
UV-B Exposure Affects the Biosynthesis of Microcystin in Toxic Microcystis aeruginosa Cells and Its Degradation in the Extracellular Space

Abstract
:1. Introduction
2. Results
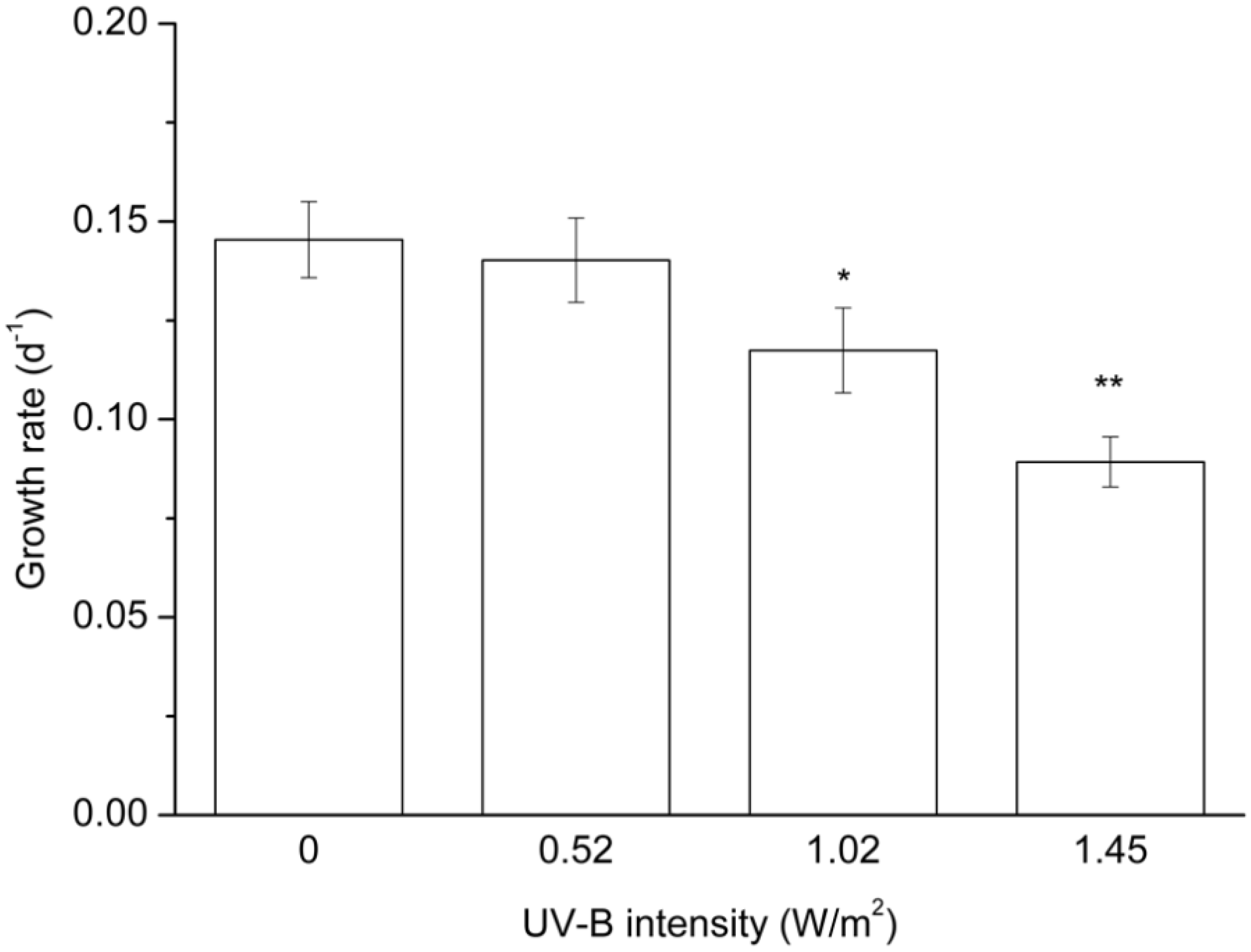
2.1. Cell Growth
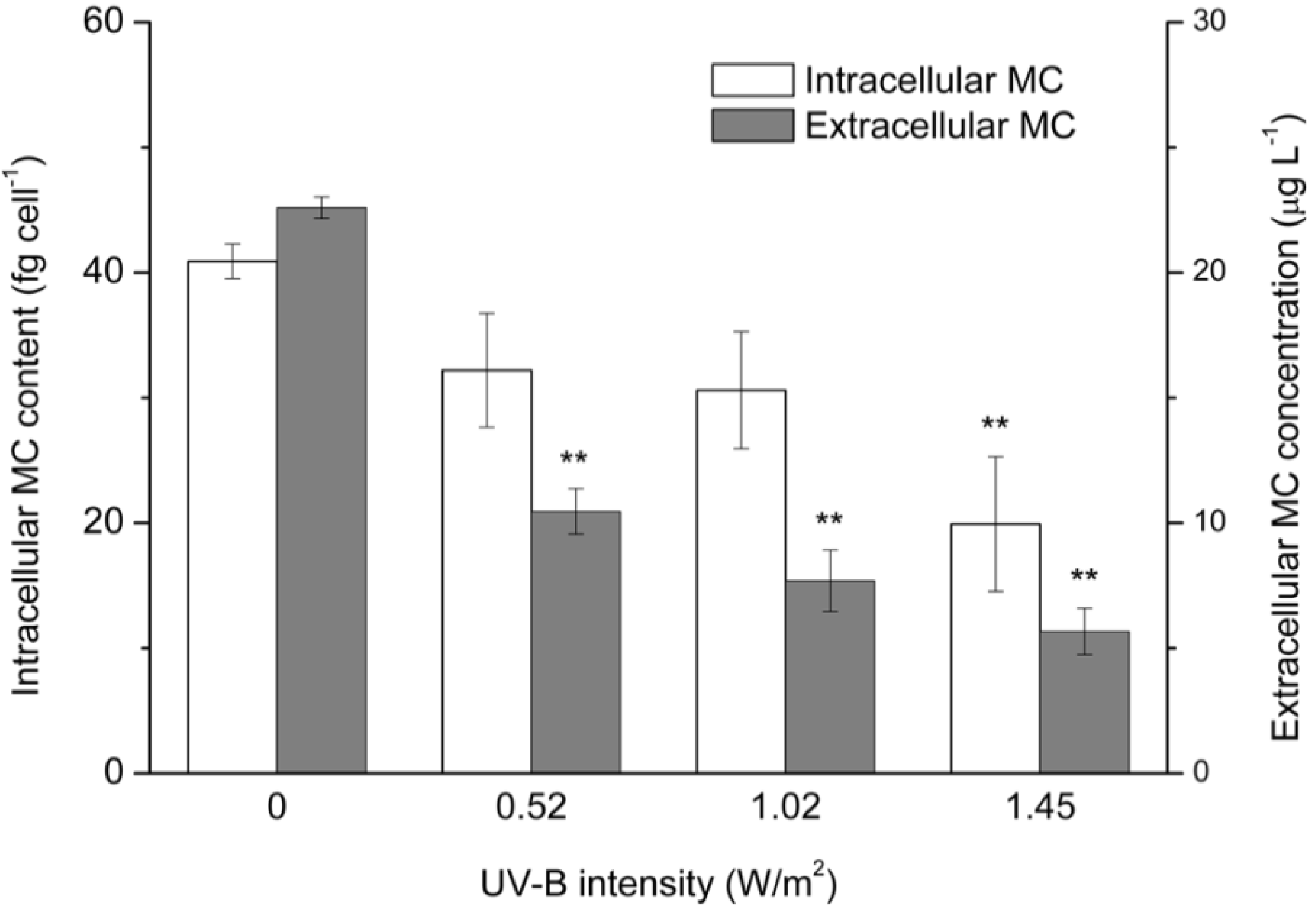
2.2. Intracellular and Extracellular MC Concentrations



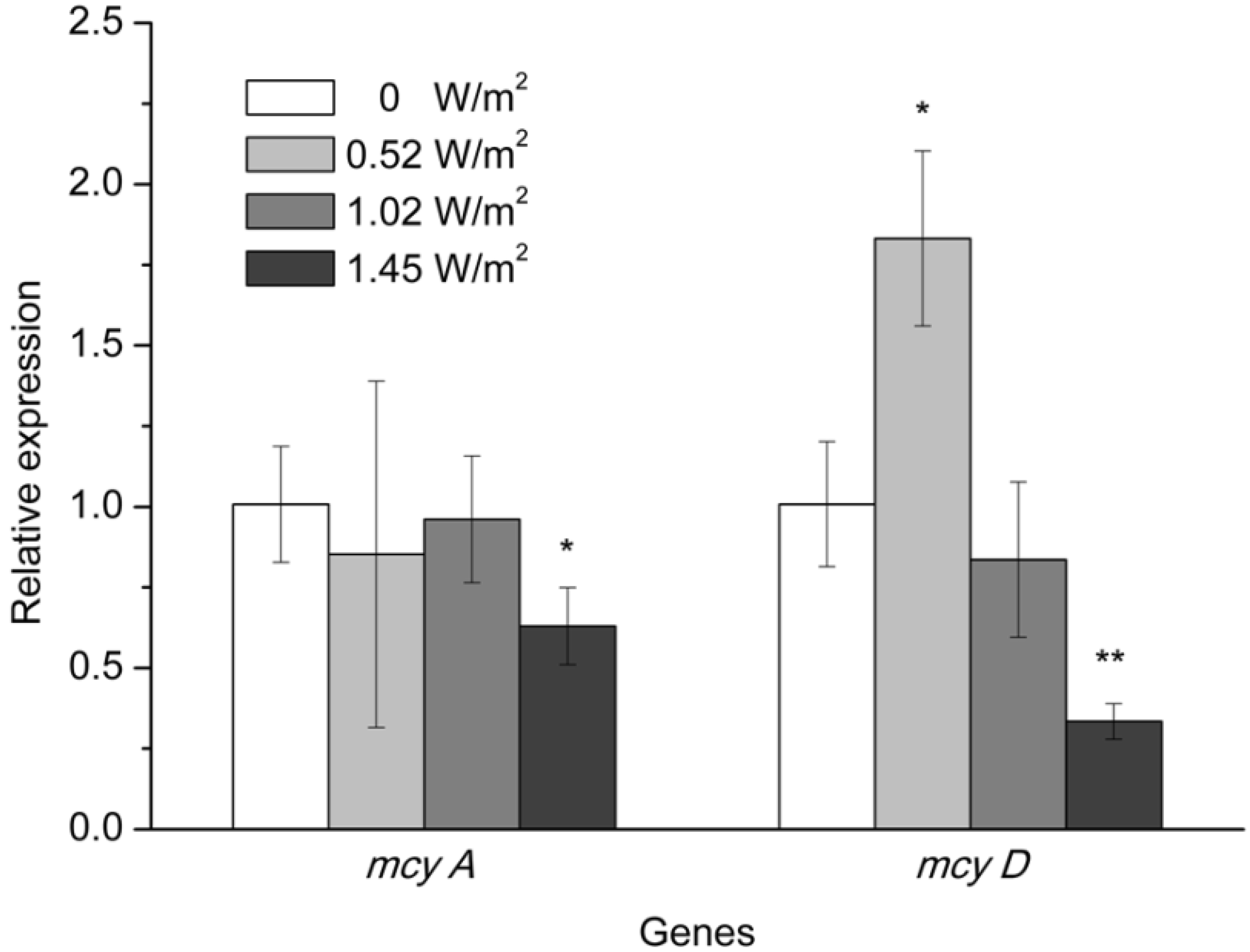
2.3. mcy Gene Expression

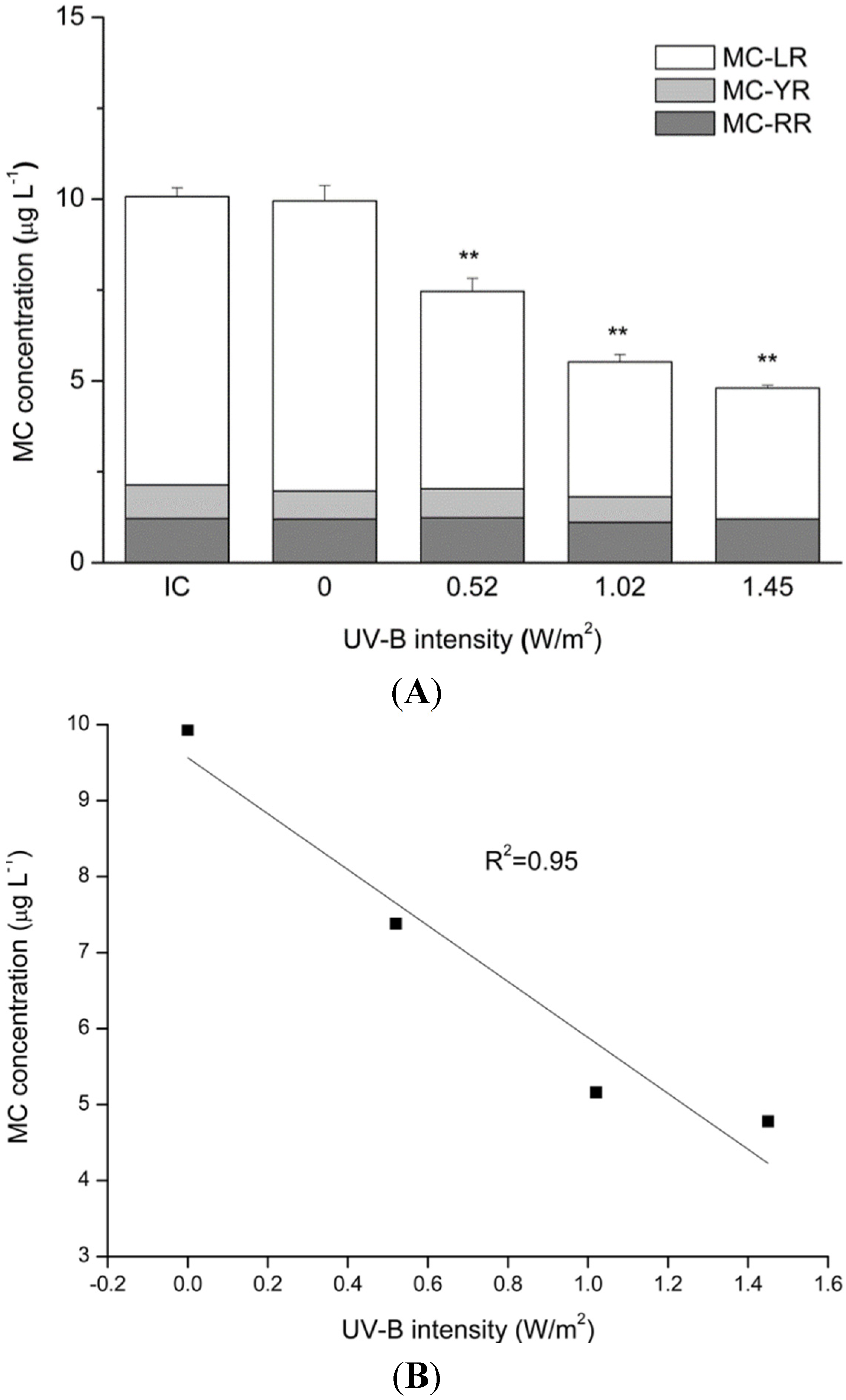
2.4. MC Degradation

3. Discussion
4. Experimental Section
4.1. Experimental Design
4.2. Gene Expression Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primers | Sequence (5′-3′) | Reference |
|---|---|---|---|
| mcyA | MSF MSF-2R | ATCCAGCAGTTGAGCAAGC GCCGATGTTTGGCTGTAAAT | [48] |
| mcyD | F2 | GGTTCGCCTGGTCAAAGTAA | [9] |
| R2 | CCTCGCTAAAGAAGGGTTGA | ||
| 16S rRNA | 184F | GCCGCRAGGTGAAAMCTAA | [49] |
| 431R | AATCCAAARACCTTCCTCCC |
4.3. MCs Extraction and Analysis
4.4. Data Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ibelings, B.W.; Backer, L.C.; Kardinaal, W.E.A.; Chorus, I. Current approaches to cyanotoxin risk assessment and risk management around the globe. Harmful Algae 2014, 40, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Sinha, R.P.; Incharoensakdi, A. The cyanotoxin-microcystins: Current overview. Rev. Environ. Sci. Biotechnol. 2014, 13, 215–249. [Google Scholar] [CrossRef]
- Srivastava, A.; Choi, G.G.; Ahn, C.Y.; Oh, H.M.; Ravi, A.K.; Asthana, R.K. Dynamics of microcystin production and quantification of potentially toxigenic Microcystis sp. using real-time PCR. Water Res. 2012, 46, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, E.; Martin-Luna, B.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. An active photosynthetic electron transfer chain required for mcyD transcription and microcystin synthesis in Microcystis aeruginosa PCC7806. Ecotoxicology 2012, 21, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Thirumavalavan, M.; Hu, Y.-L.; Lee, J.-F. Effects of humic acid and suspended soils on adsorption and photo-degradation of microcystin-LR onto samples from taiwan reservoirs and rivers. J. Hazard. Mater. 2012, 217, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Neilan, B.A.; Erhard, M.; von Dohren, H.; Borner, T. Insertional mutagenesis of a peptide synthetase gene that is responsible for hepatotoxin production in the cyanobacterium Microcystis aeruginosa PCC 7806. Mol. Microbiol. 1997, 26, 779–787. [Google Scholar] [CrossRef]
- Tillett, D.; Dittmann, E.; Erhard, M.; von Dohren, H.; Borner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC 7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef]
- Tao, M.; Xie, P.; Chen, J.; Qin, B.Q.; Zhang, D.W.; Niu, Y.; Zhang, M.; Wang, Q.; Wu, L.Y. Use of a generalized additive model to investigate key abiotic factors affecting microcystin cellular quotas in heavy bloom areas of lake Taihu. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Kaebernick, M.; Neilan, B.A.; Borner, T.; Dittmann, E. Light and the transcriptional response of the microcystin biosynthesis gene cluster. Appl. Environ. Microbiol. 2000, 66, 3387–3392. [Google Scholar] [PubMed]
- Meissner, S.; Fastner, J.; Dittmann, E. Microcystin production revisited: Conjugate formation makes a major contribution. Environ. Microbiol. 2013, 15, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.M.; Lee, S.J.; Jang, M.H.; Yoon, B.D. Microcystin production by Microcystis aeruginosa in a phosphorus-limited chemostat. Appl. Environ. Microbiol. 2000, 66, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, E.; Martin-Luna, B.; Vela, L.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. Iron availability affects mcyD expression and microcystin-LR synthesis in Microcystis aeruginosa PCC7806. Environ. Microbiol. 2008, 10, 2476–2483. [Google Scholar] [CrossRef] [PubMed]
- Jahnichen, S.; Long, B.M.; Petzoldt, T. Microcystin production by Microcystis aeruginosa: Direct regulation by multiple environmental factors. Harmful Algae 2011, 12, 95–104. [Google Scholar] [CrossRef]
- Song, L.; Sano, T.; Li, R.; Watanabe, M.M.; Liu, Y.; Kaya, K. Microcystin production of Microcystis viridis (cyanobacteria) under different culture conditions. Physiol. Res. 1998, 46, 19–23. [Google Scholar]
- Pimentel, J.S.M.; Giani, A. Microcystin production and regulation under nutrient stress conditions in toxic Microcystis strains. Appl. Environ. Microbiol. 2014, 80, 5836–5843. [Google Scholar] [CrossRef] [PubMed]
- Kuiper-Goodman, T.; Falconer, I.; Fitzgerald, J. Human Health Aspects. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; E & FN Spon: London, UK, 1999. [Google Scholar]
- Zastepa, A.; Pick, F.R.; Blais, J.M. Fate and persistence of particulate and dissolved microcystin-LA from Microcystis blooms. Hum. Ecol. Risk Assess. 2014, 20, 1670–1686. [Google Scholar] [CrossRef]
- Pantelic, D.; Svircev, Z.; Simeunovic, J.; Vidovic, M.; Trajkovic, I. Cyanotoxins: Characteristics, production and degradation routes in drinking water treatment with reference to the situation in serbia. Chemosphere 2013, 91, 421–441. [Google Scholar] [CrossRef] [PubMed]
- Manage, P.M.; Edwards, C.; Singh, B.K.; Lawton, L.A. Isolation and identification of novel microcystin-degrading bacteria. Appl. Environ. Microbiol. 2009, 75, 6924–6928. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.W.; Sui, G.D.; Liu, S.X. Characteristics of a microcystin-LR biodegrading bacterial isolate: Ochrobactrum sp. FDT5. Bull. Environ. Contam. Toxicol. 2014, 92, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.L.; Yu, X.B.; Zhang, C.; Wang, G.X. Growth inhibition and microcystin degradation effects of acinetobacter guillouiae A2 on Microcystis aeruginosa. Res. Microbiol. 2015, 166, 93–101. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, R.L.; Aucamp, P.J.; Bais, A.F.; Bjorn, L.O.; Ilyas, M.; Madronich, S. Ozone depletion and climate change: Impacts on UV radiation. Photochem. Photobiol. Sci. 2011, 10, 182–198. [Google Scholar] [CrossRef] [PubMed]
- Shelly, K.; Heraud, P.; Beardall, J. Interactive effects of par and UV-B radiation on PSII electron transport in the marine alga Dunaliella tertiolecta (chlorophyceae). J. Phycol. 2003, 39, 509–512. [Google Scholar] [CrossRef]
- Lionard, M.; Roy, S.; Tremblay-Letourneau, M.; Ferreyra, G.A. Combined effects of increased UV-B and temperature on the pigment-determined marine phytoplankton community of the St. Lawrence Estuary. Mar. Ecol. Prog. Ser. 2012, 445, 219–234. [Google Scholar] [CrossRef]
- Hader, D.P.; Williamson, C.E.; Wangberg, S.A.; Rautio, M.; Rose, K.C.; Gao, K.S.; Helbling, E.W.; Sinha, R.P.; Worrest, R. Effects of UV radiation on aquatic ecosystems and interactions with other environmental factors. Photochem. Photobiol. Sci. 2015, 14, 108–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.B.; Qiu, B.S. Photosynthetic adaptation of a bloom-forming cyanobacterium Microcystis aeruginosa (cyanophyceae) to prolonged UV-B exposure. J. Phycol. 2005, 41, 983–992. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, H.B.; Qiu, B.S. Effects of UVB radiation on competition between the bloom-forming cyanobacterium Microcystis aeruginosa and the chlorophyceae chlamydomonas microsphaera. J. Phycol. 2013, 49, 318–328. [Google Scholar] [CrossRef]
- Ding, Y.; Song, L.R.; Sedmak, B. UVB radiation as a potential selective factor favoring microcystin producing bloom forming cyanobacteria. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Yang, Z.; Kong, F.X.; Shi, X.L.; Yu, Y.; Zhang, M. Effects of UV-B radiation on microcystin production of a toxic strain of Microcystis aeruginosa and its competitiveness against a non-toxic strain. J. Hazard. Mater. 2015, 283, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Zhang, Y.; Zhu, G.; He, K.; Feng, L.; Cai, L. Effect on the growth of Microcystis aeruginosa by the simulated continuous UV-B radiation. J. Lake Sci. 2011, 23, 223–229. [Google Scholar]
- Nouguier, J.; Mostajir, B.; Le Floc’h, E.; Vidussi, F. An automatically operated system for simulating global change temperature and ultraviolet B radiation increases: Application to the study of aquatic ecosystem responses in mesocosm experiments. Limnol. Oceanogr.: Methods 2007, 5, 269–279. [Google Scholar] [CrossRef]
- Boopathi, T.; Ki, J.S. Impact of environmental factors on the regulation of cyanotoxin production. Toxins 2014, 6, 1951–1978. [Google Scholar] [CrossRef] [PubMed]
- Neilan, B.A.; Pearson, L.A.; Muenchhoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Microbiol. 2013, 15, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Kuniyoshi, T.M.; Sevilla, E.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. Phosphate deficiency (N/P 40:1) induces mcyd transcription and microcystin synthesis in Microcystis aeruginosa PCC7806. Plant Physiol. Biochem. 2013, 65, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, C.; Pinzani, P.; Bianchi, S.; Paglierani, M.; Distante, V.; Pazzagli, M.; Bustin, S.A.; Orlando, C. Quantitative real-time reverse transcription polymerase chain reaction: Normalization to rRNA or single housekeeping genes is inappropriate for human tissue biopsies. Anal. Biochem. 2002, 309, 293–300. [Google Scholar] [CrossRef]
- Lee, M.A.; Guo, R.Y.; Ebenezer, V.; Ki, J.S. Evaluation and selection of reference genes for ecotoxicogenomic study of the green alga Closterium ehrenbergii using quantitative real-time PCR. Ecotoxicology 2015, 24, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.F.; Yu, S.Q.; Sun, Z.Q.; Xie, X.C.; Liu, W.P.; Fu, Z.W. Effects of copper sulfate, hydrogen peroxide and N-phenyl-2-naphthylamine on oxidative stress and the expression of genes involved photosynthesis and microcystin disposition in Microcystis aeruginosa. Aquat. Toxicol. 2010, 99, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Phelan, R.R.; Downing, T.G. A growth advantage for microcystin production by Microcystis PCC7806 under high light. J. Phycol. 2011, 47, 1241–1246. [Google Scholar] [CrossRef]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Borner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of Microcystis under oxidative stress conditions. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Gan, N.Q.; Xiao, Y.; Zhu, L.; Wu, Z.X.; Liu, J.; Hu, C.L.; Song, L.R. The role of microcystins in maintaining colonies of bloom-forming Microcystis spp. Environ. Microbiol. 2012, 14, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Kaebernick, M.; Dittmann, E.; Borner, T.; Neilan, B.A. Multiple alternate transcripts direct the biosynthesis of microcystin, a cyanobacterial nonribosomal peptide. Appl. Environ. Microbiol. 2002, 68, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, K.; Watanuki, T.; Kondo, F.; Watanabe, M.F.; Suzuki, S.; Nakazawa, H.; Suzuki, M.; Uchida, H.; Harada, K. Stability of microcystins from cyanobacteria-II. Effect of UV light on decomposition and isomerization. Toxicon 1995, 33, 1619–1631. [Google Scholar] [CrossRef]
- Jacobs, L.C.V.; Peralta-Zamora, P.; Campos, F.R.; Pontarolo, R. Photocatalytic degradation of microcystin-LR in aqueous solutions. Chemosphere 2013, 90, 1552–1557. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; Steinberg, C. Indirect photolysis of cyanotoxins: One possible mechanism for their low persistence. Water Res. 1999, 33, 1159–1164. [Google Scholar] [CrossRef]
- Westrick, J.A.; Szlag, D.C.; Southwell, B.J.; Sinclair, J. A review of cyanobacteria and cyanotoxins removal/inactivation in drinking water treatment. Anal. Bioanal. Chem. 2010, 397, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.; Onstad, G.; von Gunten, U.; Rinck-Pfeiffer, S.; Craig, K.; Newcombe, G. Differences in the chlorine reactivity of four microcystin analogues. Water Res. 2006, 40, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Pelaez, M.; Falaras, P.; Kontos, A.G.; de la Cruz, A.A.; O’shea, K.; Dunlop, P.S.M.; Byrne, J.A.; Dionysiou, D.D. A comparative study on the removal of cylindrospermopsin and microcystins from water with NF-TiO2-P25 composite films with visible and UV-vis light photocatalytic activity. Appl. Catal. B-Environ. 2012, 121, 30–39. [Google Scholar] [CrossRef]
- Furukawa, K.; Noda, N.; Tsuneda, S.; Saito, T.; Itayama, T.; Inamori, Y. Highly sensitive real-time PCR assay for quantification of toxic cyanobacteria based on microcystin synthetase a gene. J. Biosci. Bioeng. 2006, 102, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Neilan, B.A.; Jacobs, D.; DelDot, T.; Blackall, L.L.; Hawkins, P.R.; Cox, P.T.; Goodman, A.E. rRNA sequences and evolutionary relationships among toxic and nontoxic cyanobacteria of the genus Microcystis. Int. J. Syst. Bacteriol. 1997, 47, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wiedner, C.; Visser, P.M.; Fastner, J.; Metcalf, J.S.; Codd, G.A.; Mur, L.R. Effects of light on the microcystin content of Microcystis strain PCC7806. Appl. Environ. Microbiol. 2003, 69, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Kong, F. UV-B Exposure Affects the Biosynthesis of Microcystin in Toxic Microcystis aeruginosa Cells and Its Degradation in the Extracellular Space. Toxins 2015, 7, 4238-4252. https://doi.org/10.3390/toxins7104238
Yang Z, Kong F. UV-B Exposure Affects the Biosynthesis of Microcystin in Toxic Microcystis aeruginosa Cells and Its Degradation in the Extracellular Space. Toxins. 2015; 7(10):4238-4252. https://doi.org/10.3390/toxins7104238
Chicago/Turabian StyleYang, Zhen, and Fanxiang Kong. 2015. "UV-B Exposure Affects the Biosynthesis of Microcystin in Toxic Microcystis aeruginosa Cells and Its Degradation in the Extracellular Space" Toxins 7, no. 10: 4238-4252. https://doi.org/10.3390/toxins7104238
APA StyleYang, Z., & Kong, F. (2015). UV-B Exposure Affects the Biosynthesis of Microcystin in Toxic Microcystis aeruginosa Cells and Its Degradation in the Extracellular Space. Toxins, 7(10), 4238-4252. https://doi.org/10.3390/toxins7104238