1. Introduction
Fungal contamination of foods and feeds causes negative effects on the quality of the products mainly reducing their nutritional and organoleptic properties and lead consequently to important annual economic losses worldwide [
1,
2,
3,
4]. Moreover, fungi are capable of elaborating a wide range of secondary metabolites many of which have been shown to be highly toxic. Therefore, fungal contamination creates a serious health threat for animals as well as for humans.
Aspergillus and its teleomorphs are important mycotoxin producers and these fungi constitute an important contaminant of cereals and feedstuffs [
5,
6,
7,
8]. Especially,
Aspergillus species of the section
Aspergillus—members of the teleomorphic genus
Eurotium in the previous nomenclature [
8]—have been reported to be able to contaminate a variety of biological materials of low water activity. These include stored grains and cereals [
9], dried salted fish [
10], bakery products [
11], mixed feeds and raw materials [
5], cultural assets and old books [
12,
13], and even human corpses [
14].
Aspergillus section
Aspergillus species have also been isolated from hypersaline waters of salterns [
7], air samples near buildings [
15] and from the Dead Sea [
16].
Section
Aspergillus species are generally considered benign fungi for human health and they are used in oriental food fermentation processes as starter cultures [
8]. Species of this section have been isolated and also new species described from natural fermentation products, such as meju, a dried fermented soybean paste typically consumed in Korea [
17,
18,
19]. Although generally considered free of mycotoxins, some of the secondary metabolites produced by the section
Aspergillus fungi are known to show antioxidative, antibacterial, antifungal and antiprotozoal activities [
20,
21,
22]. Some of these compounds, such as echinulin, physcion and flavoglaucin, even though not considered mycotoxins
sensu stricto, do show toxicity to animals [
23,
24,
25,
26]. In addition, the production of more potent mycotoxins such as aflatoxins, gliotoxin, sterigmatocystin and ochratoxin, by the
Aspergillus section
Aspergillus species is controversial [
9,
27,
28,
29,
30,
31,
32,
33]. Several published reports of such mycotoxic potential exist, but these studies have been criticized for lacking further confirming studies using taxonomically properly characterized fungal isolates, or no repetitive mycotoxin detection has been achieved [
9,
27,
28,
29,
30,
31,
32,
33]. Therefore, the true toxicogenic capacity of
Aspergillus section
Aspergillus requires revision. In addition, the growth of xerophilic fungi, like the section
Aspergillus, results in release of metabolic water. This increases the water activity of the contaminated materials in time. Such change in the physico-chemical properties of the substrates can permit the growth of other less xerophilic and highly toxicogenic fungi, such as
Alternaria,
Penicillium and
Fusarium, and consequently result in further contamination of foods and feeds with even more potent mycotoxins.
The mycotoxin-contaminated animal feeds do not have potentially adverse effects only on animal health and productivity but they can cause further secondary contaminations of human consumers via eggs, meat, or milk [
34,
35].
Aspergillus section
Aspergillus teleomorphs have been isolated from rabbit and chinchilla feeds as well as from poultry feeds with very high frequencies in Argentina [
36,
37]. More importantly, these studies have demonstrated that the given animal feeds were also contaminated with various potent mycotoxins (aflatoxins, deoxynivalenol, fumonisins, ochratoxin A, T2-toxin and zearalenone). Even though the detected toxin levels were lower than the regulation limits established, these contaminations with multiple mycotoxins were simultaneous and affected up to 80%–100% of the studied samples. Because the section
Aspergillus fungi can produce multiple secondary metabolites with adverse effects on animal health and also reports of even more hazardous mycotoxins production exist, these high contamination levels are suggesting that both a direct and an indirect risk for animal and human health could be linked to the use of these contaminated animal feeds. This makes the further studies on the degree of xerophilic fungal contamination, the detailed species composition identification and especially, the toxicological characterization of the isolates obtained from animal feeds very valuable. Access to such information would help to evaluate the true mycotoxicological risk generated by the use of feeds in animal production in Argentina. Moreover, fungal strains isolated from different matrices, habitats and geographical origins are expected to have different metabolite profiles and toxicogenic potentials [
38]. Therefore, local information is needed as the results obtained from animal feeds used in other geographical areas cannot be directly extrapolated.
The aim of the work presented here was to study in detail the degree of xerophilic fungal contamination in variable commercial and non-commercial animal feedstuff and primary raw materials destined to rabbit, chinchilla, and rainbow trout production in Argentina. The Aspergillus section Aspergillus species composition was thoroughly characterized and the production of the secondary metabolites was profiled in 20 isolates belonging to the dominant feedstuff contaminating section Aspergillus species. Especially, the evaluation of the secondary metabolite production capacity was expected to clarify the currently controversial position of section Aspergillus species as a toxin free taxon.
The
Aspergillus section
Aspergillus isolates obtained in this study and the previous bibliographic data were treated according to the current nomenclature rules of the Code of Botanical Nomenclature for Algae, Fungi and Plants [
39] and the International Commission on
Penicillium and
Aspergillus (ICPA) [
40]. The isolated teleomorphic fungal states are referred by their genus
Aspergillus species names. This recently proposed change in the fungal nomenclature has also faced criticism as the introduction of the teleomorphic genus
Eurotium into the genus
Aspergillus, with the consequent changes of the species nomenclature, is feared to cause widespread confusion [
41]. To avoid any confusion to the readers, the five
Aspergillus section
Aspergillus species most relevant to this study and supported by the recent taxon revision by Hubka and colleagues [
42] are presented in
Table 1, together with their corresponding former genus
Eurotium names.
Table 1.
The actual nomenclature of five
Aspergillus section
Aspergillus species. The taxa in the teleomophic genus
Eurotium have been transferred to the genus
Aspergillus, according to the one-species-one name principle, and the
Eurotium names should thus not be used anymore. The teleomorph of
A. proliferans was described after this nomenclature change [
42].
Table 1.
The actual nomenclature of five Aspergillus section Aspergillus species. The taxa in the teleomophic genus Eurotium have been transferred to the genus Aspergillus, according to the one-species-one name principle, and the Eurotium names should thus not be used anymore. The teleomorph of A. proliferans was described after this nomenclature change [42].
| Previous Nomenclature | Current Nomenclature |
|---|
| Eurotium amstelodami | Aspergillus montevidensis |
| E. chevalieri | A. chevalieri |
| E. herbariorum | A. glaucus |
| E. repens | A. pseudoglaucus |
| E. rubrum | A. ruber |
| --- | A. proliferans |
3. Discussion
Aspergillus section
Aspergillus species are found in foods and feeds at all a
w levels but they are able to cause spoilage even below 0.90 a
w [
11]. They are also important secondary metabolite producers and, therefore, the section
Aspergillus species, previously known as members of the teleomorphic genus
Eurotium, are considered one of the most destructive xerophilic fungi. Very high isolation frequencies of section
Aspergillus taxa have been previously reported for rabbit, chinchilla and poultry feeds in Argentina. These feeds have also tested positive for multiple very hazardous mycotoxins [
36,
37], suggesting that potential mycotoxicological risks could be linked to their use. Consumption of mycotoxin-contaminated feeds can provoke animal illnesses leading to unnecessary suffering and economic losses. In addition, secondary contaminations of humans as final consumers are possible via consumption of contaminated meat and other animal origin products.
Even though generally considered free of potent mycotoxins, section
Aspergillus fungi are important producers of metabolites with toxic potential [
6,
7]. They can produce a wide variety of secondary metabolites such as echinulin, neoechinulin A, neoechinulin B, preechinulin, cladosporin, questin and physcion [
7,
15,
50,
51]. These compounds have received less attention than more potent mycotoxins, produced predominantly by other fungal species. Therefore, their presence in foods and feeds is not currently controlled or regulated in any country. However, numerous reports exist on the toxic effects of the secondary metabolites produced by section
Aspergillus species on animals over the last 70 years. According to Cole and Cox [
52], some
Aspergillus species (
A. chevalieri and
A. montevidensis) should be considered mycotoxicogenic, and in these cases, echinulin has been the main toxic compound produced. Concordantly, Ali
et al. [
26] have reported liver and lung damages caused by echinulin in rabbits. Furthermore, strains of section
Aspergillus species contaminating cereal grains have been demonstrated to be toxic for experimental animals, causing lowered weight gains in chickens, toxicity to chicken embryos, dermatoxicity in rabbits, hemorrhaging in chickens, hepatotoxicity in mice, and death of calves, rabbits and mice [
53,
54,
55,
56,
57,
58,
59]. In addition, Vesonder
et al. [
59] reported that a feed refused by swine, a situation which led to decreased milk production and consequently to piglets death, was contaminated with echinulin (8 µg/g) and contained high propagule density of
A. montevidensis and
A. chevalieri. These fungal isolates were further confirmed to be able to produce echinulin
in vitro on rice or cracked corn. On the other hand, compounds isolated from
A. montevidensis and
A. chevalieri show different activity against malaria, bacteria and cancer cell lines [
21,
60], metabolites isolated from
A. cristatum have inhibitory activity on tumour cell lines [
61], alkaloids produced by
A. ruber show anti-oxidant activity [
62], and secondary metabolites isolated from
A. pseudoglaucus (physcion and echinuilin) are cytotoxic to sex cells of the sea urchin
Strongylocentrotus [
63].
In the light of such broad evidence of variable toxic effects of secondary metabolites produced by the section Aspergillus, these fungal species can hardly be simply considered benign to animal or human health without further investigation. Variation in the toxicogenic potential between different species and even different isolates are expected to exist. Therefore, in order to evaluate the potential mycotoxicogenical risk linked to section Aspergillus contaminations, further studies on the general fungal abundance, species composition and especially on the secondary metabolite profiles of different species and isolates are needed. Such data are not currently available for section Aspergillus fungi isolated from foods or animal feeds.
With these means, we have conducted a detailed evaluation of the xerophilic mycobiota present in various commercial and non-commercial feed formulations and primary raw materials used for rabbit, chinchilla and rainbow trout production in Argentina. The isolation trials and the growth morphological species identification confirmed the previous reports [
36] of the high contamination frequencies of the rabbit and chinchilla feeds by
Aspergillus section
Aspergillus taxa (>60% of the studied samples), while the rainbow trout feeds showed somewhat lower fungal charge (25%). In addition, some of the primary raw materials tested (pelleted alfalfa, soy, wheat bran, wheat millrun and corn seeds) were positive for section
Aspergillus fungi. These contaminations consisted predominantly of multiple species but the specific species compositions varied both between the feeds and the primary raw material types studied. The presence of five dominant section
Aspergillus species was identified both in the rabbit and chinchilla feeds (
A. montevidensis,
A. chevalieri,
A. pseudoglaucus,
A. ruber and
A. proliferans). However, in chinchilla feed samples
A. montevidensis was the most abundant taxon, while in rabbit feeds three species;
A. montevidensis,
A. chevalieri and
A. pseudoglaucus were the most frequently isolated ones, led by
A. pseudoglaucus. On the other hand, rainbow trout feeds showed a completely different species composition formed only by two species,
A. pseudoglaucus and
A. proliferans, both with quite similar isolation frequencies. These variations in species abundance and composition most probably reflect different natures and complexities of the primary raw materials used for manufacturing different animal feed formulations, even though possible impacts of post-fabrication storage conditions cannot be excluded. This conclusion was supported further by the isolation results from the primary raw materials studied. When tested positive, fungal contamination was by a single or maximum two section
Aspergillus species and species compositions were clearly variable between the different types of primary raw material analysed.
The correct and solid species identification is the base of any biological study. The taxonomy and nomenclature of
Aspergillus section
Aspergillus species has been under revision during the last years. The transfer of the teleomorphic genus
Eurotium to the anamorphic
Aspergillus genus, according to the one-species-one scientific name concept of the botanical code, has led to re-evaluation of the previous data on linkage of anamorphic and teleomorphic fungal states and the species identity. Unfortunate misidentifications of culture collection deposited isolates, contradictory posterior species descriptions and the use of various synonymous species names during the years has seriously complicated this task. However, recently, DNA analyses, together with physiological and growth characters, have been used for clarifying the true taxonomic species position and to solidly link the teleomorphs with their anamorphic states. The studies of Peterson [
64] and Hubka
et al. [
42] have profoundly revised the previous species nomenclature and the identity of taxa in the section
Aspergillus. The growth morphological characters and light microscopical observation of the ascospores, traditionally used for section
Aspergillus species identification, are to some extent error prone due to their subjectivity. Currently, access to vast molecular taxonomic data from type-strains of section
Aspergillus species allows a fast and precise direct isolate identification or growth morphological species identity confirmations.
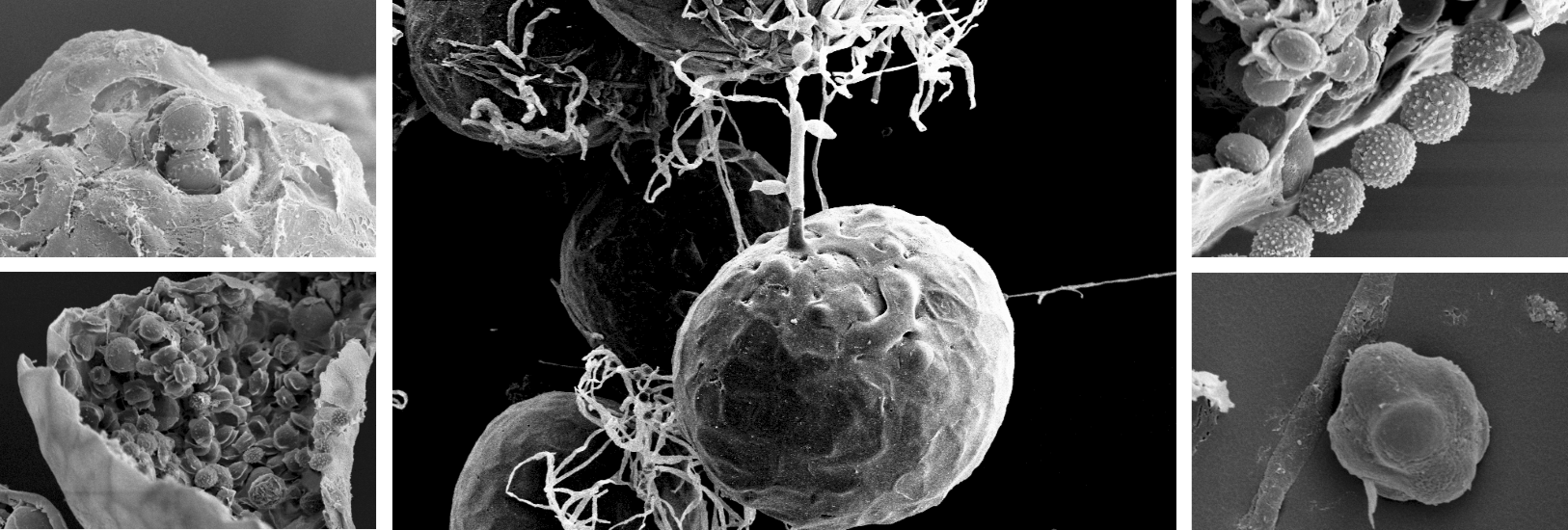
The taxonomic position of the 20 isolates, representatives of the five dominant feedstuff contaminant species of section
Aspergillus: A. montevidensis,
A. chevalieri,
A. proliferans,
A. pseudoglaucus and
A. ruber, identified by their growth morphological characters, was confirmed further using both scanning electron microscopy (SEM) and two independent DNA loci. Scanning electron microscopy allows detailed observation and description of ascospore size and structural characteristics, such as surface ornamentation, pores and their arrangement, longitudinal groove and ridges. These have served and still do serve as important characteristics for species identification in the section
Aspergillus [
8,
17,
42,
47,
65,
66]. However, the ascospore characteristics can also be highly similar between different species, like is the case with
A. ruber and
A. proliferans. In addition, a relative high intraspecific and even intraisolate variation of the ascospore characteristics can exist. Therefore, SEM as a species classification and identification tool has a supplementary value, but it is not conclusive. This was confirmed by our SEM study as it allowed to correct some erroneous growth morphological species identifications but did not offer separation between
A. ruber and
A. proliferans isolates.
The conclusive confirmation species identity of the isolates was reached with the phylogenetic analysis based on the beta-tubulin gene fragment. The nuclear ITS region has been established as a universal DNA barcode marker for fungi [
67]. However, this marker does not necessary have a species level resolution in all fungal taxa. Specifically in the genus
Aspergillus, identical ITS sequences are known to be shared between several complexes. Therefore, other general protein encoding DNA loci (such as beta-tubulin, calmodulin or RNA polymerase subunit 2) are frequently used for species identification [
40,
42,
64,
68]. The degree of ITS species resolution can also vary between different sections of
Aspergillus. For example, a phylogenetic study based on ITS sequences of the
Aspergillus section
Circumdati resulted in identification of 18 out of 27 species [
69]. In our phylogenetic analyses of 18 section
Aspergillus species, the ITS did not allow species level identification for any of them. On the other hand, the analysis based on the beta-tubulin gene fragment led to species level identification for 16 out of 18 of the reference taxa and of all the studied isolates. These results are concordant with reports of beta-tubulin locus as a very potent single DNA marker for species level resolution in
Aspergillus section
Aspergillus [
17,
42,
66,
70].
High isolation frequencies of section
Aspergillus taxa were obtained from the animal feeds analysed in the present study similar to earlier reports from Argentina [
36,
37]. This raises the question of the already existing or potential contamination of such feeds with multiple bioactive and toxic secondary metabolites known to be produced by the section
Aspergillus fungi. In addition, the evaluation of the potential toxicological risks requires previous knowledge on the toxicogenic capacity of the section
Aspergillus species contaminating feeds. No previous information exist on secondary metabolite profiles of the section
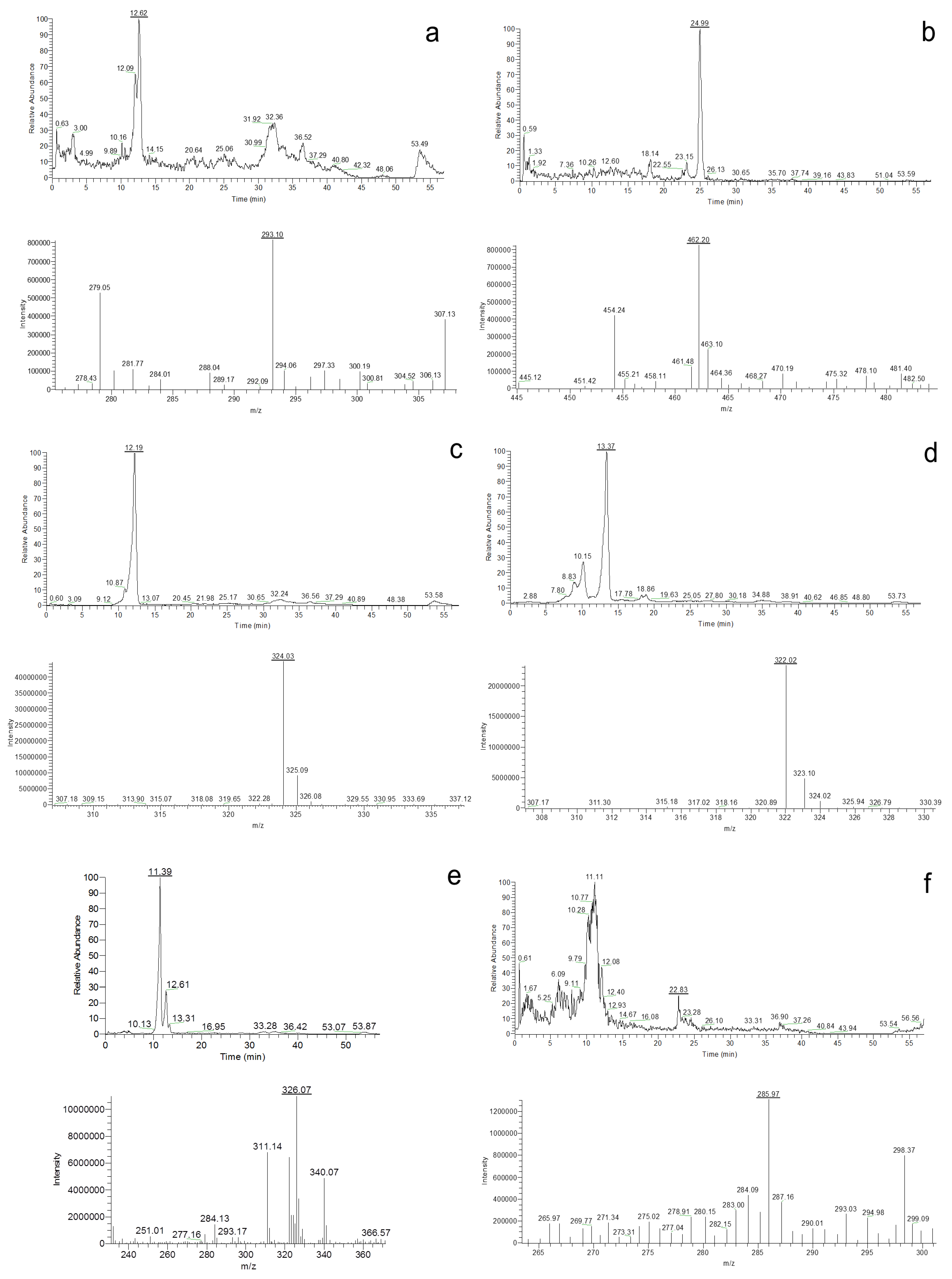
Aspergillus species isolated from animal feeds in Argentina and, to our knowledge, from any other country in the world. The secondary metabolite profiling of the 20 isolates belonging to the five most predominant section
Aspergillus species contaminating different animal feeds revealed that, on the one hand, the five species produce a shared common metabolite profile. All the species were able to produce, under the experimental culture conditions used, cladosporin, echinulin, and neoechinulin A and B. However, variations in the secondary metabolite profile exist between the isolates of the same species. Only in the case of
A. montevidensis the three isolates tested were all positive for these four secondary metabolites. In addition, preechinulin production was detected in one
A. montevidensis and
A. chevalieri isolates and physcion in two
A. chevalieri isolates.
Production of auroglaucins, flavoglaucins and anthraquinones, such as physcion, catenarin, questin and questinol, has been reported before for
A. montevidensis,
A. chevalieri,
A. pseudoglaucus. A. ruber,
A. glaucus and
A. proliferans [
7]. None of these compounds, except physcion, was detected in the studied isolates. In addition,
A. montevidensis and
A. ruber isolates have also been reported to produce epiheveadride [
15]. We did not detect this secondary metabolite in any of the studied isolates. On the other hand, all three of our
A. montevidensis isolates and two of four
A. ruber isolates tested positive for cladosporin, a compound that was not detected in the given species by Slack
et al. [
15].
Our secondary metabolite profiling results demonstrate that the
Aspergillus section
Aspergillus species isolated from different matrices and geographical origins do show different toxinogenic potential. All the five main species contaminating the animal feeds under study in Argentina had a consistent but, at the same time, rather limited secondary metabolite profile, at least under the culture conditions tested. No significant variation exists between species, though minor intraspecies isolate variations were detected. However, all the isolates were capable of producing echinulin, the precise secondary metabolite demonstrated to have serious mycotoxic effects on pigs, mice, rabbits [
26,
52]. The highest toxinogenic potential was detected in
A. chevalieri as this species, depending on the isolate, tested positive for altogether six secondary metabolites.
This is the first report on detailed taxonomic identification and secondary metabolite profiling of
Aspergillus section
Aspergillus fungi contaminating feeds and primary raw materials used for animal feeding in Argentina. So far, most studies have observed additive or synergistic effects between consumption of different potent mycotoxins [
71,
72,
73]. However, the effects of chronic simultaneous consumption of sub-toxic concentrations of fungal metabolites of lower toxicogenic potential, like the ones produced by the section
Aspergillus, on animal or human health are currently poorly studied. Neither it is known if these compounds show additive or synergistic effects with each other or with other more hazardous mycotoxins. Therefore, further studies are needed to determine the full toxicogenic potential of
Aspergillus section
Aspergillus species under variable growth conditions to assess mycotoxicological risk that these xerophilic fungi can generate for animal and human health as feed and food contaminants.




{kind=link}
{kind=link}
{kind=link}
{kind=link}


