
The Effect of Cyanobacterial Biomass Enrichment by Centrifugation and GF/C Filtration on Subsequent Microcystin Measurement


 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
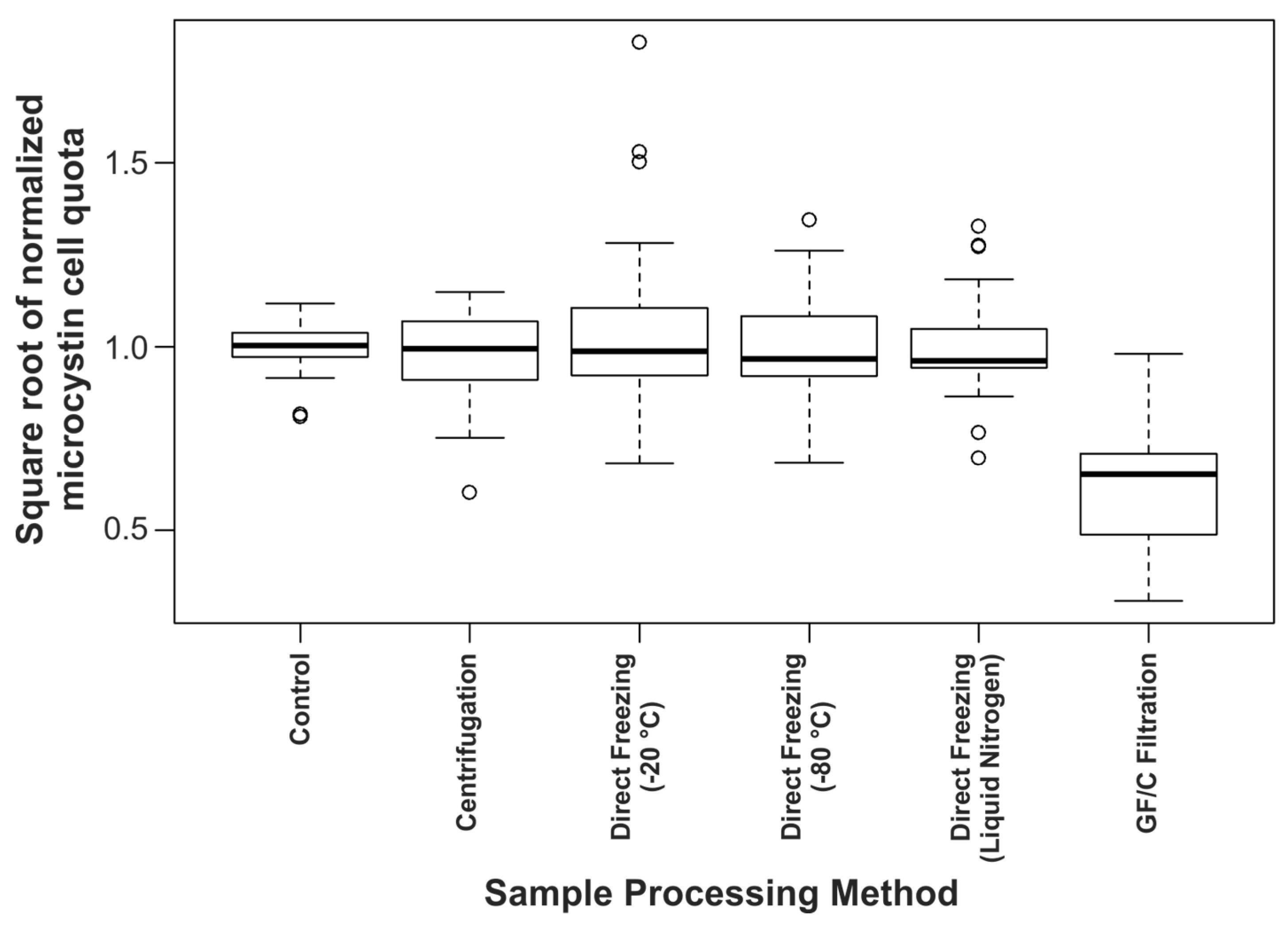
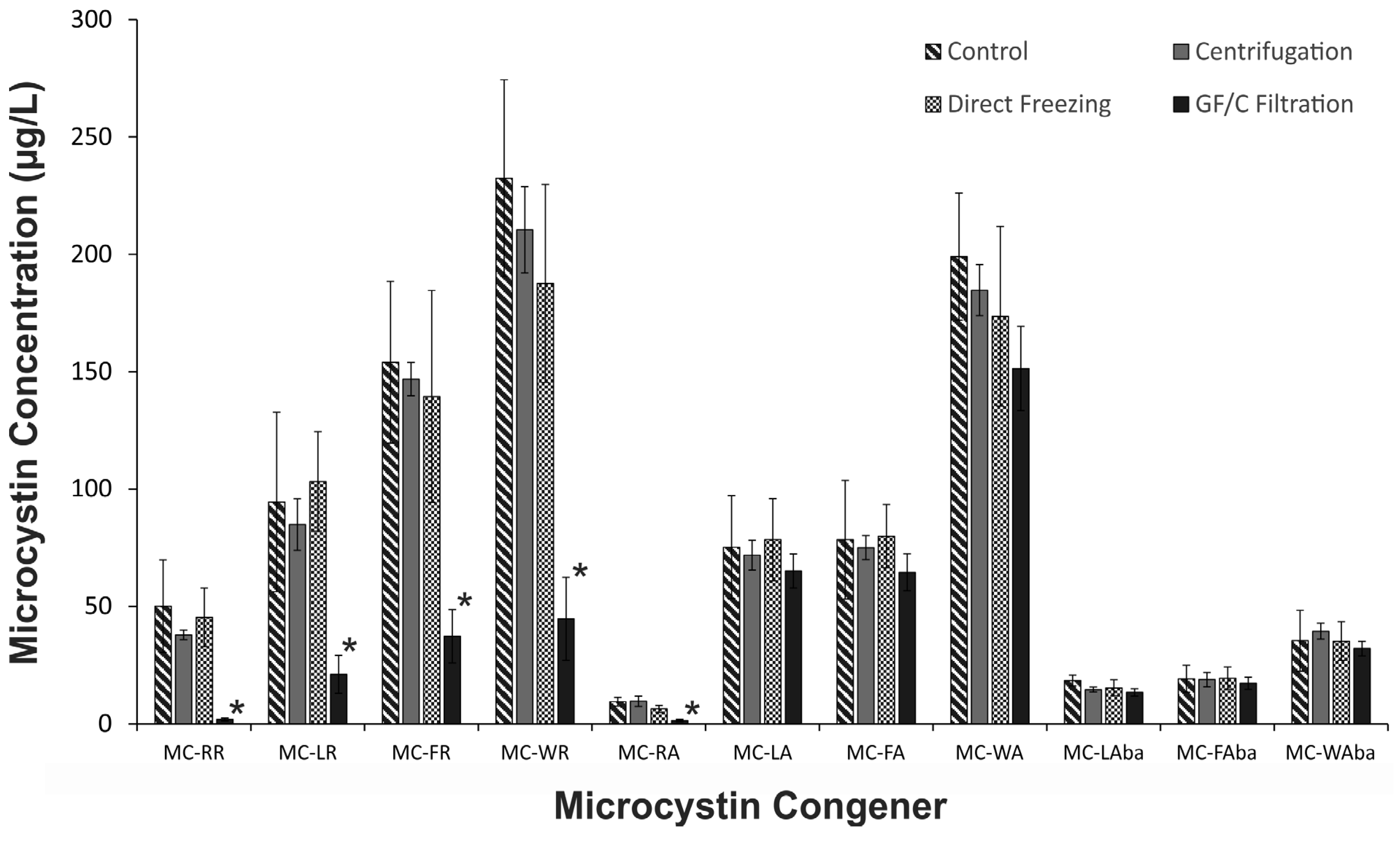
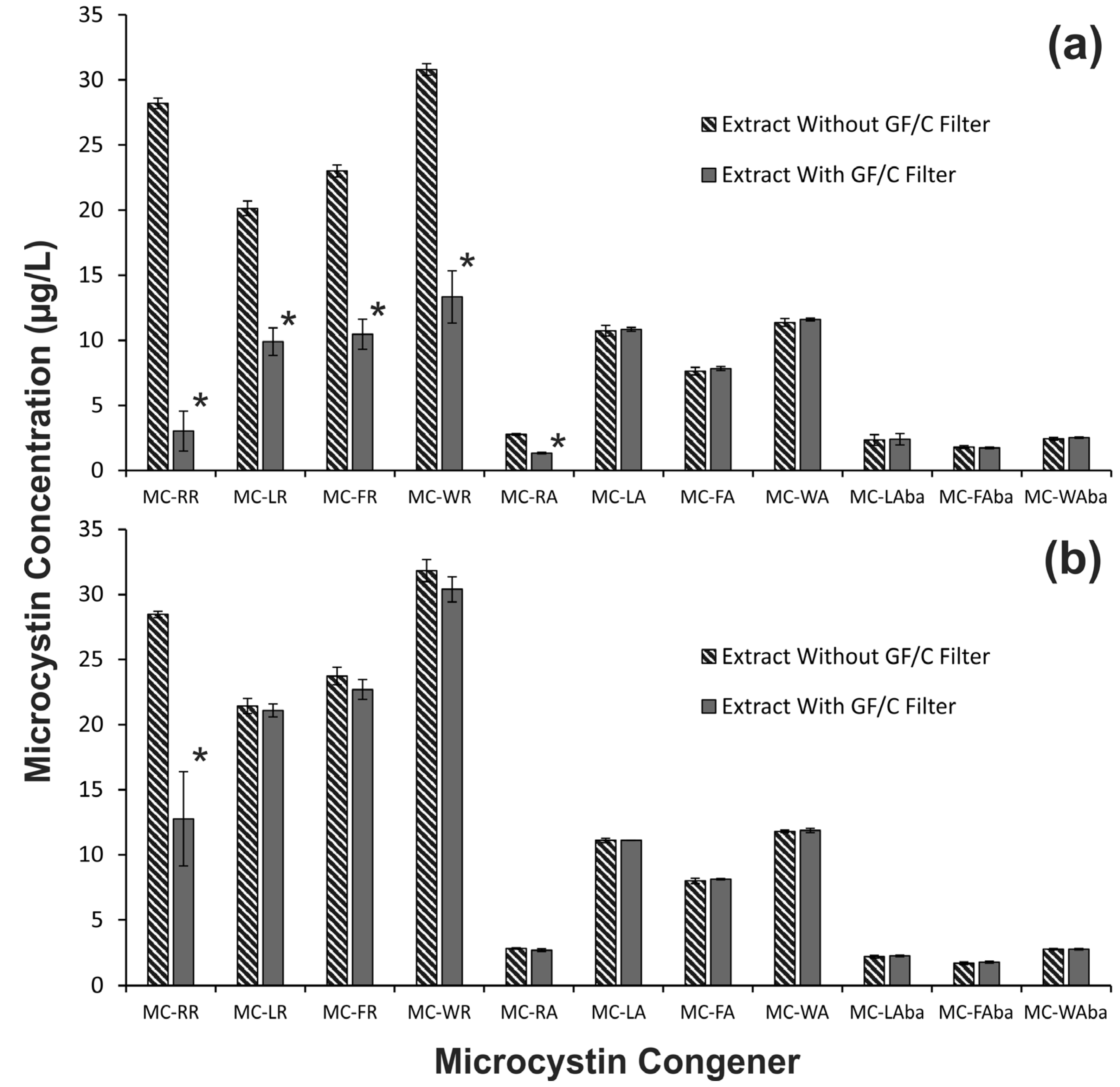
2.1. The Effect of Cyanobacterial Biomass Enrichment through Sample Processing Procedures


2.2. Investigation of the Low Microcystin Yields from GF/C Filtration Samples


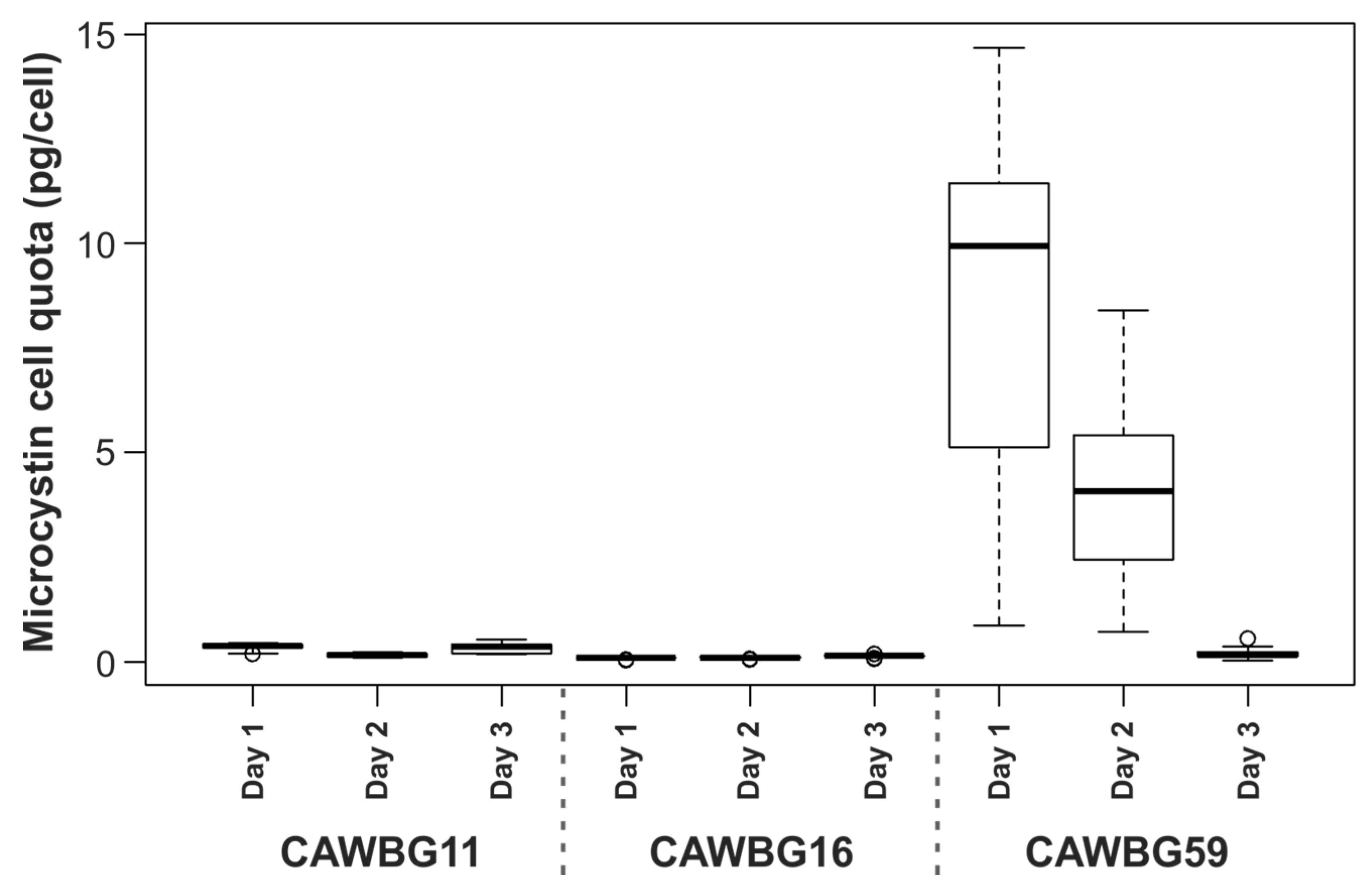
2.3. Temporal Effect of Sample Centrifugation on Microcystin Metabolism

2.4. Sample Processing Considerations
3. Experimental Section
3.1. Cyanobacterial Strains
3.2. Comparison of Sample Processing Methods
3.3. Microcystin Adherence to GF/C Filters
3.4. Temporal Effect of Centrifugation
3.5. Microcystin Analysis
3.6. Cell Enumeration
3.7. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; CRC Press: London, UK, 1999. [Google Scholar]
- Falconer, I.R.; Yeung, D.S.K. Cytoskeletal changes in hepatocytes induced by Microcystis toxins and their relation to hyperphosphorylation of cell proteins. Chem. Biol. Interact. 1992, 81, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, S.; Matsushima, R.; Watanabe, M.F.; Harada, K.-I.; Ichihara, A.; Carmichael, W.W.; Fujiki, H. Inhibition of protein phosphatases by microcystis and nodularin associated with hepatotoxicity. J. Cancer Res. Clin. Oncol. 1990, 116, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Ueno, Y.; Nagata, S.; Tsutsumi, T.; Hasegawa, A.; Watanabe, M.F.; Park, H.-D.; Chen, G.-C.; Chen, G.; Yu, S.-Z. Detection of microcystins, a blue-green algal hepatotoxin, in drinking water sampled in Haimen and Fusui, endemic areas of primary liver cancer in China, by highly sensitive immunoassay. Carcinogenesis 1996, 17, 1317–1321. [Google Scholar] [CrossRef] [PubMed]
- Falconer, I.R. An overview of problems caused by toxic blue-green algae (cyanobacteria) in drinking and recreational water. Environ. Toxicol. 1999, 14, 5–12. [Google Scholar] [CrossRef]
- Grosse, Y.; Baan, R.; Straif, K.; Secretan, B.; El Ghissassi, F.; Cogliano, V. Carcinogenicity of nitrate, nitrite, and cyanobacterial peptide toxins. Lancet Oncol. 2006, 7, 628–629. [Google Scholar] [CrossRef] [PubMed]
- Msagati, T.A.M.; Siame, B.A.; Shushu, D.D. Evaluation of methods for the isolation, detection and quantification of cyanobacterial hepatotoxins. Aquat. Toxicol. 2006, 78, 382–397. [Google Scholar] [CrossRef] [PubMed]
- Spoof, L. Microcystins and nodularins. In TOXIC: Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Codd, G.A., Eds.; Åbo Akademi University Press: Turku, Finland, 2005. [Google Scholar]
- Sangolkar, L.N.; Maske, S.S.; Chakrabarti, T. Methods for determining microcystins (peptide hepatotoxins) and microcystin-producing cyanobacteria. Water Res. 2006, 40, 3485–3496. [Google Scholar] [CrossRef] [PubMed]
- Moollan, R.W.; Rae, B.; Verbeek, A. Some comments on the determination of microcystin toxins in waters by high-performance liquid chromatography. Analyst 1996, 121, 233–238. [Google Scholar] [CrossRef]
- Ortea, P.M.; Allis, O.; Healy, B.M.; Lehane, M.; Nı́ Shuilleabháin, A.; Furey, A.; James, K.J. Determination of toxic cyclic heptapeptides by liquid chromatography with detection using ultra-violet, protein phosphatase assay and tandem mass spectrometry. Chemosphere 2004, 55, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Rapala, J.; Erkomaa, K.; Kukkonen, J.; Sivonen, K.; Lahti, K. Detection of microcystins with protein phosphatase inhibition assay, high-performance liquid chromatography-UV detection and enzyme-linked immunosorbent assay: Comparison of methods. Anal. Chim. Acta 2002, 466, 213–231. [Google Scholar] [CrossRef]
- Spoof, L.; Vesterkvist, P.; Lindholm, T.; Meriluoto, J. Screening for cyanobacterial hepatotoxins, microcystins and nodularin in environmental water samples by reversed-phase liquid chromatography-electrospray ionisation mass spectrometry. J. Chromatogr. A 2003, 1020, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Codd, G.A. Microwave oven and boiling waterbath extraction of hepatotoxins from cyanobacterial cells. FEMS Microbiol. Lett. 2000, 184, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Barco, M.; Lawton, L.A.; Rivera, J.; Caixach, J. Optimization of intracellular microcystin extraction for their subsequent analysis by high-performance liquid chromatography. J. Chromatogr. A 2005, 1074, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Zhang, Q.; Jia, C.; Liu, P.; Yang, M. Optimization of microcystin extraction for their subsequent analysis by HPLC-MS/MS method in urban lake water. Int. J. Environ. Sci. Dev. 2013, 4, 600–603. [Google Scholar]
- Li, F.; Liu, W.; Zhao, N.; Duan, J.; Wang, Z.; Zhang, Y.; Xiao, X.; Liu, J.; Yin, G.; Shi, C. Studies on extracting microcystin-LR From Microcystis aeruginosa by water bath. J. Environ. Prot. 2013, 4, 70–73. [Google Scholar] [CrossRef]
- Kim, I.S.; Nguyen, G.H.; Kim, S.Y.; Lee, J.; Yu, H.W. Evaluation of methods for cyanobacterial cell lysis and toxin (microcystin-LR) extraction using chromatographic and mass spectrometric analyses. Environ. Eng. Res. 2009, 14, 250–254. [Google Scholar] [CrossRef]
- Wood, S.A.; Dietrich, D.R.; Cary, S.C.; Hamilton, D.P. Increasing Microcystis cell density enhances microcystin synthesis: A mesocosm study. Inland Waters 2012, 2, 17–22. [Google Scholar] [CrossRef]
- Wood, S.A.; Rueckert, A.; Hamilton, D.P.; Cary, S.C.; Dietrich, D.R. Switching toxin production on and off: Intermittent microcystin synthesis in a Microcystis bloom. Environ. Microbiol. Rep. 2011, 3, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.A.; Giani, A. Cell density-dependent oligopeptide production in cyanobacterial strains. FEMS Microbiol. Ecol. 2014, 88, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Kaufononga, S.A.F.; Cary, S.C.; Hamilton, D.P. High levels of structural diversity observed in microcystins from Microcystis CAWBG11 and characterization of six new microcystin congeners. Mar. Drugs 2014, 12, 5372–5395. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Rhodes, L.L.; Adams, S.L.; Adamson, J.E.; Smith, K.F.; Smith, J.F.; Tervit, H.R.; Cary, S.C. Maintenance of cyanotoxin production by cryopreserved cyanobacteria in the New Zealand culture collection. N. Z. J. Mar. Freshwat. Res. 2008, 42, 277–283. [Google Scholar] [CrossRef]
- Wood, S.A.; Heath, M.W.; Holland, P.T.; Munday, R.; McGregor, G.B.; Ryan, K.G. Identification of a benthic microcystin-producing filamentous cyanobacterium (Oscillatoriales) associated with a dog poisoning in New Zealand. Toxicon 2010, 55, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Carmichael, W.W.; Miller, I. Immuno-gold localization of hepatotoxins in cyanobacterial cells. Arch. Microbiol. 1995, 163, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Young, F.M.; Thomson, C.; Metcalf, J.S.; Lucocq, J.M.; Codd, G.A. Immunogold localisation of microcystins in cryosectioned cells of Microcystis. J. Struct. Biol. 2005, 151, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Zilliges, Y.; Kehr, J.-C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Börner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of Microcystis under oxidative stress conditions. PLoS One 2011, 6, e17615. [Google Scholar] [CrossRef] [PubMed]
- Walsby, A.E. Gas vesicles. Microbiol. Rev. 1994, 58, 94. [Google Scholar]
- Wood, S.A. Personal observation on the difficulty in centrifuging buoyant colonial Microcystis.
- Walsby, A.E. The mechanical properties of the Microcystis gas vesicle. J. Gen. Microbiol. 1991, 137, 2401–2408. [Google Scholar] [CrossRef]
- Harada, K.-I.; Kondo, F.; Lawton, L. Laboratory analysis of cyanotoxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring And Management; Chorus, I., Bartram, J., Eds.; E & FN Spon: London, UK, 1999. [Google Scholar]
- Hyenstrand, P.; Metcalf, J.S.; Beattie, K.A.; Codd, G.A. Effects of adsorption to plastics and solvent conditions in the analysis of the cyanobacterial toxin microcystin-LR by high performance liquid chromatography. Water Res. 2001, 35, 3508–3511. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; Altaner, S.; Kamp, L.; Rubio, F.; Dietrich, D.R. Pitfalls in microcystin extraction and recovery from human blood serum. Chem. Biol. Interact. 2014, 223, 87–94. [Google Scholar] [CrossRef]
- Hyenstrand, P.; Metcalf, J.S.; Beattie, K.A.; Codd, G.A. Losses of the cyanobacterial toxin microcystin-LR from aqueous solution by adsorption during laboratory manipulations. Toxicon 2001, 39, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Bolch, C.; Blackburn, S. Isolation and purification of Australian isolates of the toxic cyanobacterium; Microcystis aeruginosa; Kütz. J. Appl. Phycol. 1996, 8, 5–13. [Google Scholar] [CrossRef]
- Institute for Statistics and Mathematics of Wirtschaftsuniversität Wien. The R Project for Statistical Computing. Available online: http://www.r-project.org/ (accessed on 17 November 2014).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rogers, S.; Puddick, J.; Wood, S.A.; Dietrich, D.R.; Hamilton, D.P.; Prinsep, M.R. The Effect of Cyanobacterial Biomass Enrichment by Centrifugation and GF/C Filtration on Subsequent Microcystin Measurement. Toxins 2015, 7, 821-834. https://doi.org/10.3390/toxins7030821
Rogers S, Puddick J, Wood SA, Dietrich DR, Hamilton DP, Prinsep MR. The Effect of Cyanobacterial Biomass Enrichment by Centrifugation and GF/C Filtration on Subsequent Microcystin Measurement. Toxins. 2015; 7(3):821-834. https://doi.org/10.3390/toxins7030821
Chicago/Turabian StyleRogers, Shelley, Jonathan Puddick, Susanna A. Wood, Daniel R. Dietrich, David P. Hamilton, and Michele R. Prinsep. 2015. "The Effect of Cyanobacterial Biomass Enrichment by Centrifugation and GF/C Filtration on Subsequent Microcystin Measurement" Toxins 7, no. 3: 821-834. https://doi.org/10.3390/toxins7030821
APA StyleRogers, S., Puddick, J., Wood, S. A., Dietrich, D. R., Hamilton, D. P., & Prinsep, M. R. (2015). The Effect of Cyanobacterial Biomass Enrichment by Centrifugation and GF/C Filtration on Subsequent Microcystin Measurement. Toxins, 7(3), 821-834. https://doi.org/10.3390/toxins7030821