
Mycobiota and Mycotoxins in Traditional Medicinal Seeds from China

Abstract
:1. Introduction
2. Results and Discussion
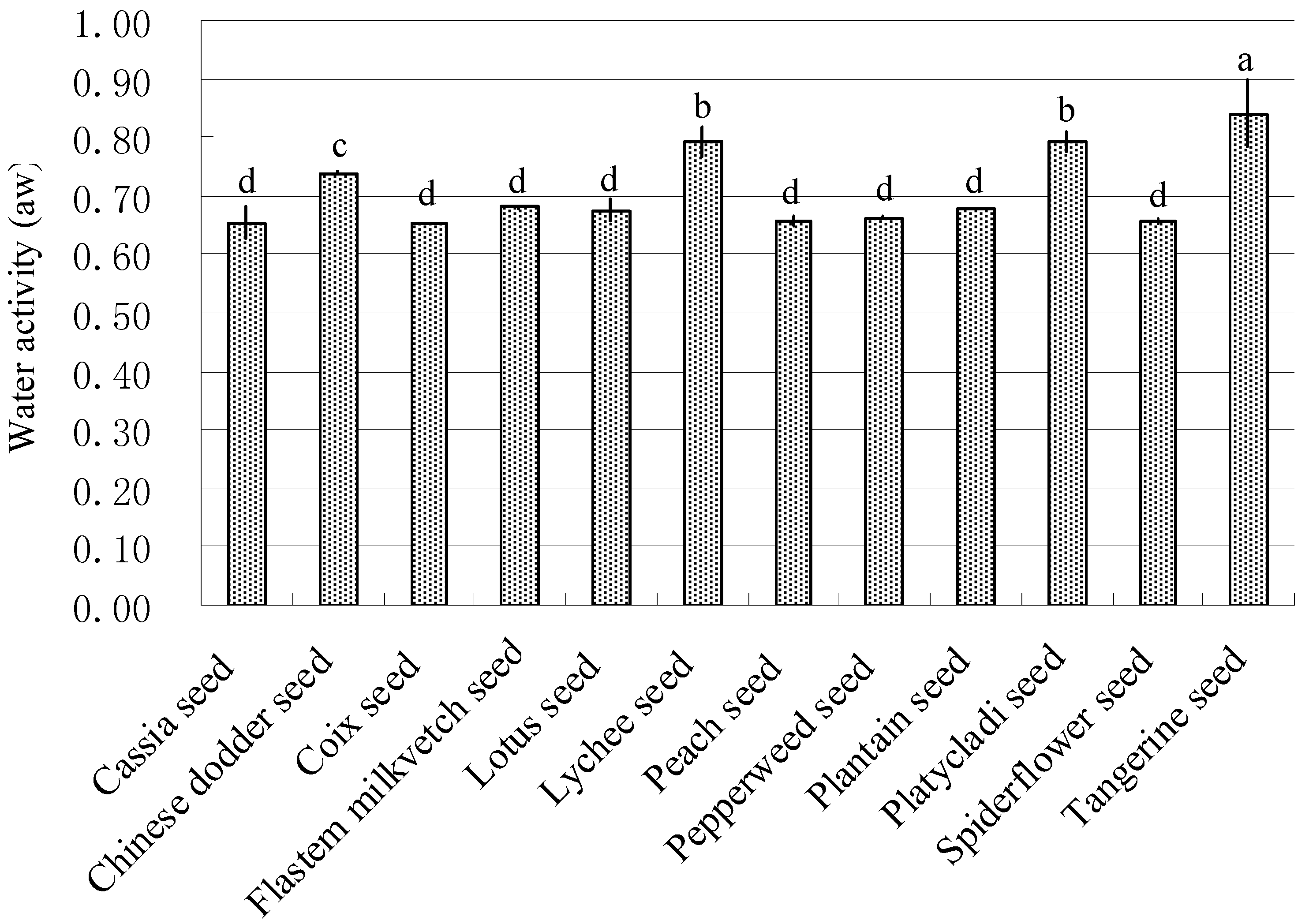
2.1. Water Activity

2.2. Internal Fungal Incidence

{kind=link}
{kind=link}
{kind=link}
| Fungal Species | Tangerine Seed | Lychee Seed | Platycladi Seed | Spiderflower Seed | Peach Seed | Coix Seed | Plantain Seed | Lotus Seed | Cassia Seed | Chinese Dodder Seed | Sum | Frequency % |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Acremonium sp. | 9 | 9 | 2.56 | |||||||||
| Alternaria alternata | 28 | 2 | 30 | 8.52 | ||||||||
| Aspergillus flavus | 25 | 1 | 26 | 7.39 | ||||||||
| Aspergillus fumigatus | 15 | 14 | 29 | 8.24 | ||||||||
| Aspergillus nidulans | 20 | 20 | 5.68 | |||||||||
| Aspergillus niger | 27 | 27 | 7.67 | |||||||||
| Aspergillus proliferans | 2 | 2 | 0.57 | |||||||||
| Aspergillus steynii | 2 | 2 | 0.57 | |||||||||
| Aspergillus tubingensis | 2 | 2 | 0.57 | |||||||||
| Aspergillus versicolor | 2 | 2 | 0.57 | |||||||||
| Botryophaeria sp. | 1 | 1 | 0.28 | |||||||||
| Chaetomium globosporum | 39 | 16 | 22 | 4 | 81 | 23.01 | ||||||
| Cochliobolus sativus | 2 | 2 | 0.57 | |||||||||
| Eurotium amstelodami | 19 | 19 | 5.4 | |||||||||
| Eurotium chevalieri | 3 | 3 | 0.85 | |||||||||
| Eurotium repens | 5 | 1 | 6 | 1.7 | ||||||||
| Eurotium rubrum | 16 | 16 | 4.55 | |||||||||
| Fusarium incarnatum | 2 | 2 | 0.57 | |||||||||
| Microascus cirrosus | 5 | 5 | 1.42 | |||||||||
| Paecilomyces variotii | 2 | 2 | 0.57 | |||||||||
| Penicillium aurantiogriseum | 2 | 2 | 0.57 | |||||||||
| Penicillium bilaiae | 6 | 6 | 1.7 | |||||||||
| Penicillium polonicum | 7 | 2 | 1 | 10 | 2.84 | |||||||
| Penicillium solitum | 3 | 3 | 0.85 | |||||||||
| Penicillium spinulosum | 2 | 2 | 0.57 | |||||||||
| Phoma sp. | 2 | 2 | 0.57 | |||||||||
| Microascus trigonosporus | 41 | 41 | 11.65 | |||||||||
| Infected seeds No. /100 seeds | 100 | 92 | 59 | 30 | 22 | 6 | 4 | 2 | 2 | 2 | ||
| Seeds infected with more than two fungal species | 20 | 9 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Co-infected fungal species | Chaetomiun globosporum + Penicillium spp. | Eurotium spp. + Aspergillus spp./ Eurotium spp. + Chaetomium globosporum | Aspergillus niger + Aspergillus flavus |
2.3. Incidence of Superficial Fungi on Seed Surfaces
| Fungal Species | Tangerine Seed | Lychee Seed | Platycladi Seed | Spiderflower Seed | Peach Seed | Coix Seed | Plantain Seed | Lotus Seed | Cassia Seed | Chinese Dodder Seed | Flastem Milkvetch Seed | Pepperweed Seed | Relative Frequency % |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Absedia sp. | 8.00 | 0.67 | |||||||||||
| Acremonium sp. | 3.03 | 0.25 | |||||||||||
| Alternaria alternata | 33.33 | 5.88 | 3.27 | ||||||||||
| Aspergillus candidus | 4.11 | 0.34 | |||||||||||
| Aspergillus flavus | 12.12 | 23.03 | 2.67 | 3.03 | 3.40 | ||||||||
| Aspergillus fumigatus | 9.84 | 5.33 | 2.74 | 40.00 | 3.70 | 5.13 | |||||||
| Aspergillus nidulans | 1.33 | 5.56 | 0.57 | ||||||||||
| Aspergillus niger | 72.47 | 22.67 | 16.00 | 28.00 | 9.09 | 12.35 | |||||||
| Aspergillus ochraceus | 31.15 | 8.00 | 3.26 | ||||||||||
| Aspergillus sydowii | 4.00 | 12.12 | 23.53 | 3.30 | |||||||||
| Aspergillus tubingensis | 56.00 | 12.12 | 5.88 | 6.17 | |||||||||
| Aspergillus versicolor | 4.00 | 4.00 | 66.67 | 5.88 | 6.71 | ||||||||
| Bispora sp. | 5.88 | 0.49 | |||||||||||
| Chaetomium globosporum | 5.56 | 3.03 | 5.88 | 1.21 | |||||||||
| Cladosporium cladosporioides | 13.89 | 12.00 | 6.06 | 2.66 | |||||||||
| Eurotium repens | 16.44 | 1.37 | |||||||||||
| Fusarium incarnatum | 50.93 | 4.24 | |||||||||||
| Fusarium solani | 11.76 | 0.98 | |||||||||||
| Mucor racemosus | 1.33 | 12.00 | 4.00 | 3.70 | 26.00 | 27.27 | 6.19 | ||||||
| Paecilomyces variotii | 64.38 | 5.37 | |||||||||||
| Penicillium aurantiogriseum | 65.33 | 5.44 | |||||||||||
| Penicillium chrysogenum | 3.28 | 3.03 | 0.53 | ||||||||||
| Penicillium citrinum | 48.48 | 4.49 | 4.00 | 3.03 | 5.88 | 5.49 | |||||||
| Penicillium commune | 1.37 | 3.70 | 0.42 | ||||||||||
| Penicillium implicatum | 5.56 | 0.46 | |||||||||||
| Penicillium oxalicum | 1.21 | 1.54 | 3.03 | 0.48 | |||||||||
| Penicillium polonicum | 38.18 | 55.74 | 1.33 | 5.48 | 20.00 | 4.00 | 18.52 | 11.11 | 15.15 | 11.76 | 15.11 | ||
| Penicillium variabile | 4.00 | 0.33 | |||||||||||
| Penicillium viridicatum | 3.70 | 0.31 | |||||||||||
| Phialophora sp. | 4.11 | 0.34 | |||||||||||
| Phoma sp. | 5.88 | 0.49 | |||||||||||
| Scopulariopsis sp. | 11.76 | 0.98 | |||||||||||
| Trichoderma koningii | 11.11 | 0.93 | |||||||||||
| Vlocladium sp. | 1.37 | 5.56 | 0.58 | ||||||||||
| Total fungal counts (CFU/g) | 4.13 × 104 | 1.53 × 102 | 4.25 × 104 | 8.1 × 104 | 1.88 × 104 | 1.83 × 102 | 6.25 × 104 | 6.5 × 101 | 6.75 × 103 | 4.5 × 102 | 8.25 × 101 | 5.75 × 104 |
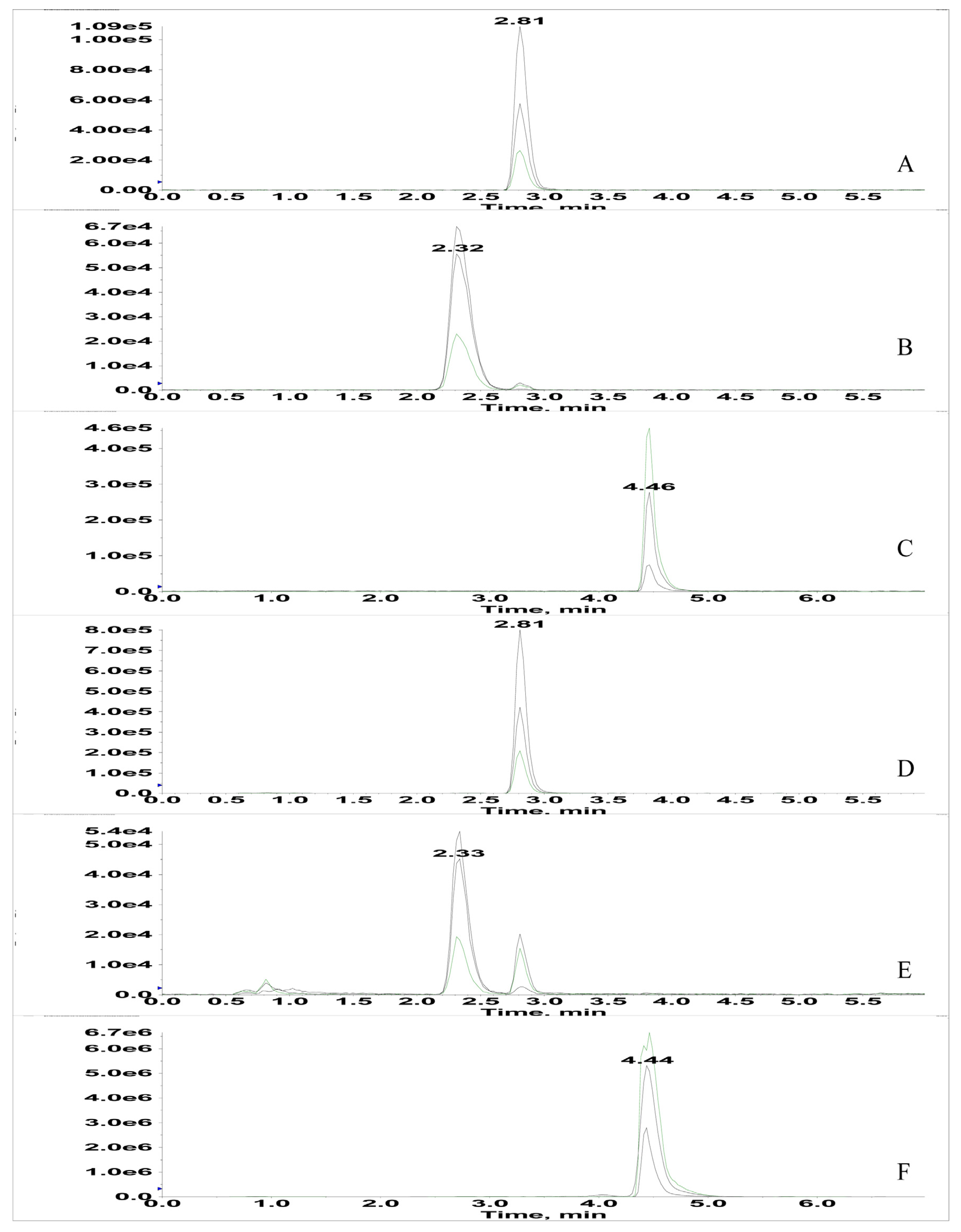
2.4. Occurrence of Mycotoxins (Aflatoxins and OTA) in Medicinal Seeds and Fungal Cultures

3. Experimental Section
3.1. Sample Collection

| No. | Name | Original Plant | Producing Regions |
|---|---|---|---|
| 1 | Cassia seed | Cassia obtusifolia L. | Hebei province |
| 2 | Chinese dodder seed | Cuscuta chinensis Lam. | Inner Mongolia autonomous region |
| 3 | Coix seed | Coix lachryma-jobi L. var. ma-yuen (Roman.) Stpf | Guizhou province |
| 4 | Flastem milkvetch seed | Astragalus complanatus R. Br. | Gansu province |
| 5 | Lotus seed | Nelumbo nucifera Gaertn. | Shandong province |
| 6 | Lychee seed | Litchi chinensis Sonn. | Guangxi autonomous region |
| 7 | Peach seed | Prunus persica (L.) Batsch | Shandong province |
| 8 | Pepperweed seed | Lepidium apetalum Willd. | Hebei province |
| 9 | Plantain seed | Plantago asiatica L. | Liaoning province |
| 10 | Platycladi seed | Platycladus orientalis (L.) Franco | Shandong province |
| 11 | Spiderflower seed | Cleome gynandra L. | Hebei province |
| 12 | Tangerine seed | Citrus reticulata Blanco | Guangdong province |
3.2. Isolation of Fungi
3.3. Identification of Fungi
3.4. Mycotoxin Production by Fungal Isolates
3.5. Analysis of Medicinal Seed Samples to Detect Natural Occurrence of AFs (AFB1, AFB2, AFG1, AFG2) and OTA
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Akerele, O. Medicinal plants in traditional medicine. In Economic and Medicinal Plant Research; Wagner, H., Farnsworth, N., Eds.; Academic Press: London, UK, 1990; Volume 4, pp. 1–174. [Google Scholar]
- MacLennan, A.H.; Wilson, D.H.; Taylor, A.W. Prevalence and cost of alternative medicine in Australia. Lancet 1996, 347, 569–573. [Google Scholar] [CrossRef]
- Barnes, P.M.; Powell-Griner, E.; McFann, K.; Nahin, R.L. Complementary and alternative medicine use among adults: United States, 2002. Semin. Integr. Med. 2004, 2, 54–71. [Google Scholar] [CrossRef]
- TCM exports set to rise at a healthy clip. Available online: http://www.chinese.cn/onlinelearning/ article/2012-03/22/content_420529.htm (accessed on 22 March 2012).
- Chinese Pharmacopoeia Council. Chinese Pharmacopoeia; Chemical Industry Press: Beijing, China, 2010; Volume 1, pp. 1–361. [Google Scholar]
- Hou, C.J.; Zhang, P.P.; Huo, D.Q. Advance in studies on the application of Semen Cassiae. Strait Pharm. J. 2007, 19, 7–9. [Google Scholar]
- Sridhar, K.R.; Bhat, R. Lotus-A potential nutraceutical source. J. Agric. Technol. 2007, 3, 143–155. [Google Scholar]
- Binder, E.M.; Tan, L.M.; Chin, L.J.; HandI, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 2007, 3–4, 265–282. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Aflatoxins: B1, B2, G1, G2, M1. In Evaluation of Carcinogenic Risks of Chemicals to Humans; IARC Monographs: Lyon, France, 1993; Volume 56, pp. 245–395. [Google Scholar]
- International Agency for Research on Cancer (IARC). Ochratoxin A. In Evaluation of Carcinogenic Risks of Chemicals to Humans; IARC Monographs: Lyon, France, 1993; Volume 56, pp. 489–521. [Google Scholar]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.E. Human ochratoxicoses. J. Toxicol. Toxin Rev. 1999, 18, 273–293. [Google Scholar] [CrossRef]
- O’Brien, E.; Dietrich, D.R. Ochratoxin A: The continuing enigma. Crit. Rev. Toxicol. 2005, 35, 33–60. [Google Scholar] [CrossRef] [PubMed]
- Abarca, M.L.; Bragulat, M.R.; Castellá, G.; Cabañes, F.J. Ochratoxin A production by strains of Aspergillus niger var. niger. Appl. Environ. Microbiol. 1994, 60, 2650–2652. [Google Scholar] [PubMed]
- Frisvad, J.C.; Frank, J.M.; Houbraken, J.A.M.P.; Kuijpers, A.F.A.; Samson, R.A. New ochratoxin A producing species of Aspergillus section Circumdati. Stud. Mycol. 2004, 50, 23–43. [Google Scholar]
- Medina, A.; Mateo, R.; López-Ocaña, L.; Valle-Algarra, F.M.; Jiménez, M. Study of Spanish grape mycobiota and ochratoxin A production by isolates of Aspergillus tubingensis and other members of Aspergillus Section. Nigri. Appl. Environ. Microbiol. 2005, 71, 4696–4702. [Google Scholar] [CrossRef] [PubMed]
- Cabañas, R.; Bragulat, M.R.; Abarca, M.L.; Castellá, G.; Cabañes, F.J. Occurrence of Penicillium verrucosum in retail wheat flours from the Spanish market. Food Microbiol. 2008, 25, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Joint FAO/WHO Expert Committee on Food Additives (JECFA). Safety Evaluation of Certain Mycotoxins in Food: WHO Food Additives Series, No. 59, 2008. Available online: http://whqlibdoc.who.int/publications/2008/9789241660594_eng.pdf (accessed on 18 November 2009).
- Logrieco, A.; Bottalico, A.; Mulé, G.; Moretti, A.; Perrone, G. Epidemiology of toxigenic fungi and their associated mycotoxins for some Mediterranean crops. Eur. J. Plant Pathol. 2003, 109, 645–667. [Google Scholar] [CrossRef]
- Chen, A.J.; Huang, L.F.; Wang, L.Z.; Tang, D.; Cai, F.; Gao, W.W. Occurrence of toxigenic fungi in ochratoxin A contaminated licorice root. Food Addit. Contam. A 2011, 28, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.J.; Tang, D.; Zhou, Y.Q.; Sun, B.D.; Li, X.J.; Wang, L.Z.; Gao, W.W. Identification of ochratoxin a producing fungi associated with fresh and dry liquorice. PLoS ONE 2013, 8, e78285. [Google Scholar] [CrossRef] [PubMed]
- Commission of the European Communities. Commission regulation (EC) No 105/2010 of 5 February 2010 setting maximum levels for certain contaminants in foodstuffs as regards ochratoxin A. Off. J. Eur. Union 2010, 35, 7–8. [Google Scholar]
- Commission of the European Communities. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 1881, 364, 5–24. [Google Scholar]
- Hao, A.Y.; Zhao, L.Y.; Liu, Y.H.; Wang, G.; Jin, H.Y.; Bi, X.L.; Men, Q.M. HPLC determination of aflatoxin residues in traditional Chinese medicine Yinpian with post column photochemical derivation and fluorescence detection. Chin. J. Pharm. Anal. 2012, 32, 2203–2207. [Google Scholar]
- Kong, W.J.; Wei, R.W.; Logrieco, A.F.; Wei, J.H.; Wen, J.; Xiao, X.H.; Yang, M.H. Occurrence of toxigenic fungi and determination of mycotoxins by HPLC-FLD in functional foods and spices in China markets. Food Chem. 2014, 146, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.H.; Chen, J.M.; Zhang, X.H. Immunoaffinity column clean-up and liquid chromatography with post-column derivatization for analysis of aflatoxins in traditional Chinese medicine. Chromatographia 2005, 62, 499–504. [Google Scholar] [CrossRef]
- Covarelli, L.; Beccari, G.; Salvi, S. Infection by mycotoxigenic fungal species and mycotoxin contamination of maize grain in Umbria, central Italy. Food Chem. Toxicol. 2011, 49, 2365–2369. [Google Scholar] [CrossRef] [PubMed]
- Goncalez, E.; Nogueira, J.H.C.; Fonseca, H.; Felicio, J.D.; Pino, F.A.; Corrêa, B. Mycobiota and mycotoxins in Brazilian peanut kernels from sowing to harvest. Int. J. Food Microbiol. 2008, 123, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Kaaya, A.N.; Kyamuhangire, W. The effect of storage time and agroecological zone on mould incidence and aflatoxin contamination of maize from traders in Uganda. Int. J. Food Microbiol. 2006, 110, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Ok, H.E.; Kim, D.M.; Kim, D.; Chung, S.H.; Chung, M.; Park, K.H.; Chun, H.S. Mycobiota and natural occurrence of aflatoxin, deoxynivalenol, nivalenol and zearalenone in rice freshly harvested in South Korea. Food Control 2014, 37, 284–291. [Google Scholar] [CrossRef]
- Sánchez-Hervás, M.; Gil, J.V.; Bisbal, F.; Ramón, D.; Martínez-Culebras, P.V. Mycobiota and mycotoxin producing fungi from cocoa beans. Int. J. Food Microbiol. 2008, 125, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Sacchi, C.; González, H.H.L.; Broggi, L.E.; Pacind, A.; Resnik, S.L.; Cano, G.; Taglieri, D. Fungal contamination and mycotoxin natural occurrence in oats for race horses feeding in Argentina. Anim. Feed Sci. Technol. 2009, 152, 330–335. [Google Scholar] [CrossRef]
- Adebajo, L.O.; Popoola, O.J. Mycoflora and mycotoxins in kolanuts during storage. Afr. J. Biotechnol. 2003, 2, 365–368. [Google Scholar]
- Atehnkeng, J.; Ojiambo, P.S.; Donner, M.; Ikotun, T.; Sikora, R.A.; Cotty, P.J.; Bandyopadhyay, R. Distribution and toxigenicity of Aspergillus species isolated from maize kernels from three agro-ecological zones in Nigeria. Int. J. Food Microbiol. 2008, 122, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Nakai, V.K.; Rocha, L.O.; Goncalez, E.; Fonseca, H.; Ortega, E.M.M.; Corrêa, B. Distribution of fungi and aflatoxins in a stored peanut variety. Food Chem. 2008, 106, 285–290. [Google Scholar] [CrossRef]
- Rodrigues, P.; Venâncio, A.; Lima, N. Mycobiota and mycotoxins of almonds and chestnuts with special reference to aflatoxins. Food Res. Int. 2012, 48, 76–90. [Google Scholar] [CrossRef]
- Li, R.H.; Feng, L.; Ma, X.Y.; Sun, Y.J.; Liu, S.P.; Jia, T.Z. Relationship between contents of fatty oil, total diterpenoids and quality of Platycladi Semen. Chin. J. Exp. Tradit. Med. Form. 2015, 21, 9–11. [Google Scholar]
- Mellon, J.E.; Cotty, P.J.; Dowd, M.K. Influence of lipids with and without other cottonseed reserve materials on aflatoxin B1 production by Aspergillus flavus. J. Agric. Food Chem. 2000, 48, 3611–3615. [Google Scholar] [CrossRef] [PubMed]
- Bayman, P.; Baker, J.L.; Mahoney, N.E. Aspergillus on tree nuts: Incidence and associations. Mycopathologia 2002, 155, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Esteban, A.; Abarca, M.L.; Bragulat, M.R.; Cabañes, F.J. Effect of water activity on ochratoxin A production by Aspergillus niger aggregate species. Int. J. Food Microbiol. 2006, 108, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Klich, M.A. Environmental and developmental factors influencing aflatoxin production by Aspergillus flavus and Aspergillus parasiticus. Mycoscience 2007, 48, 71–80. [Google Scholar] [CrossRef]
- Umikalsom, M.S.; Ariff, A.B.; Zulkifli, H.S.; Tong, C.C.; Hassan, M.A.; Karim, M.I.A. The treatment of oil palm empty fruit bunch fiber for subsequent use as substrate for cellulase production by Chaetomium globosum Kunze. Bioresource Technol. 1997, 62, 1–9. [Google Scholar] [CrossRef]
- El-Said, A.H.M.; El-Hady, G. Effect of moisture contents on the biodiversity of fungi contaminating Cuminum cyminum and Pimpinella anisum seeds under storage periods and amylolytic activity of fungal isolates. Int. J. Food Microbiol. 2014, 3, 969–991. [Google Scholar]
- Hassegawa, R.H.; Fonseca, H.; Fancelli, A.L.; da Silva, V.N.; Schammass, E.A.; Reis, T.A.; Corrêa, B. Influence of macro and micronutrient fertilization on fungal contamination and fumonisin production in corn grains. Food Control 2008, 19, 36–43. [Google Scholar] [CrossRef]
- Marin, S.; Sanchis, V.; Arnau, F.; Ramos, A.J.; Magan, N. Colonisation and competitiveness of Aspergillus and Penicillium species on maize grain in the presence of Fusarium moniliforme and Fusarium proliferatum. Int. J. Food Microbiol. 1998, 45, 107–117. [Google Scholar] [CrossRef]
- Barros, G.; Torres, A.; Chulze, S. Aspergillus flavus population isolated from soil of Argentina’s peanut-growing region. Sclerotia production and toxigenic proflle. J. Sci. Food Agric. 2005, 85, 2349–2353. [Google Scholar] [CrossRef]
- Vaamonde, G.; Patriarca, A.; Pinto, V.F.; Comeria, R.; Degrossi, C. Variability of aflatoxin and cyclopiazonic acid production by Aspergillus section Flavi from different substrates in Argentina. Int. J. Food Microbiol. 2003, 88, 79–84. [Google Scholar] [CrossRef]
- Huang, W.Y.; Cai, Y.Z.; Xing, J.; Corke, H.; Sun, M. A potential antioxidant resource: Endophytic fungi from medicinal plants. Econ. Bot. 2007, 61, 14–30. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Methuen Eyrn: London, UK, 1978; pp. 1–252. [Google Scholar]
- Carmichael, J.W.; Kendrick, W.B.; Conners, I.L.; Sigler, L. Genera of Hyphomycetes; The University of Alberta Press: Edmonton, AB, Canada, 1980; pp. 1–386. [Google Scholar]
- Klich, M.A. Identification of Common Aspergillus Species; Centraalbureau voor Schimmelculturs: Utrecht, The Netherlands, 2002; pp. 1–116. [Google Scholar]
- Pitt, J.I. The Genus Penicillium and Its Teleomorphic States Eupenicillium and Talaromyces, 1st ed.; Academic Press: Lodon, UK, 1979; pp. 1–634. [Google Scholar]
- Samson, R.A.; Frisvad, J.C. Penicillium subgenus Penicillium: New taxonomic schemes, mycotoxins and other extrolites. Stud. Mycol. 2004, 49, 1–253. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D., Sninsky, J., White, T., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–312. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, A.J.; Jiao, X.; Hu, Y.; Lu, X.; Gao, W. Mycobiota and Mycotoxins in Traditional Medicinal Seeds from China. Toxins 2015, 7, 3858-3875. https://doi.org/10.3390/toxins7103858
Chen AJ, Jiao X, Hu Y, Lu X, Gao W. Mycobiota and Mycotoxins in Traditional Medicinal Seeds from China. Toxins. 2015; 7(10):3858-3875. https://doi.org/10.3390/toxins7103858
Chicago/Turabian StyleChen, Amanda Juan, Xiaolin Jiao, Yongjian Hu, Xiaohong Lu, and Weiwei Gao. 2015. "Mycobiota and Mycotoxins in Traditional Medicinal Seeds from China" Toxins 7, no. 10: 3858-3875. https://doi.org/10.3390/toxins7103858
APA StyleChen, A. J., Jiao, X., Hu, Y., Lu, X., & Gao, W. (2015). Mycobiota and Mycotoxins in Traditional Medicinal Seeds from China. Toxins, 7(10), 3858-3875. https://doi.org/10.3390/toxins7103858