Molecular and Insecticidal Characterization of a Novel Cry-Related Protein from Bacillus Thuringiensis Toxic against Myzus persicae



Abstract
:
1. Introduction
2. Results
2.1. Draft Genome Sequence of Strain H1.5
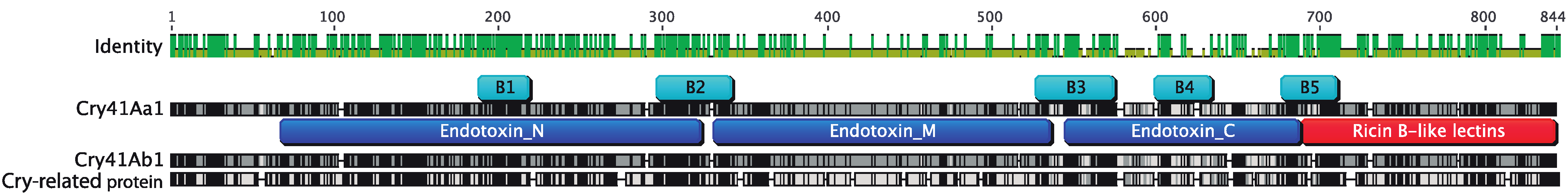
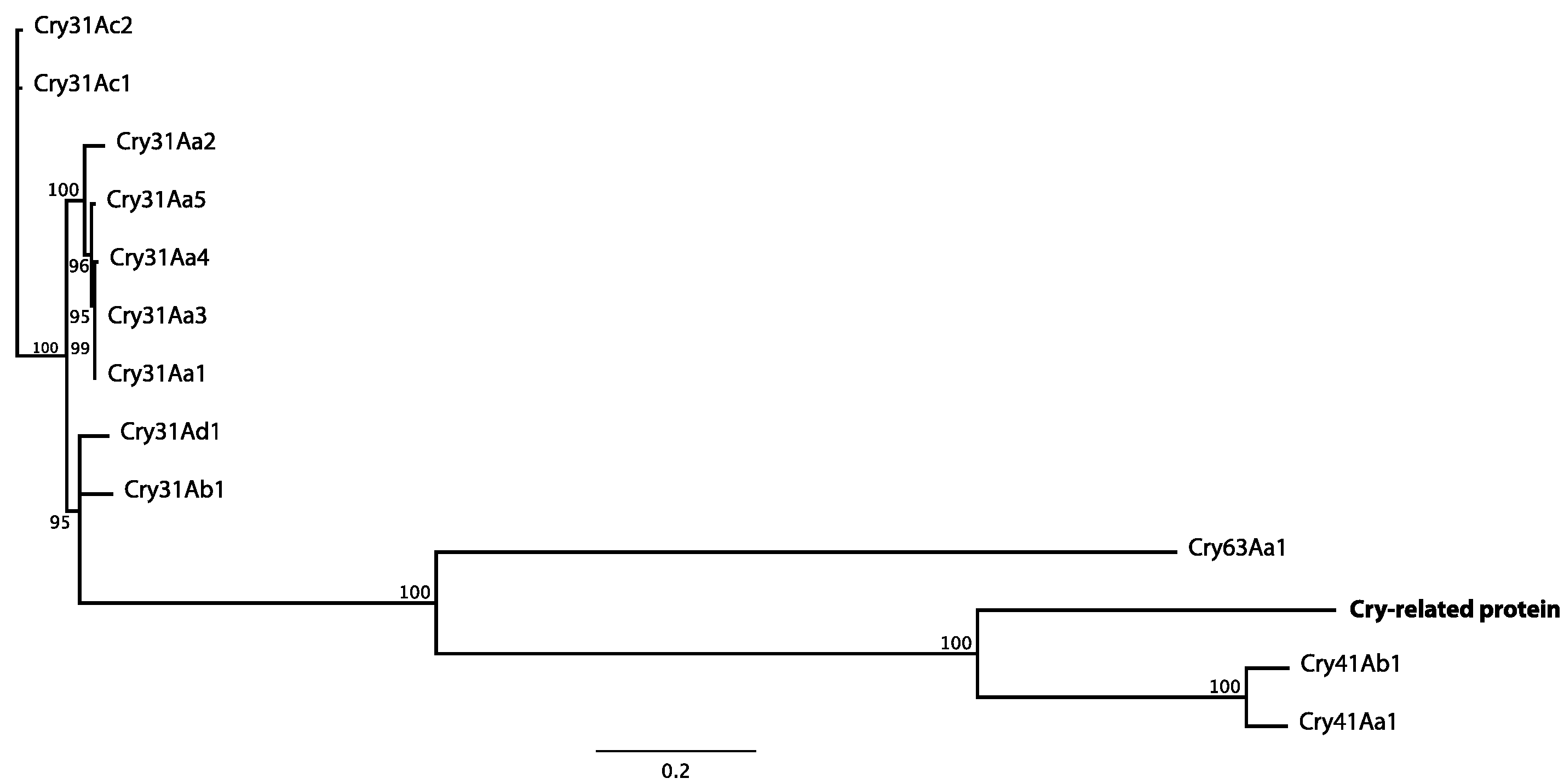
2.2. Molecular Characterization of the Novel Cry Gene


2.3. Protein Expression of the Cry-Related Gene

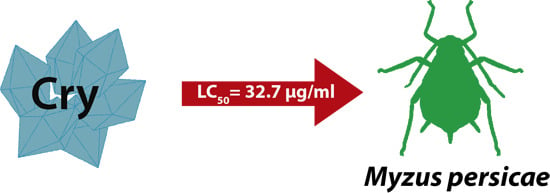
2.4. Insecticidal Activity of the Novel Cry-Related Protein

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | LC50 (µg/mL) | Regression line | Goodness of fit value | ||
|---|---|---|---|---|---|
| Slope ± SE | a * ± SE | χ2 | d.f. | ||
| Cry-related protein | 32.7 | 10.3 ± 1.4 | −10.6 ± 2.1 | 0.55 | 2 |
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Plasmids
4.2. Genome Sequencing
4.3. Computational Analysis of DNA and Protein Sequences
4.4. Amplification and Cloning of the Novel Cry-Related Gene Sequence
4.5. Protein Expression and Purification
4.6. Insect Rearing and Bioassays
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schnepf, E.; Crickmore, N.; van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar]
- Bravo, A.; Gill, S.S.; Soberón, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar]
- MacIntosh, S.C.; Stone, T.B.; Sims, S.R.; Hunst, P.L.; Greenplate, J.T.; Marrone, P.G.; Perlak, F.J.; Fischhoff, D.A.; Fuchs, R.L. Specificity and efficacy of purified Bacillus thuringiensis proteins against agronomically important insects. J. Invertebr. Pathol. 1990, 56, 258–266. [Google Scholar]
- Porcar, M.; Grenier, A.M.; Federici, B.; Rahbe, Y. Effects of Bacillus thuringiensis δ-endotoxins on the pea aphid (Acyrthosiphon pisum). Appl. Environ. Microbiol. 2009, 75, 4897–4900. [Google Scholar]
- Wei, J.Z.; Hale, K.; Carta, L.; Platzer, E.; Wong, C.; Fang, S.C.; Aroian, R.V. Bacillus thuringiensis crystal proteins that target nematodes. Proc. Natl. Acad. Sci. USA 2003, 100, 2760–2765. [Google Scholar]
- Chougule, N.P.; Bonning, B.C. Toxins for transgenic resistance to hemipteran pests. Toxins 2012, 4, 405–429. [Google Scholar]
- Shi, X.; Jiang, L.; Wang, H.; Qiao, K.; Wang, D.; Wang, K. Toxicities and sublethal effects of seven neonicotinoid insecticides on survival, growth and reproduction of imidacloprid-resistant cotton aphid, Aphis gossypii. Pest. Manag. Sci. 2011, 67, 1528–1533. [Google Scholar]
- Wang, K.Y.; Liu, T.X.; Yu, C.H.; Jiang, X.Y.; Yi, M.Q. Resistance of Aphis gossypii (Homoptera: Aphididae) to fenvalerate and imidacloprid and activities of detoxification enzymes on cotton and cucumber. J. Econ. Entomol. 2002, 95, 407–413. [Google Scholar]
- Sampson, S.K.; Dumitru, V.; Tomso, J.D. Pesticidal proteins and methods for their use. U.S. Patent 8,318,900, 27 November 2012. [Google Scholar]
- Walters, F.S.; English, L.H. Toxicity of Bacillus thuringiensis delta-endotoxins toward the potato aphid in an artificial diet bioassay. Entomol. Exp. Appl. 1995, 77, 211–216. [Google Scholar]
- Sattar, S.; Maiti, M.K. Molecular characterization of a novel vegetative insecticidal protein from Bacillus thuringiensis effective against sap-sucking insect pest. J. Microbiol. Biotechnol. 2011, 21, 937–946. [Google Scholar]
- Baum, J.A.; Sukuru, U.R.; Penn, S.R.; Meyer, S.E.; Subbarao, S.; Shi, X.; Flasinski, S.; Heck, G.R.; Brown, R.S.; Clark, T.L. Cotton plants expressing a hemipteran-active Bacillus thuringiensis crystal protein impact the development and survival of Lygus hesperus (Hemiptera: Miridae) nymphs. J. Econ. Entomol. 2012, 105, 616–624. [Google Scholar]
- Yu, Y.; Wei, Z.M. Increased oriental armyworm and aphid resistance in transgenic wheat stably expressing Bacillus thuringiensis (Bt) endotoxin and Pinellia ternate agglutinin (PTA). Plant. Cell. Tiss Org. 2008, 94, 33–44. [Google Scholar]
- Ohba, M.; Mizuki, E.; Uemori, A. Parasporin, a new anticancer protein group from Bacillus thuringiensis. Anticancer Res. 2009, 29, 427–433. [Google Scholar]
- Mizuki, E.; Park, Y.S.; Saitoh, H.; Yamashita, S.; Akao, T.; Higuchi, K.; Ohba, M. Parasporin, a human leukemic cell-recognizing parasporal protein of Bacillus thuringiensis. Clin. Diagn. Lab. Immunol. 2000, 7, 625–634. [Google Scholar]
- Yasutake, K.; Uemori, A.; Kagoshima, K.; Ohba, M. Serological identification and insect toxicity of Bacillus thuringiensis isolated from the island Okinoerabu-jima, Japan. Appl. Entomol. Zool. 2007, 42, 285–290. [Google Scholar]
- Hayakawa, T.; Kanagawa, R.; Kotani, Y.; Kimura, M.; Yamagiwa, M.; Yamane, Y.; Takebe, S.; Sakai, H. Parasporin-2Ab, a newly isolated cytotoxic crystal protein from Bacillus thuringiensis. Curr. Microbiol. 2007, 55, 278–283. [Google Scholar]
- Kondo, S.; Mizuki, E.; Akao, T.; Ohba, M. Antitrichomonal strains of Bacillus thuringiensis. Parasitol. Res. 2002, 88, 1090–1092. [Google Scholar]
- Iriarte, J.; Bel, Y.; Ferrandis, M.D.; Andrew, R.; Murillo, J.; Ferré, J.; Caballero, P. Environmental distribution and diversity of Bacillus thuringiensis in Spain. Syst. Appl. Microbiol. 1998, 21, 97–106. [Google Scholar]
- Iriarte, J.; Porcar, M.; Lecadet, M.; Caballero, P. Isolation and characterization of Bacillus thuringiensis strains from aquatic environments in Spain. Curr. Microbiol. 2000, 40, 402–408. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar]
- Ye, W.; Zhu, L.; Liu, Y.; Crickmore, N.; Peng, D.; Ruan, L.; Sun, M. Mining new crystal protein genes from Bacillus thuringiensis based on mixed plasmid-enriched genome sequencing and a computational pipeline. Appl. Environ. Microbiol. 2012, 78, 4795–4801. [Google Scholar]
- Drummond, A.J.; Ashton, B.; Buxton, S.; Cheung, M.; Cooper, A.; Duran, C.; Field, M.; Heled, J.; Kearse, M.; Markowitz, S.; et al. Geneious Pro v5.5.6. Available online: http://www.geneious.com/ (accessed on 31 October 2014).
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Pence, R.J. The antimetabolite, imidazole as a pesticide. J. Econ. Entomol. 1963, 56, 1–7. [Google Scholar]
- Ross, D.C.; Brown, T.M. Inhibition of larval growth in Spodoptera frugiperda by sublethal dietary concentrations of insecticides. J. Agric. Food Chem. 1982, 30, 193–196. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Greene, G.L.; Leppla, N.C.; Dickerson, W.A. The velvetbean caterpillar: A rearing procedure and artificial medium. J. Econ. Entomol. 1976, 69, 487–488. [Google Scholar]
- LeOra-Software POLO-PC: A User’s Guide to Probit or Logit Analysis; LeOra Software: Berkeley, CA, USA, 1987.
- Yamashita, S.; Katayama, H.; Saitoh, H.; Akao, T.; Park, Y.S.; Mizuki, E.; Ohba, M.; Ito, A. Typical three-domain Cry proteins of Bacillus thuringiensis strain A1462 exhibit cytocidal activity on limited human cancer cells. J. Biochem. 2005, 138, 663–672. [Google Scholar]
- Okumura, S.; Ohba, M.; Mizuki, E.; Crickmore, N.; Coté, J.-C.; Nagamatsu, Y.; Kitada, S.; Sakai, H.; Harata, K.; Shin, T. Parasporin nomenclature. Available online: http://parasporin.fitc.pref.fukuoka.jp/2013 (accessed on 28 October 2014).
- Burgio, G.; Lanzoni, A.; Accinelli, G.; Dinelli, G.; Bonetti, A.; Marotti, I.; Ramilli, F. Evaluation of Bt-toxin uptake by the non-target herbivore, Myzus persicae (Hemiptera: Aphididae), feeding on transgenic oilseed rape. Bull. Entomol. Res. 2007, 97, 211–215. [Google Scholar]
- Lawo, N.C.; Wäckers, F.L.; Romeis, J. Indian Bt cotton varieties do not affect the performance of cotton aphids. PLoS One 2009, 4. [Google Scholar] [CrossRef] [Green Version]
- Chakroun, M.; Bel, Y.; Caccia, S.; Abdelkefi-Mesrati, L.; Escriche, B.; Ferré, J. Susceptibility of Spodoptera frugiperda and S exigua to Bacillus thuringiensis Vip3Aa insecticidal protein. J. Invertebr. Pathol. 2012, 110, 334–339. [Google Scholar]
- Crespo, A.L.; Spencer, T.A.; Nekl, E.; Pusztai-Carey, M.; Moar, W.J.; Siegfried, B.D. Comparison and validation of methods to quantify Cry1Ab toxin from Bacillus thuringiensis for standardization of insect bioassays. Appl. Environ. Microbiol. 2008, 74, 130–135. [Google Scholar]
- Ibargutxi, M.A.; Estela, A.; Ferré, J.; Caballero, P. Use of Bacillus thuringiensis toxins for control of the cotton pest Earias insulana (Boisd.) (Lepidoptera: Noctuidae). Appl. Environ. Microbiol. 2006, 72, 437–442. [Google Scholar]
- Gatehouse, J.A. Biotechnological prospects for engineering insect-resistant plants. Plant. Physiol. 2008, 146, 881–887. [Google Scholar]
- Mehlo, L.; Gahakwa, D.; Nghia, P.T.; Loc, N.T.; Capell, T.; Gatehouse, J.A.; Gatehouse, A.M.; Christou, P. An alternative strategy for sustainable pest resistance in genetically enhanced crops. Proc. Natl. Acad. Sci. USA 2005, 102, 7812–7816. [Google Scholar]
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 37, 3–22. [Google Scholar]
- Bravo, A.; Soberón, M. How to cope with insect resistance to Bt toxins? Trends Biotechnol 2008, 26, 573–579. [Google Scholar]
- Yao, J.; Pang, Y.; Qi, H.; Wan, B.; Zhao, X.; Kong, W.; Sun, X.; Tang, K. Transgenic tobacco expressing Pinellia ternata agglutinin confers enhanced resistance to aphids. Transgenic Res. 2003, 12, 715–722. [Google Scholar]
- Monerrat, R.G.; Soares, C.M.; Capdeville, G.; Jones, G.; Soares Martins, E.; Praça, L.; Cordeiro, B.A.; Braz, S.V.; Dos Santos, R.C.; Berry, C. Translocation and insecticidal activity of Bacillus thuringiensis bacteria living inside of plants. Microbial Biotechnol. 2009, 2, 512–520. [Google Scholar]
- Melatti, V.M.; Praça, L.B.; Martins, E.S.; Sujii, E.; Berry, C.; Monnerat, R.G. Selection of Bacillus thuringiensis strains toxic against cotton aphid, Aphis gossypii Glover (Hemiptera: Aphididae). BioAssay 2010, 5, 1–4. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; De Escudero, I.R.; Caballero, P. Molecular and Insecticidal Characterization of a Novel Cry-Related Protein from Bacillus Thuringiensis Toxic against Myzus persicae. Toxins 2014, 6, 3144-3156. https://doi.org/10.3390/toxins6113144
Palma L, Muñoz D, Berry C, Murillo J, De Escudero IR, Caballero P. Molecular and Insecticidal Characterization of a Novel Cry-Related Protein from Bacillus Thuringiensis Toxic against Myzus persicae. Toxins. 2014; 6(11):3144-3156. https://doi.org/10.3390/toxins6113144
Chicago/Turabian StylePalma, Leopoldo, Delia Muñoz, Colin Berry, Jesús Murillo, Iñigo Ruiz De Escudero, and Primitivo Caballero. 2014. "Molecular and Insecticidal Characterization of a Novel Cry-Related Protein from Bacillus Thuringiensis Toxic against Myzus persicae" Toxins 6, no. 11: 3144-3156. https://doi.org/10.3390/toxins6113144
APA StylePalma, L., Muñoz, D., Berry, C., Murillo, J., De Escudero, I. R., & Caballero, P. (2014). Molecular and Insecticidal Characterization of a Novel Cry-Related Protein from Bacillus Thuringiensis Toxic against Myzus persicae. Toxins, 6(11), 3144-3156. https://doi.org/10.3390/toxins6113144