Nitrogen Limitation Promotes Accumulation and Suppresses Release of Cylindrospermopsins in Cells of Aphanizomenon Sp.

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
2.1.1. Influence of Nitrogen Supply


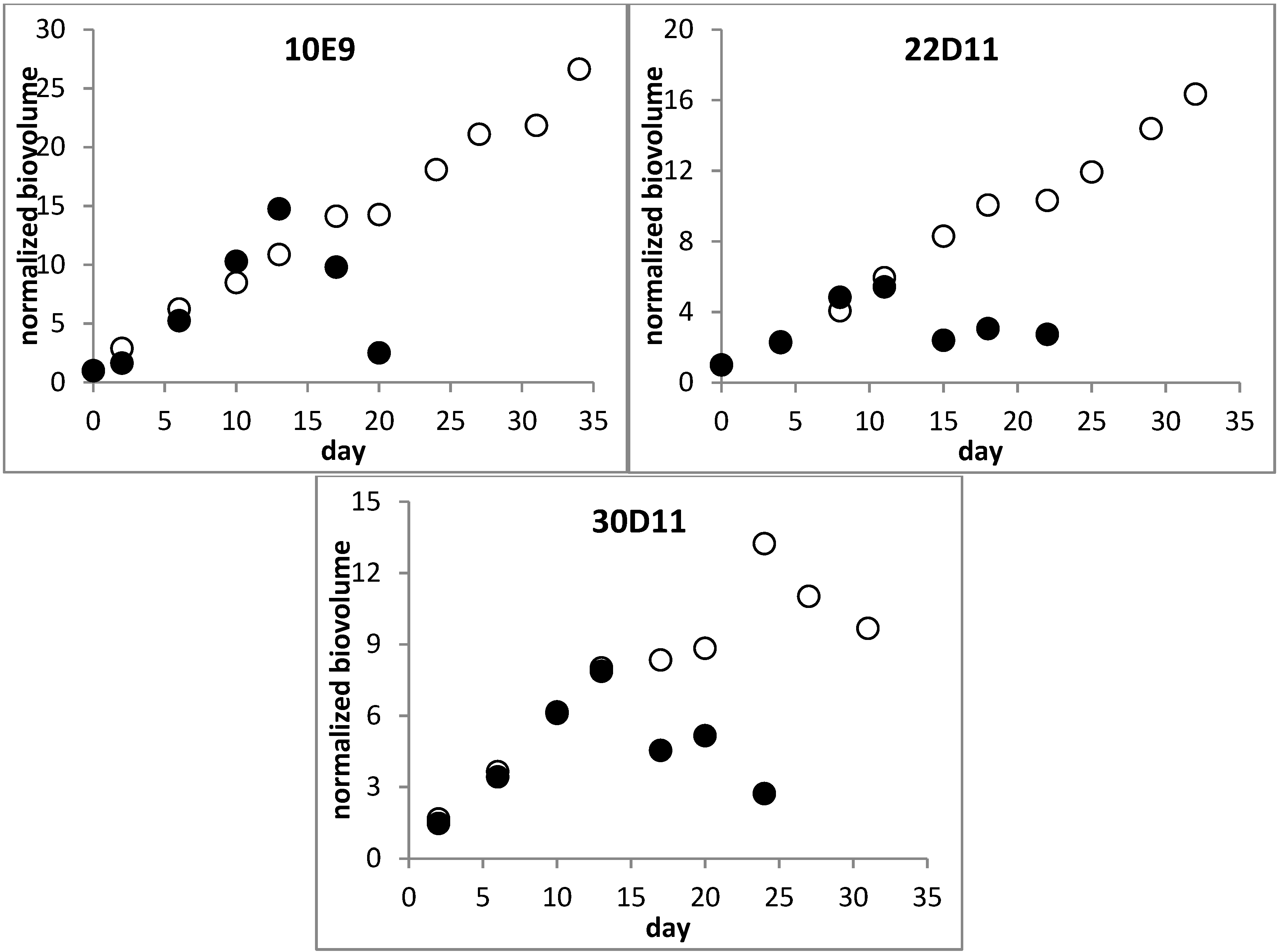{kind=link}
{kind=link}
{kind=link}
{kind=link}
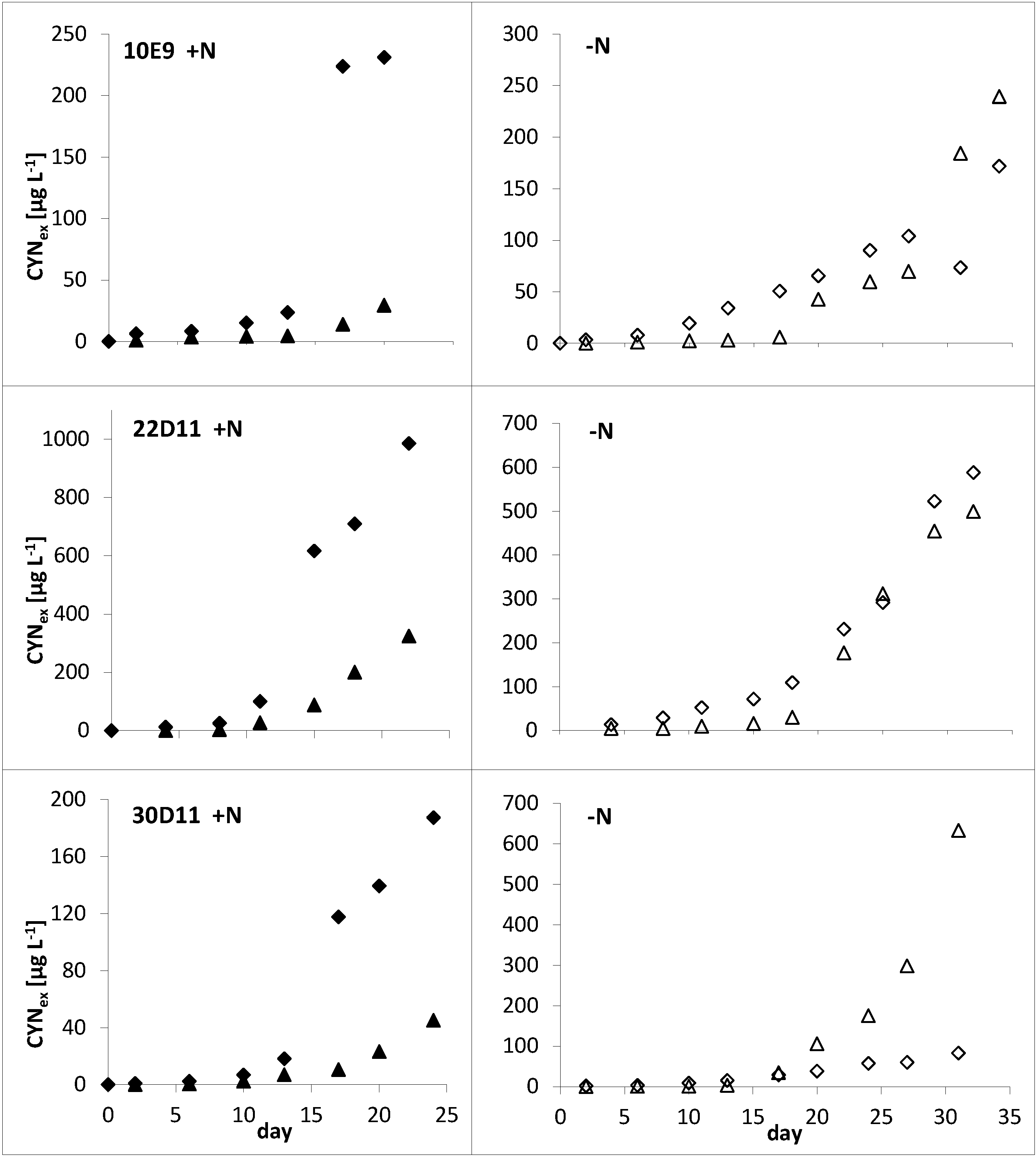{kind=link}
| Strains | 10E9 | 22D11 | 30D11 | |||
|---|---|---|---|---|---|---|
| Conitions | +N | −N | +N | −N | +N | −N |
| Biovolume (mm3 L−1) | 25.5 | 25.5 | 17.9 | 17.9 | 75.1 | 75.1 |
| CYNsin (ng mm−3) | 386.4 | 289.6 | 390.3 | 341.9 | 801.0 | 751.0 |
| CYNsex (ng mm−3) | 80.8 | 24.9 | 17.9 | 24.2 | 22.1 | 23.6 |

2.1.2. Influence of Phosphorus Supply
| Strains | Conditions | µ a (d−1) | CYNin b (ng mm−3) | D-CYNin b (ng mm−3) | CYNex c (ng mm−3) | D-CYNex c (ng mm−3) | D-CYN d (%) |
|---|---|---|---|---|---|---|---|
| 10E9 | +P | 0.229 ± 0.027 | 431 ± 54 | 36 ± 8 | 78 ± 8 | 8 ± 2 | 8 ± 2 |
| −P | 0.122 ± 0.023 | 313 ± 38 | 44 ± 11 | 286 ± 49 | 81 ± 23 | 17 ± 3 | |
| 22D11 | +P | 0.247 ± 0.032 | 339 ± 26 | 87 ± 7 | 29 ± 3 | 11 ± 1 | 21 ± 2 |
| −P | 0.133 ± 0.102 | 8 ± 2 | 4 ± 2 | 219 ± 85 | 172 ± 68 | 44 ± 1 | |
| 30D11 | +P | 0.231 ± 0.054 | 559 ± 82 | 56 ± 18 | 151 ± 135 | 31 ±33 | 10 ± 2 |
| −P | 0.125 ± 0.040 | 589 ± 114 | 105 ± 22 | 320 ± 83 | 80 ± 24 | 17 ± 2 |
2.1.3. Toxin Release


2.2. Discussion
3. Experimental Section
3.1. Cyanobacterial Strains
3.2. Experiments for Testing the Influence of Nitrogen Supply
3.3. Experiments for Testing the Influence of Phosphorus Supply
3.4. Determination of Cyanobacterial Biovolume
3.5. Toxin Analysis
3.6. Epifluorescence Microscopy and Staining to Identify Impaired Filaments
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- De la Cruz, A.A.; Hiskia, A.; Kaloudis, T.; Chernoff, N.; Hill, D.; Antoniou, M.G.; He, X.; Loftin, K.; O’Shea, K.; Zhao, C.; et al. A review on cylindrospermopsin: The global occurrence, detection, toxicity and degradation of a potent cyanotoxin. Environ. Sci. Proc. Impacts 2013, 15, 1979–2003. [Google Scholar]
- Kleinteich, J.; Hildebrand, F.; Wood, S.A.; Cirés, S.; Agha, R.; Quesada, A.; Pearce, D.A.; Convey, P.; Küpper, F.C.; Dietrich, D.R. Diversity of toxin and non-toxin containing cyanobacterial mats of meltwater ponds on the Antarctic Peninsula: A pyrosequencing approach. Antarct. Sci. 2014. [Google Scholar] [CrossRef]
- McGregor, G.B.; Fabbro, L.D. Dominance of Cylindrospermopsis raciborskii (Nostocales, Cyanoprokaryota) in Queensland tropical and subtropical reservoirs: Implications for monitoring and management. Lakes Reserv. Res. Manag. 2000, 5, 195–205. [Google Scholar] [CrossRef]
- Brient, L.; Lengronne, M.; Bormans, M.; Fastner, J. First occurrence of cylindrospermopsin in freshwater in France. Environ. Toxicol. 2009, 24, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Fastner, J.; Rücker, J.; Stüken, A.; Preußel, K.; Nixdorf, B.; Chorus, I.; Köhler, A.; Wiedner, C. Occurrence of the cyanobacterial toxin cylindrospermopsin in northeast Germany. Environ. Toxicol. 2007, 22, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Preußel, K.; Stüken, A.; Wiedner, C.; Chorus, I.; Fastner, J. First report on cylindrospermopsin producing Aphanizomenon flos-aquae (Cyanobacteria) isolated from two German lakes. Toxicon 2006, 47, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Bláhová, L.; Babica, P.; Adamovský, O.; Kohoutek, J.; Maršálek, B.; Bláha, L. Analyses of cyanobacterial toxins (microcystins, cylindrospermopsin) in the reservoirs of the Czech Republic and evaluation of health risks. Env. Chem. Lett. 2008, 6, 223–227. [Google Scholar] [CrossRef]
- Rücker, J.; Stüken, A.; Nixdorf, B.; Fastner, J.; Chorus, I.; Wiedner, C. Concentrations of particulate and dissolved cylindrospermopsin in 21 Aphanizomenon-dominated temperate lakes. Toxicon 2007, 50, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Messineo, V.; Bogialli, S.; Melchiorre, S.; Sechi, N.; Lugliè, A.; Casiddu, P.; Mariani, M.A.; Padedda, B.M.; Corcia, A.D.; Mazza, R.; et al. Cyanobacterial toxins in Italian freshwaters. Limnologica 2009, 39, 95–106. [Google Scholar]
- Elliott, J.A.; May, L. The sensitivity of phytoplankton in Loch Leven (U.K.) to changes in nutrient load and water temperature. Freshw. Biol. 2008, 53, 32–41. [Google Scholar]
- Köhler, J.; Hilt, S.; Adrian, R.; Nicklisch, A.; Kozerski, H.P.; Walz, N. Long-term response of a shallow, moderately flushed lake to reduced external phosphorus and nitrogen loading. Freshw. Biol. 2005, 50, 1639–1650. [Google Scholar] [CrossRef]
- Wiedner, C.; Rücker, J.; Brüggemann, R.; Nixdorf, B. Climate change affects timing and size of populations of an invasive cyanobacterium in temperate regions. Oecologia 2007, 152, 473–484. [Google Scholar] [CrossRef]
- Li, R.; Carmichael, W.W.; Brittain, S.; Eaglesham, G.K.; Shaw, G.R.; Liu, Y.; Watanabe, M.M. First report of the cyanotoxins cylindrospermopsin and deoxycylindrospermopsin from Raphidiopsis curvata (Cyanobacteria). J. Phycol. 2001, 37, 1121–1126. [Google Scholar] [CrossRef]
- Banker, R.; Teltsch, B.; Sukenik, A.; Carmeli, S. 7-Epicylindrospermopsin, a Toxic Minor Metabolite of the Cyanobacterium Aphanizomenon ovalisporum from Lake Kinneret, Israel. J. Nat. Prod. 2000, 63, 387–389. [Google Scholar] [CrossRef]
- Davis, T.W.; Orr, P.T.; Boyer, G.L.; Burford, M.A. Investigating the production and release of cylindrospermopsin and deoxy-cylindrospermopsin by Cylindrosermopsis raciborskii over a natural growth cycle. Harmful Algae 2014, 31, 18–25. [Google Scholar] [CrossRef]
- Chiswell, R.K.; Shaw, G.R.; Eaglesham, G.; Smith, M.J.; Norris, R.L.; Seawright, A.A.; Moore, M.R. Stability of cylindrospermopsin, the toxin from the cyanobacterium Cylindrospermopsis raciborskii: Effect of pH, temperature, and sunlight on decomposition. Environ. Toxicol. 1999, 14, 155–161. [Google Scholar] [CrossRef]
- Klitzke, S.; Apelt, S.; Weiler, C.; Fastner, J.; Chorus, I. Retention and degradation of the cyanobacterial toxin cylindrospermopsin in sediments—The role of sediment preconditioning and DOM composition. Toxicon 2010, 55, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Wörmer, L.; Cirés, S.; Carrasco, D.; Quesada, A. Cylindrospermopsin is not degraded by co-occurring natural bacterial communities during a 40-day study. Harmful Algae 2008, 7, 206–213. [Google Scholar] [CrossRef]
- Orr, P.T.; Jones, G.J. Relationship between microcystin production and cell division rates in nitrogen-limited Microcystis aeruginosa cultures. Limnol. Oceanogr. 1998, 43, 1604–1614. [Google Scholar] [CrossRef]
- Dyble, J.; Tester, P.A.; Litaker, R.W. Effects of light intensity on cylindrospermopsin production in the cyanobacterial HAB species Cylindrospermopsis raciborskii. Afr. J. Mar. Sci. 2006, 28, 309–312. [Google Scholar] [CrossRef]
- Preußel, K.; Wessel, G.; Fastner, J.; Chorus, I. Response of cylindrospermopsin production and release in Aphanizomenon flos-aquae (Cyanobacteria) to varying light and temperature conditions. Harmful Algae 2009, 8, 645–650. [Google Scholar] [CrossRef]
- Hawkins, P.R.; Putt, E.; Falconer, I.; Humpage, A. Phenotypical variation in a toxic strain of the phytoplankter, Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) during batch culture. Environ. Toxicol. 2001, 16, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Terao, K.; Ohmori, S.; Igarashi, K.; Ohtani, I.; Watanabe, M.F.; Harada, K.I.; Ito, E.; Watanabe, M. Electron microscopic studies on experimental poisoning in mice induced by cylindrospermopsin isolated from blue-green alga Umezakia natans. Toxicon 1994, 32, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, D.J.; Saker, M.L. The Palm Island mystery disease 20 years on: A review of research on the cyanotoxin cylindrospermopsin. Environ Toxicol. 2003, 18, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Lindsay, J.; Beattie, K.A.; Birmingham, S.; Saker, M.L.; Törökné, A.K.; Codd, G.A. Toxicity of cylindrospermopsin to the brine shrimp Artemia salina: Comparisons with protein synthesis inhibitors and microcystins. Toxicon 2002, 40, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Beyer, D.; Surányi, G.; Vasas, G.; Roszik, J.; Erdodi, F.; M-Hamvas, M.; Bácsi, I.; Bátori, R.; Serfozo, Z.; Szigeti, Z.M.; et al. Cylindrospermopsin induces alterations of root histology and microtubule organization in common reed (Phragmites australis) plantlets cultured in vitro. Toxicon 2009, 54, 440–449. [Google Scholar]
- Westrick, J.A.; Szlag, D.C.; Southwell, B.J.; Sinclair, J. A review of cyanobacteria and cyanotoxins removal/inactivation in drinking water treatment. Anal. Bioanal. Chem. 2010, 397, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Mazmouz, R.; Chapuis-Hugon, F.; Mann, S.; Pichon, V.; Mejean, A.; Ploux, O. Biosynthesis of cylindrospermopsin and 7-epicylindrospermopsin in Oscillatoria sp. strain PCC 6506: Identification of the cyr gene cluster and toxin analysis. Appl. Environ. Microbiol. 2010, 76, 4943–4949. [Google Scholar]
- Mihali, T.K.; Kellmann, R.; Muenchhoff, J.; Barrow, K.D.; Neilan, B.A. Characterization of the gene cluster responsible for cylindrospermopsin biosynthesis. Appl. Environ. Microbiol. 2008, 74, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Stüken, A.; Jakobsen, K.S. The cylindrospermopsin gene cluster of Aphanizomenon sp. strain 10E6: Organization and recombination. Microbiology 2010, 156, 2438–2451. [Google Scholar]
- Shalev-Malul, G.; Lieman-Hurwitz, J.; Viner-Mozzini, Y.; Sukenik, A.; Gaathon, A.; Lebendiker, M.; Kaplan, A. An AbrB-like protein might be involved in the regulation of cylindrospermopsin production by Aphanizomenon ovalisporum. Environ. Microbiol. 2008, 10, 988–999. [Google Scholar] [CrossRef] [PubMed]
- Saker, M.L.; Griffiths, D.J. The effect of temperature on growth and cylindrospermopsin content of seven isolates of Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) from water bodies in northern Australia. Phycologia 2000, 39, 349–354. [Google Scholar] [CrossRef]
- Saker, M.L.; Neilan, B.A. Varied diazotrophies, morphologies, and toxicities of genetically similar isolates of Cylindrospermopsis raciborskii (nostocales, cyanophyceae) from Northern Australia. Appl. Environ. Microbiol. 2001, 67, 1839–1845. [Google Scholar] [CrossRef] [PubMed]
- Vasas, G.; Surányi, G.; Bácsi, I.; M-Hamvas, M.; Máthé, C.; Gonda, S.; Borbely, G. Alteration of cylindrospermopsin content of Aphanizomenon ovalisporum (Cyanobacteria, Nostocales) due to step-down from combined nitrogen to dinitrogen. Adv. Microbiol. 2013, 3, 557–564. [Google Scholar] [CrossRef]
- Cirés, S.; Wörmer, L.; Ballot, A.; Agha, R.; Wiedner, C.; Velázquez, D.; Casero, M.C.; Quesada, A. Phylogeography of cylindrospermopsin and paralytic shellfish toxin-producing Nostocales cyanobacteria from Mediterranean Europe. Appl. Environ. Microbiol. 2014, 8084, 1359–1370. [Google Scholar] [CrossRef]
- Stucken Marin, K.S. Physiogenomics of Cylindrospermopsis Raciborskii and Raphidiopsis Brookii (Cyanobacteria) with Emphasis on Evolution, Nitrogen Control and Toxin Biosynthesis; University of Bremen: Bremen, Germany, 2010. [Google Scholar]
- Bar-Yosef, Y.; Sukenik, A.; Hadas, O.; Viner-Mozzini, Y.; Kaplan, A. Enslavement in the water body by toxic Aphanizomenon ovalisporum, inducing alkaline phosphatase in phytoplanktons. Curr. Biol. 2010, 20, 1557–1561. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Poniedziałek, B.; Kokociński, M.; Jurczak, T.; Lipski, D.; Wiktorowicz, K. Interspecific allelopathy in Cylindrospermopsin: Cylindrospermopsin and Cylindrospermopsis raciborskii effect on the growth and metabolism of Microcystis aeruginosa. Harmful Algae 2014, 35, 1–8. [Google Scholar]
- Wiedner, C.; Rücker, J.; Fastner, J.; Chorus, I.; Nixdorf, B. Seasonal dynamics of cylindrospermopsin and cyanobacteria in two German lakes. Toxicon 2008, 52, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Mazmouz, R.; Chapuis-Hugon, F.; Pichon, V.; Mejean, A.; Ploux, O. The last step of the biosynthesis of the cyanotoxins cylindrospermopsin and 7-epi-cylindrospermopsin is catalysed by CyrI, a 2-Oxoglutarate-dependent iron oxygenase. Chembiochem 2011, 12, 858–862. [Google Scholar] [CrossRef] [PubMed]
- McGregor, G.B.; Sendall, B.C.; Hunt, L.T.; Eaglesham, G.K. Report of the cyanotoxins cylindrospermopsin and deoxy-cylindrospermopsin from Raphidiopsis mediterranea Skuja (Cyanobacteria/Nostocales). Harmful Algae 2011, 10, 402–410. [Google Scholar] [CrossRef]
- Jiang, Y.; Xiao, P.; Yu, G.; Sano, T.; Pan, Q.; Li, R. Molecular basis and phylogenetic implications of deoxycylindrospermopsin biosynthesis in the cyanobacterium Raphidiopsis curvata. Appl. Env. Microbiol. 2012, 78, 2256–2263. [Google Scholar] [CrossRef]
- Norris, R.L.; Eaglesham, G.K.; Pierens, G.; Shaw, G.R.; Smith, M.J.; Chiswell, R.K.; Seawright, A.A.; Moore, M.R. Deoxycylindrospermopsin, an analog of cylindrospermopsin from Cylindrospermopsis raciborskii. Environ. Toxicol. 1999, 14, 163–166. [Google Scholar] [CrossRef]
- Neumann, C.; Bain, P.; Shaw, G. Studies of the comparative in vitro toxicology of the cyanobacterial metabolite deoxycylindrospermopsin. J. Toxicol. Environ. Health A 2007, 70, 1679–1686. [Google Scholar] [CrossRef] [PubMed]
- Stüken, A.; Campbell, R.J.; Quesada, A.; Sukenik, A.; Dadheech, P.K.; Wiedner, C. Genetic and morphologic characterization of four putative cylindrospermopsin producing species of the cyanobacterial genera Anabaena and Aphanizomenon. J. Plankton Res. 2009, 31, 465–480. [Google Scholar] [CrossRef]
- Staub, R. Ernährungsphysiologisch-autökologische Untersuchungen an der planktischen Blaualge Oscillatoria rubescens DC. Aquat. Sci. 1961, 23, 82–198. [Google Scholar] [CrossRef]
- Welker, M.; Bickel, H.; Fastner, J. HPLC-PDA detection of cylindrospermopsin—Opportunities and limits. Water Res. 2002, 36, 4659–4663. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Preußel, K.; Chorus, I.; Fastner, J. Nitrogen Limitation Promotes Accumulation and Suppresses Release of Cylindrospermopsins in Cells of Aphanizomenon Sp. Toxins 2014, 6, 2932-2947. https://doi.org/10.3390/toxins6102932
Preußel K, Chorus I, Fastner J. Nitrogen Limitation Promotes Accumulation and Suppresses Release of Cylindrospermopsins in Cells of Aphanizomenon Sp. Toxins. 2014; 6(10):2932-2947. https://doi.org/10.3390/toxins6102932
Chicago/Turabian StylePreußel, Karina, Ingrid Chorus, and Jutta Fastner. 2014. "Nitrogen Limitation Promotes Accumulation and Suppresses Release of Cylindrospermopsins in Cells of Aphanizomenon Sp." Toxins 6, no. 10: 2932-2947. https://doi.org/10.3390/toxins6102932
APA StylePreußel, K., Chorus, I., & Fastner, J. (2014). Nitrogen Limitation Promotes Accumulation and Suppresses Release of Cylindrospermopsins in Cells of Aphanizomenon Sp. Toxins, 6(10), 2932-2947. https://doi.org/10.3390/toxins6102932