–733. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, X.; Zhou, W.; Qiao, Q.; Liang, H.; Li, G.; Wang, J.; Cai, F. The Interactive effects of cytoskeleton disruption and mitochondria dysfunction lead to reproductive toxicity induced by microcystin-LR. PLoS One 2013, 8, e53949. [Google Scholar]
- WHO. Guidelines for Drinking-Water Quality, Volume 2, Health Criteria and Other Supporting Information, Addendum, WHO/EOS/98.1. World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Nicholson, B.C.; Burch, M.D. Evaluation of Analytical Methods for Detection and Quantification of Cyanotoxins in Relation to Australian Drinking Water Guidelines. The National Health and Medical Research Council of Australia; The Water Services Association of Australia; The Cooperative Research Centre for Water Quality and Treatment: Canberra, Australia, 2001. [Google Scholar]
- Namikoshi, M.; Sivonen, K.; Evans, W.R.; Caemichael, W.W.; Rouhiainen, L.; Luukkainen, R.; Rinehart, K.L. Structures of three new homotyrosine-containing microcystins and a new homophenylalanine variant from Anabaena sp. strain 66. Chem. Res. Toxicol. 1992, 5, 661–666. [Google Scholar] [CrossRef]
- Sivonen, K.; Namikoshi, M.; Evans, W.R.; Färdig, M.; Carmichael, W.W.; Rinehart, K.L. Three new microcystins, cyclic heptapeptides hepatotoxins, from Nostoc sp. strain 152. Chem. Res. Toxicol. 1992, 5, 464–469. [Google Scholar] [CrossRef]
- Sano, T.; Takagi, H.; Kaya, K. A Dhb-microcystin from the filamentous cyanobacterium Planktothrix rubescens. Phytochemistry 2004, 65, 2159–2162. [Google Scholar] [CrossRef]
- Wood, S.A.; Mountfort, D.; Selwood, A.I.; Holland, P.T.; Puddick, J.; Cary, S.C. Widespread distribution and identification of eight novel microcystins in Antarctic cyanobacterial mats. Appl. Environ. Microbiol. 2008, 74, 7243–7251. [Google Scholar] [CrossRef]
- Feurstein, D.; Stemmer, K.; Kleinteich, J.; Speicher, T.; Dietrich, D.R. Microcystin congener- and concentration-dependent induction of murine neuron apoptosis and neurite degeneration. Toxicol. Sci. 2011, 124, 424–431. [Google Scholar] [CrossRef]
- Vesterkvist, P.S.; Misiorek, J.O.; Spoof, L.E.; Toivola, D.M.; Meriluoto, J.A. Comparative cellular toxicity of hydrophilic and hydrophobic microcystins on Caco-2 cells. Toxins 2012, 4, 1008–1023. [Google Scholar] [CrossRef]
- Huguet, A.; Henri, J.; Petitpas, M.; Hogeveen, K.; Fessard, V. Comparative cytotoxicity, oxidative stress, and cytokine secretion induced by two cyanotoxin variants, microcystin LR and RR, in human intestinal Caco-2 cells. J. Biochem. Mol. Toxicol. 2013, 27, 253–258. [Google Scholar] [CrossRef]
- Maatouk, I.; Bouaïcha, N.; Plessis, M.J.; Périn, F. Detection by 32P-postlabelling of 8-oxo-7,8-dihydro-2'-deoxyguanosine in DNA as biomarker of microcystin-LR and nodularin-induced DNA damage in vitro in primary cultured rat hepatocytes and in vivo in rat liver. Mutat. Res. 2004, 564, 9–20. [Google Scholar] [CrossRef]
- Espiña, B.; Louzao, M.C.; Cagide, E.; Alfonso, A.; Vieytes, M.R.; Yasumoto, T.; Botana, L.M. The methyl ester of okadaic acid is more potent than okadaic acid in disrupting the actin cytoskeleton and metabolism of primary cultured hepatocytes. Br. J. Pharmacol. 2010, 159, 337–344. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Maatouk, I. Microcystin-LR and nodularin induce intracellular glutathione alteration, reactive oxygen species production and lipid peroxidation in primary cultured rat hepatocytes. Toxicol. Lett. 2004, 148, 53–63. [Google Scholar] [CrossRef]
- Ulukaya, E.; Ozdikicioglu, F.; Yilmaztepe, O.; Demirci, M. The MTT assay yields a relatively lower result of growth inhibition than the ATP assay depending on the chemotherapeutic drugs tested. Toxicol. In Vitro 2008, 22, 232–239. [Google Scholar] [CrossRef]
- Mueller, H.; Kassack, M.U.; Wiese, M. Comparison of the usefulness of the MTT, ATT, and Calcein assay to predict the potency of cytotoxic agents in various human cancer cell lines. J. Biomol. Screen. 2004, 9, 506–515. [Google Scholar] [CrossRef]
- Blom, J.F.; Robinson, J.A.; Jüttner, F. High grazer toxicity of [d-Asp3, (E)-Dhb7] microcystin-RR of Planktothrix rubescens as compared to different microcystins. Toxicon 2001, 39, 1923–1932. [Google Scholar] [CrossRef]
- Blom, J.F.; Jüttner, F. High crustacean toxicity of microcystin congeners does not correlate with high protein phosphatase inhibitory activity. Toxicon 2005, 46, 465–470. [Google Scholar]
- Feurstein, D; Holst, K.; Fischer, A.; Dietrich, D.R. Oatp-associated uptake and toxicity of microcystins in primary murine whole brain cells. Toxicol. Appl. Pharmacol. 2009, 234, 247–255. [Google Scholar] [CrossRef]
- Sano, T.; Takagi, H.; Nishikawa, M.; Kaya, K. NIES certified reference material for microcystins, hepatotoxic cyclic peptide toxins from cyanobacterial blooms in eutrophic water bodies. Anal. Bioanal. Chem. 2008, 391, 2005–2010. [Google Scholar] [CrossRef]
- Sano, T.; Beattie, K.A.; Codd, G.A.; Kaya, K. Two (Z)-dehydrobutyrine-containing microcystins from a hepatotoxic bloom of Oscillatoria agardhii from Soulseat Loch, Scotland. J. Nat. Prod. 1998, 61, 851–853. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shimizu, K.; Sano, T.; Kubota, R.; Kobayashi, N.; Tahara, M.; Obama, T.; Sugimoto, N.; Nishimura, T.; Ikarashi, Y. Effects of the Amino Acid Constituents of Microcystin Variants on Cytotoxicity to Primary Cultured Rat Hepatocytes. Toxins 2014, 6, 168-179. https://doi.org/10.3390/toxins6010168
Shimizu K, Sano T, Kubota R, Kobayashi N, Tahara M, Obama T, Sugimoto N, Nishimura T, Ikarashi Y. Effects of the Amino Acid Constituents of Microcystin Variants on Cytotoxicity to Primary Cultured Rat Hepatocytes. Toxins. 2014; 6(1):168-179. https://doi.org/10.3390/toxins6010168
Chicago/Turabian StyleShimizu, Kumiko, Tomoharu Sano, Reiji Kubota, Norihiro Kobayashi, Maiko Tahara, Tomoko Obama, Naoki Sugimoto, Tetsuji Nishimura, and Yoshiaki Ikarashi. 2014. "Effects of the Amino Acid Constituents of Microcystin Variants on Cytotoxicity to Primary Cultured Rat Hepatocytes" Toxins 6, no. 1: 168-179. https://doi.org/10.3390/toxins6010168
APA StyleShimizu, K., Sano, T., Kubota, R., Kobayashi, N., Tahara, M., Obama, T., Sugimoto, N., Nishimura, T., & Ikarashi, Y. (2014). Effects of the Amino Acid Constituents of Microcystin Variants on Cytotoxicity to Primary Cultured Rat Hepatocytes. Toxins, 6(1), 168-179. https://doi.org/10.3390/toxins6010168