
Relationship of Deoxynivalenol Content in Grain, Chaff, and Straw with Fusarium Head Blight Severity in Wheat Varieties with Various Levels of Resistance

Abstract
:1. Introduction
2. Results
2.1. FHB Severity in Wheat Cultivars

{kind=link}
{kind=link}
{kind=link}
| NO. | Accession name | Origin | PIS 1 |
|---|---|---|---|
| 1 | Loyal | Sweden | 31.69 ± 10.56 |
| 2 | Hereford | Sweden | 24.34 ± 7.64 |
| 3 | Skagen | Sweden | 23.35 ± 6.04 |
| 4 | PAJ706-575A | Sweden | 45.82 ± 22.35 |
| 5 | Mariboss | Sweden | 29.96 ± 22.27 |
| 6 | Harnesk-7 | Sweden | 65.15 ± 11.67 |
| 7 | IDUNA | Sweden | 39.17 ± 16.00 |
| 8 | ANKAR | Sweden | 38.53 ± 20.22 |
| 9 | ÅRING III | Sweden | 100.00 |
| 10 | EROICA | Sweden | 22.61 ± 19.92 |
| 11 | AROS | Sweden | 33.08 ± 22.40 |
| 12 | BANCO | Sweden | 16.86 ± 8.73 |
| 13 | NORRE | Sweden | 12.19 ± 8.80 |
| 14 | STURE | Sweden | 56.28 ± 31.01 |
| 15 | HELGE | Sweden | 14.40 ± 0.73 |
| 16 | LINNA | Sweden | 29.39 ± 12.89 |
| 17 | JYVÄ | Sweden | 95.14 ± 6.82 |
| 18 | SIGYN II | Sweden | 100 ± 0.00 |
| 19 | FOLKE | Sweden | 32.61 ± 13.48 |
| 20 | HOLGER | Sweden | 24.12 ± 9.90 |
| 21 | ALEMAR UST. HV. SG. | Sweden | 61.48 ± 13.55 |
| 22 | GUSALEK K.17 | Sweden | 96.07 ± 6.79 |
| 23 | LANTVETE FRÅN UPPSALA | Sweden | 100 ± 0.00 |
| 24 | KOTTE | Sweden | 53.70 ± 11.88 |
| 25 | EXTRA SQUAREHEAD | Sweden | 47.02 ± 11.75 |
| 26 | RENODLAT SAMMETSVETE | Sweden | 91.61 ± 6.20 |
| 27 | THULE II | Sweden | 92.81 ± 11.84 |
| 28 | PANSAR II | Sweden | 29.68 ± 6.63 |
| 29 | SVEA I | Sweden | 97.27 ± 6.10 |
| 30 | PANSAR III | Sweden | 79.84 ± 10.06 |
| 31 | GYLLEN II | Sweden | 85.29 ± 10.23 |
| 32 | GLUTEN | Sweden | 93.75 ± 12.50 |
| 33 | BORG | Sweden | 89.91 ± 14.39 |
| 34 | PÄRL II | Sweden | 37.50 ± 17.68 |
| 35 | ROBUR | Sweden | 26.34 ± 10.49 |
| 36 | SEBA | Sweden | 68.30 ± 11.44 |
| 37 | VIRGO | Sweden | 88.38 ± 18.02 |
| 38 | SOLID | Sweden | 40.82 ± 15.72 |
| 39 | HILDUR | Sweden | 17.09 ± 6.66 |
| 40 | HANKKIJAN ILVES | Sweden | 39.52 ± 10.72 |
| 41 | Harnesk-5 | Sweden | 24.68 ± 9.89 |
| 42 | GUSALEK K.10 A | Sweden | 38.41 ± 23.08 |
| 43 | KOSACK | Sweden | 46.04 ± 11.00 |
| 44 | MK 2-114 | Sweden | 74.57 ± 11.86 |
| 45 | MK 2-302 | Sweden | 32.31 ± 6.84 |
| 46 | MK 2-304 | Sweden | 85.23 ± 17.94 |
| 47 | MK 2-306 | Sweden | 95.56 ± 9.94 |
| 48 | MK 2-313 | Sweden | 27.27 ± 6.43 |
| 49 | MK 2-502 | Sweden | 41.36 ± 10.15 |
| 50 | MK 2-503 | Sweden | 100.00 ± 0.00 |
| 51 | MK 2-506 | Sweden | 46.11 ± 16.35 |
| 52 | MK 2-508 | Sweden | 31.11 ± 12.57 |
| 53 | MK 2-542 | Sweden | 59.82 ± 15.41 |
| 54 | MK 2-543 | Sweden | 83.43 ± 9.19 |
| 55 | MK 2-547 | Sweden | 97.37 ± 5.26 |
| 56 | MK 2-548 | Sweden | 21.67 ± 21.62 |
| 57 | MK 2-556 | Sweden | 90.73 ± 11.73 |
| 58 | MK 2-557 | Sweden | 77.06 ± 11.22 |
| 59 | MK 2-558 | Sweden | 84.31 ± 13.25 |
| 60 | MK 2-564 | Sweden | 63.92 ± 25.45 |
| 61 | SALUT | Sweden | 53.91 ± 12.70 |
| 62 | ØSTBY | Sweden | 30.81 ± 9.26 |
| 63 | MK 2-501 | Sweden | 59.04 ± 11.22 |
| 64 | MK 2-510 | Sweden | 94.18 ± 8.85 |
| 65 | PORTAL | Sweden | 30.01 ± 10.00 |
| 66 | TJELVAR | Sweden | 45.51 ± 9.95 |
| 67 | TRYGGVE | Sweden | 88.81 ± 10.62 |
| 68 | MK 2-549 | Sweden | 79.50 ± 11.95 |
| 69 | MK 2-566 | Sweden | 91.67 ± 14.43 |
| 70 | MK 2-651 | Sweden | 42.84 ± 11.66 |
| 71 | MK 2-656 | Sweden | 66.51 ± 28.36 |
| 72 | MK 2-659 | Sweden | 39.18 ± 13.15 |
| 73 | MK 2-660 | Sweden | 61.33 ± 21.33 |
| 74 | MK 2-775 | Sweden | 44.96 ± 7.58 |
| 75 | MK 2-780 | Sweden | 88.18 ± 21.70 |
| 76 | MK 2-786 | Sweden | 86.93 ± 15.12 |
| 77 | MK 2-787 | Sweden | 44.74 ± 12.32 |
| 78 | PANU | Sweden | 65.90 ± 34.88 |
| 79 | KONGE III | Sweden | 37.50 ± 15.52 |
| 80 | TYSTOFTE SMAAHVEDE | Sweden | 34.03 ± 10.88 |
| 81 | RENTAL | Sweden | 57.33 ± 13.20 |
| 82 | STAVA | Sweden | 53.88 ± 15.44 |
| 83 | RUDOLF RUBIN | Sweden | 90.58 ± 13.79 |
| 84 | KIRSTEN | Sweden | 42.80 ± 11.91 |
| 85 | LONE | Sweden | 43.68 ± 8.71 |
| 86 | BRANDT | Sweden | 40.56 ± 11.73 |
| 87 | KARAT | Sweden | 39.61 ± 17.13 |
| 88 | MK 2-101 | Sweden | 41.43 ± 17.05 |
| 89 | MK 2-113 | Sweden | 49.57 ± 24.98 |
| 90 | MK 2-116 | Sweden | 71.81 ± 24.64 |
| 91 | MK 2-122 | Sweden | 94.44 ± 13.61 |
| 92 | MK 2-130 | Sweden | 29.91 ± 10.39 |
| 93 | MK 2-316 | Sweden | 47.09 ± 15.38 |
| 94 | MK 2-317 | Sweden | 69.01 ± 13.76 |
| 95 | MK 2-337 | Sweden | 64.83 ± 15.38 |
| 96 | MK 2-567 | Sweden | 57.99 ± 39.13 |
| 97 | MK 2-655 | Sweden | 68.97 ± 26.56 |
| 98 | MK 2-138 | Sweden | 32.40 ± 16.47 |
| 99 | MK 2-679 | Sweden | 81.00 ± 7.12 |
| 100 | MK 2-788 | Sweden | 70.70 ± 10.31 |
| 101 | MK 2-847 | Sweden | 25.99 ± 13.17 |
| 102 | Saxild | Sweden | 37.17 ± 18.92 |
| 103 | Abba | Sweden | 33.86 ± 4.74 |
| 104 | Konsul | Sweden | 100.00 ± 0.00 |
| 105 | Rektor | Sweden | 35.89 ± 8.00 |
| 106 | Stakado | Sweden | 100.00 ± 0.00 |
| 107 | Kerimäkeläinen | Sweden | 95.45 ± 9.09 |
| 108 | Sampo | Sweden | 95.83 ± 9.32 |
| 109 | Väinö | Sweden | 87.20 ± 11.37 |
| 110 | Speltti Vaalea | Sweden | 67.79 ± 19.61 |
| 111 | Speltti Ruskea Baulander | Sweden | 56.30 ± 18.53 |
| 112 | Kökar | Sweden | 31.96 ± 8.09 |
| 113 | Olympia | Sweden | 77.45 ± 13.20 |
| 114 | H 8703 | Sweden | 100.00 ± 0.00 |
| 115 | H 8196 | Sweden | 100.00 ± 0.00 |
| 116 | H 8202 | Sweden | 100.00 ± 0.00 |
| 117 | H 8222 | Sweden | 76.78 ± 35.98 |
| 118 | H 8296 | Sweden | 34.72 ± 28.69 |
| 119 | H 8300 | Sweden | 88.75 ± 15.56 |
| 120 | H 8305 | Sweden | 19.57 ± 7.03 |
| 121 | H 8310 | Sweden | 94.44 ± 9.62 |
| 122 | H 8311 | Sweden | 33.55 ± 7.08 |
| - | Control cultivar | - | - |
| - | Annong 8455 | China | 89.65 ± 2.33 |
| - | Sumai 3 | China | 24.21 ± 9.04 |
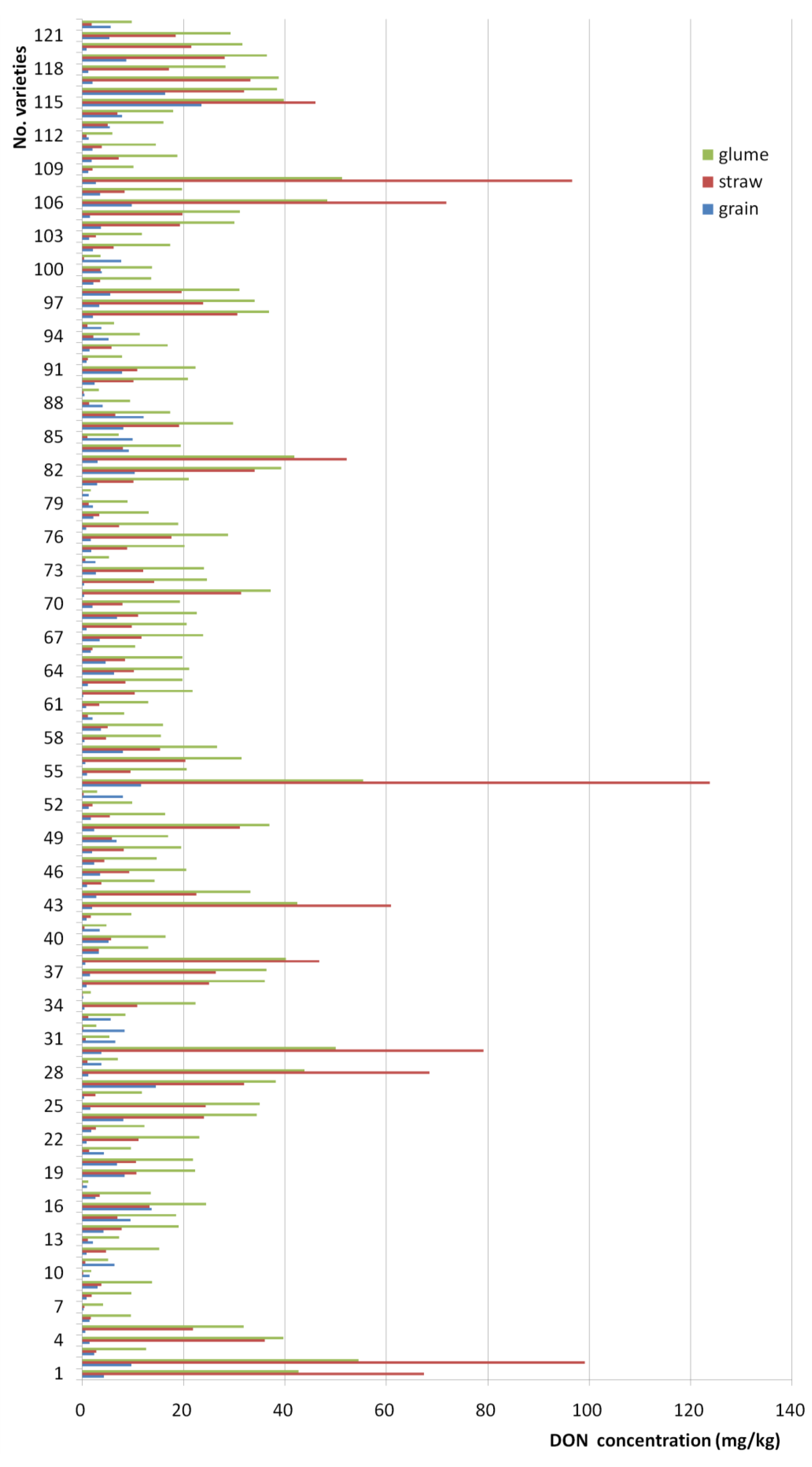
2.2. Deoxynivalenol (DON) Levels in Wheat Grain, Straw, and Glume Tissues after Artificial Inoculation of Wheat Ears
| DON (mg/kg) | |||||
|---|---|---|---|---|---|
| Tissue | n | Mean * | Median | Max | Min |
| grain | 122 | 3.88 A | 2.38 | 23.46 | 0.16 |
| straw | 122 | 15.30 B | 7.85 | 123.74 | 0.04 |
| glume | 122 | 20.95 C | 19.13 | 55.42 | 1.15 |

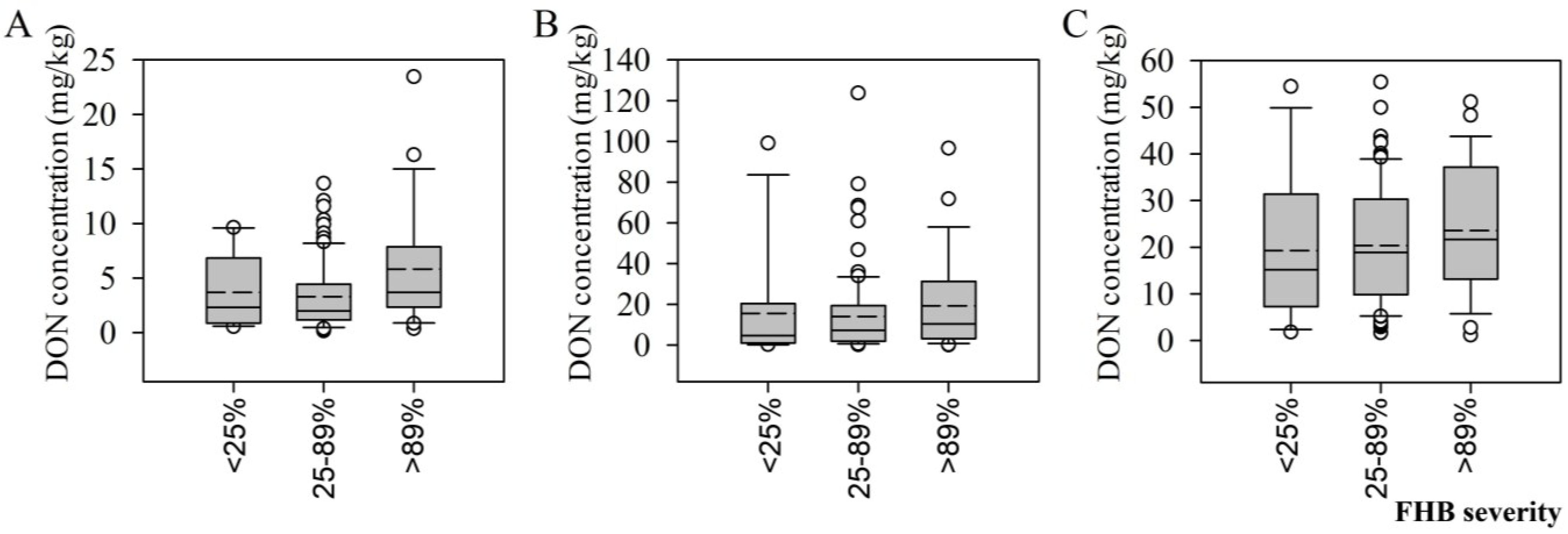
2.3. DON Levels in Tissues from Wheat Varieties Exhibiting Differing FHB Disease Severity

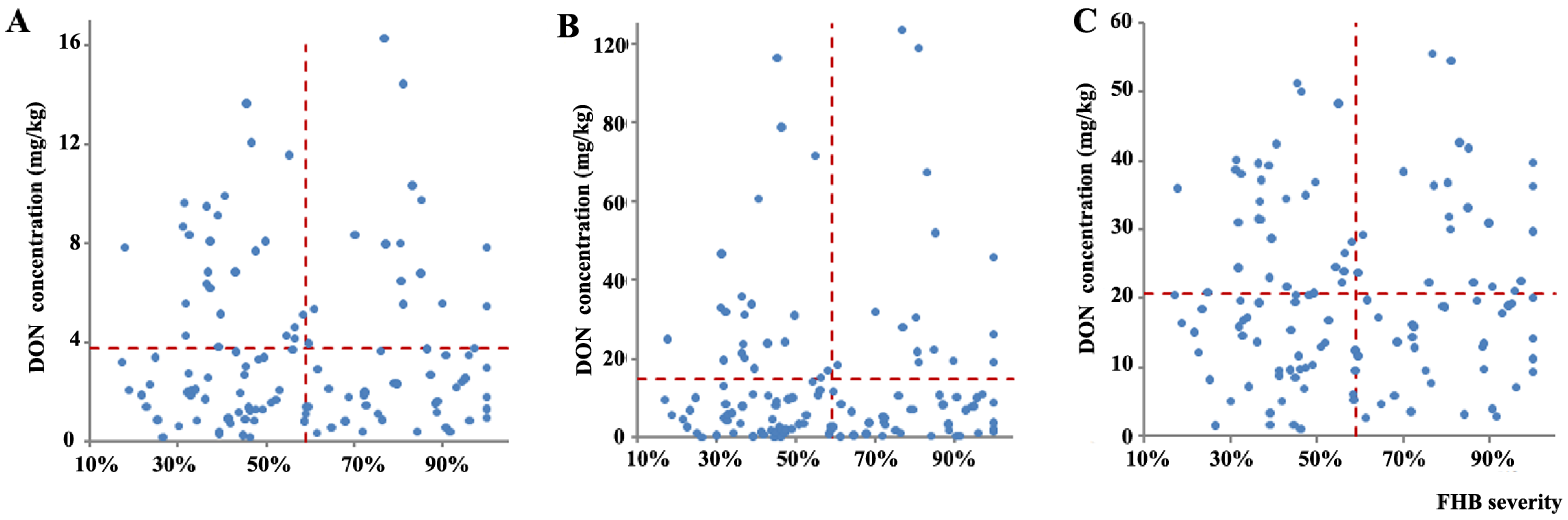
2.4. Relationship between FHB Severity and DON Levels among Wheat Varieties and Selection of Wheat Genotypes with Reduced DON Content

3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Plant Material
4.3. Mycotoxin Analysis
4.3.1. Preparation of Analytical Standard Solutions
4.3.2. Sample Preparation
4.3.3. LC-MS/MS Analysis
4.4. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bai, G.; Shaner, G. Scab of wheat: Prospects for control. Plant Dis. 1994, 78, 760–766. [Google Scholar] [CrossRef]
- McMullen, M.; Jones, R.; Gallenberg, D. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar] [CrossRef]
- Parry, D.; Jenkinson, P.; McLeod, L. Fusarium ear blight (scab) in small grain cereals—A review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Xu, X.-M.; Berrie, A. Epidemiology of mycotoxigenic fungi associated with fusarium ear blight and apple blue mould: A review. Food Addit. Contam. 2005, 22, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Opinion of the scientific panel on contaminants in the food chain on a request from the commission related to deoxynivalenol(don) as undesirable substance in animal feed. EFSA 2004, 73, 1–41. [Google Scholar]
- Rotter, B.A.; Prelusky, D.B.; Pestka, J.L. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Snijders, C.; Perkowski, J. Effects of head blight caused by fusarium culmorum on toxin content and weight of wheat kernels. Phytopathology 1990, 80, 566–570. [Google Scholar] [CrossRef]
- Hussein, H.S.; Brasel, J.M. Toxicity, metabolism, and impact of mycotoxins on humans and animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef] [PubMed]
- Maier, F.J.; Miedaner, T.; Hadeler, B.; Felk, A.; Salomon, S.; Lemmens, M.; Kassner, H.; Schäfer, W. Involvement of trichothecenes in fusarioses of wheat, barley and maize evaluated by gene disruption of the trichodiene synthase (Tri5) gene in three field isolates of different chemotype and virulence. Mol. Plant Pathol. 2006, 7, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.; Tekauz, A. Review: Recent developments in research on fusarium head blight of wheat in Canada. Can. J. Plant Pathol. 2000, 22, 1–8. [Google Scholar] [CrossRef]
- National Criterion of China. Maximum levels of mycotoxins in foods 2001. GB/T 2761–2011. Available online: http://down.foodmate.net/standard/sort/3/28221.html (accessed on 20 April 2011).
- Kang, Z.; Buchenauer, H. Ultrastructural and cytochemical studies on cellulose, xylan and pectin degradation in wheat spikes infected by Fusarium culmorum. J. Phytopathol. 2000, 148, 263–275. [Google Scholar] [CrossRef]
- Matthäus, K.; Dänicke, S.; Vahjen, W.; Simon, O.; Wang, J.; Valenta, H.; Meyer, K.; Strumpf, A.; Zieseni, H.; Flachowsky, G. Progression of mycotoxin and nutrient concentrations in wheat after inoculation with Fusarium culmorum. Arch. Anim. Nutr. 2004, 58, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Seeling, K.; Boguhn, J.; Strobel, E.; Danicke, S.; Valenta, H.; Ueberschar, K.H.; Rodehutscord, M. On the effects of Fusarium toxin contaminated wheat and wheat chaff on nutrient utilisation and turnover of deoxynivalenol and zearalenone in vitro (Rusitec). Toxicol. In Vitro 2006, 20, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D.; Young, J.C.; Sampson, D.R. Deoxynivalenol and Fusarium head blight resistance in spring cereals. Phytopath. Z. 1985, 113, 359–367. [Google Scholar] [CrossRef]
- Mirocha, C.J.; Xie, W.; Xu, Y.; Wilcoxson, R.D.; Woodward, R.P.; Etebarian, R.H.; Behele, G. Production of trichothecene mycotoxins by Fusarium graminearum and Fusarium culmorum on barley and wheat. Mycopathologia 1994, 128, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-Z.; Langseth, W.; Skinnes, H.; Elen, O.N.; Sundheim, L. Comparison of visual head blight ratings-seed infection levels and deoxynivalenol production for assessment of resistance in cereals inoculated with Fusarium culmorum. Eur. J. Plant Pathol. 1997, 103, 589–595. [Google Scholar] [CrossRef]
- Hestbjerg, H.; Felding, G.; Elmholt, S. Fusarium culmorum infection of barley seedlings: Correlation between aggressiveness and deoxynivalenol content. J. Phytopathol. 2002, 150, 308–312. [Google Scholar] [CrossRef]
- Bai, G.H.; Plattner, R.; Desjardins, A.; Kolb, F.; McIntosh, R. Resistance to fusarium head blight and deoxynivalenol accumulation in wheat. Plant Breed. 2001, 120, 1–6. [Google Scholar] [CrossRef]
- Ji, F.; Xu, J.; Liu, X.; Yin, X.; Shi, J. Natural occurrence of deoxynivalenol and zearalenone in wheat from jiangsu province, china. Food Chem. 2014, 157, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.; Christensen, J. Factors affecting resistance of wheat to scab caused by gibberella zeae. Phytopathology 1963, 53, 831–838. [Google Scholar]
- Miller, J.; Young, J.; Sampson, D. Deoxynivalenol and fusarium head blight resistance in spring cereals. J. Phytopathol. 1985, 113, 359–367. [Google Scholar] [CrossRef]
- Cumagun, C.J.R.; Miedaner, T. Segregation for aggressiveness and deoxynivalenol production of a population of gibberella zeae causing head blight of wheat. Eur. J. Plant Pathol. 2004, 110, 789–799. [Google Scholar] [CrossRef]
- Snijders, C.H.A. Resistance in wheat to fusarium infection and trichothecene formation. Toxicol. Lett. 2004, 153, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Homdork, S.; Fehrmann, H.; Beck, R. Effects of field application of tebuconazole on yield, yield components and the mycotoxin content of fusarium-infected wheat grain. J. Phytopathol. 2000, 148, 1–6. [Google Scholar] [CrossRef]
- Miedaner, T.; Schneider, B.; Geiger, H.H. Deoxynivalenol (don) content and fusarium head blight resistance in segregating populations of winter rye and winter wheat. Crop Sci. 2003, 43, 519–526. [Google Scholar] [CrossRef]
- Mesterhazy, A.; Logrieco, A.; Corazza, L.; Cooke, B.M. Role of deoxynivalenol in aggressiveness of Fusarium graminearum and F. Culmorum and in resistance to Fusarium head blight. Eur. J. Plant Pathol. 2002, 108, 675–684. [Google Scholar] [CrossRef]
- Lamper, C.; Teren, J.; Bartok, T.; Komoroczy, R.; Mesterhazy, A.; Sagi, F. Predicting don contamination in fusarium-infected wheat grains via determination of the ergosterol content. Cereal Res. Commun. 2000, 28, 337–344. [Google Scholar]
- Chen, L.; Song, Y.; Xu, Y. Variation in the concentrations of deoxynivalenol in the spikes of winter wheat infected by fusarium graminearum schw. Acta Phytopathol. Sin. 1995, 26, 25–28. [Google Scholar]
- Magan, N.; Hope, R.; Colleate, A.; Baxter, E.S. Relationship between growth and mycotoxin production by fusarium species, biocides and environment. Eur. J. Plant Pathol. 2002, 108, 685–690. [Google Scholar] [CrossRef]
- Champeil, A.; Fourbet, J.; Dore, T.; Rossignol, L. Influence of cropping system on fusarium head blight and mycotoxin levels in winter wheat. Crop Prot. 2004, 23, 531–537. [Google Scholar] [CrossRef]
- Birzele, B.; Meier, A.; Hindorf, H.; Krämer, J.; Dehne, H.W. Epidemiology of fusarium infection and deoxynivalenol content in winter wheat in the rhineland, germany. Eur. J. Plant Pathol. 2002, 108, 667–673. [Google Scholar] [CrossRef]
- Beyer, M.; Klix, M.B.; Verreet, J.-A. Estimating mycotoxin contents of fusarium-damaged winter wheat kernels. Int. J. Food Microbiol. 2007, 119, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Hope, R.; Aldred, D.; Magan, N. Comparison of environmental profies for growth and deoxynivalenol production by fusarium culmorum and f. Graminearum on wheat grain. Lett. Appl. Microbiol. 2005, 40, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Snijders, C.H.A.; Krechting, C.F. Inhibition of deoxynivalenol translocation and fungal colonization in fusarium head blight resistant wheat. Can. J. Bot. 1992, 70, 1570–1576. [Google Scholar] [CrossRef]
- Doohan, F.M.; Weston, G.; Rezanoor, H.N.; Parry, D.W.; Nicholson, P. Development and use of a reverse transcription-pcr assay to study expression of tri5 by fusarium species in vitro and in planta. Appl. Environ. Microbiol. 1999, 65, 3850–3854. [Google Scholar] [PubMed]
- Clear, R.M.; Patrick, S.K.; Vowicki, T.; Gaba, D.; Edney, M.; Babb, J.C. The effect of hull removal and pearling on fusarium species and trichothecenes in hulless barley. Can. J. Plant Sci. 1997, 77, 161–166. [Google Scholar] [CrossRef]
- Legzdina, L.; Buerstmayr, H. Comparison of infection with Fusarium head blight and accumulation of mycotoxins in grain of hulless and covered barley. J. Cereal Sci. 2004, 40, 61–67. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Miller, J.D. Effects of fusarium graminearum metabolites on wheat tissue in relation to fusarium head blight resistance. J. Phytopathol. 1988, 122, 118–125. [Google Scholar] [CrossRef]
- Sinha, R.; Savard, M. Concentration of deoxynivalenol in single kernels and various tissues of wheat heads. Can. J. Plant Pathol. 1997, 19, 8–12. [Google Scholar] [CrossRef]
- Bateman, G.; Murray, G.; Gutteridge, R.; Coşlkun, H. Effects of method of straw disposal and depth of cultivation on populations of fusarium spp. In soil and on brown foot rot in continuous winter wheat. Ann. Appl. Biol. 1998, 132, 35–47. [Google Scholar] [CrossRef]
- Zhang, X.; Ren, L.; Tan, X.; Zang, Y.; Ma, H.; Qin, J.; Lu, W. Evaluation method of wheat Fusarium head blight resistance for genetic populations based on molecular markers. J. Nanjing Univ. (Natl. Sci.) 2005, 41, 125–132. [Google Scholar]
- Ye, X.; Shirley, S.; Xu, H.; Du, L.; Huang, Y.; Lu, W.; Clemente, T. Transformation and identification of BCL and RlP genes related to cell apodosis into wheaf mediated by Agrobacterium. Acta Agron. Sin. 2005, 31, 1389–1393. [Google Scholar]
- Shi, J.; Xu, D.; Yang, H.; Lu, Q.; Ban, T. DNA marker analysis for pyramided of fusarium head blight (fhb) resistance qtls from different germplasm. Genetica 2008, 133, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Szabo-Hever, A.; Bjørnstad, A.; Lillemo, M.; Semagn, K.; Mesterhazy, A.; Ji, F.; Shi, J.; Skinnes, H. Two major resistance quantitative trait loci are required to counteract the increased susceptibility to Fusarium head blight of the dwarfing gene in wheat. Crop Sci. 2011, 51, 2430–2438. [Google Scholar] [CrossRef]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef] [PubMed]
- Spanjer, M.C.; Rensen, P.M.; Scholten, J.M. Lc–ms/ms multi-method for mycotoxins after single extraction, with validation data for peanut, pistachio, wheat, maize, cornflakes, raisins and figs. Food Addit. Contam. 2008, 25, 472–489. [Google Scholar] [CrossRef]
- Soleimany, F.; Jinap, S.; Abas, F. Determination of mycotoxins in cereals by liquid chromatography tandem mass spectrometry. Food Chem. 2012, 130, 1055–1060. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, F.; Wu, J.; Zhao, H.; Xu, J.; Shi, J. Relationship of Deoxynivalenol Content in Grain, Chaff, and Straw with Fusarium Head Blight Severity in Wheat Varieties with Various Levels of Resistance. Toxins 2015, 7, 728-742. https://doi.org/10.3390/toxins7030728
Ji F, Wu J, Zhao H, Xu J, Shi J. Relationship of Deoxynivalenol Content in Grain, Chaff, and Straw with Fusarium Head Blight Severity in Wheat Varieties with Various Levels of Resistance. Toxins. 2015; 7(3):728-742. https://doi.org/10.3390/toxins7030728
Chicago/Turabian StyleJi, Fang, Jirong Wu, Hongyan Zhao, Jianhong Xu, and Jianrong Shi. 2015. "Relationship of Deoxynivalenol Content in Grain, Chaff, and Straw with Fusarium Head Blight Severity in Wheat Varieties with Various Levels of Resistance" Toxins 7, no. 3: 728-742. https://doi.org/10.3390/toxins7030728
APA StyleJi, F., Wu, J., Zhao, H., Xu, J., & Shi, J. (2015). Relationship of Deoxynivalenol Content in Grain, Chaff, and Straw with Fusarium Head Blight Severity in Wheat Varieties with Various Levels of Resistance. Toxins, 7(3), 728-742. https://doi.org/10.3390/toxins7030728