
Growth-Phase Sterigmatocystin Formation on Lactose Is Mediated via Low Specific Growth Rates in Aspergillus nidulans


 and
and
Abstract
:1. Introduction
2. Results
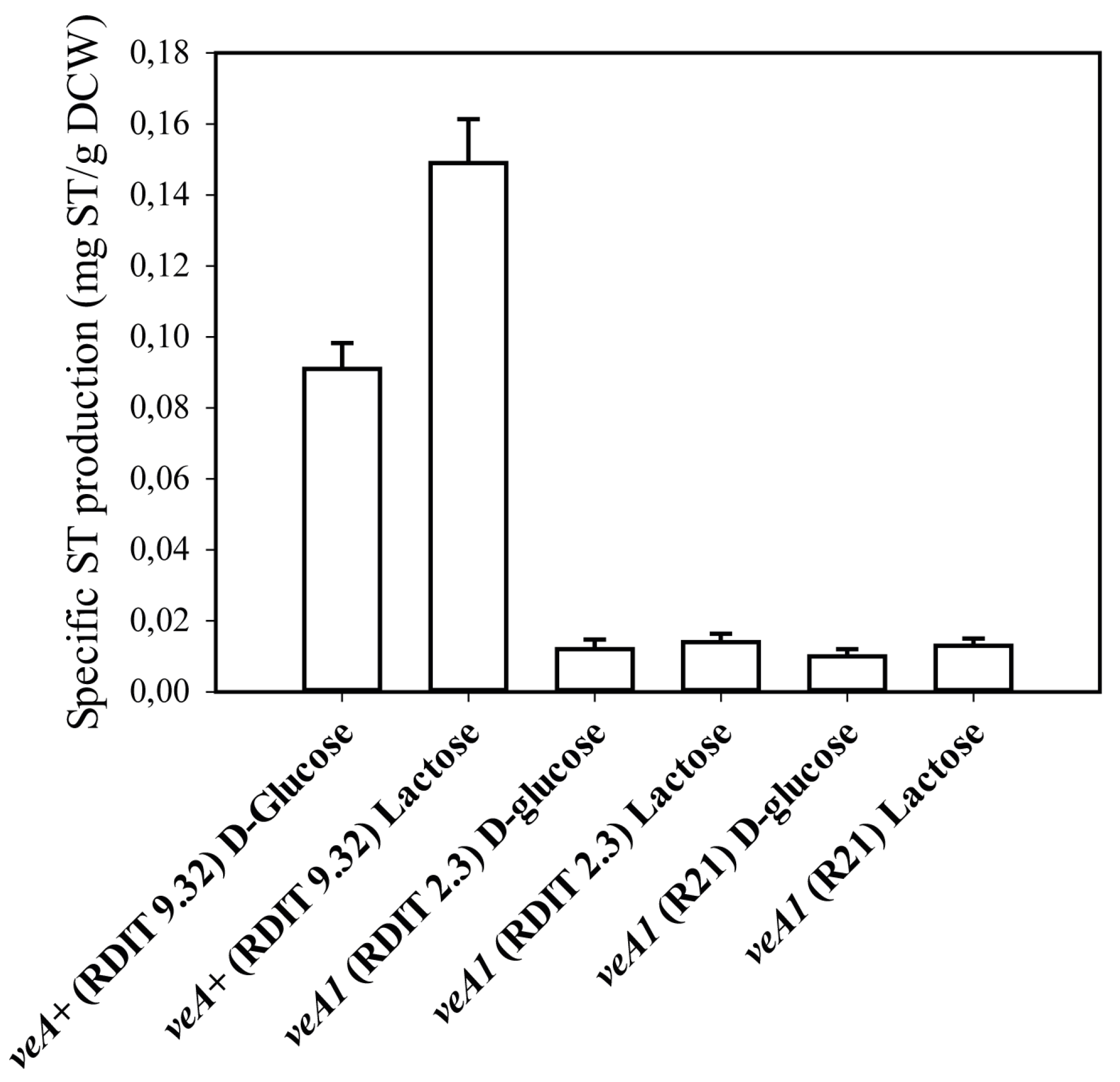
2.1. Verification of the Experimental Strategy
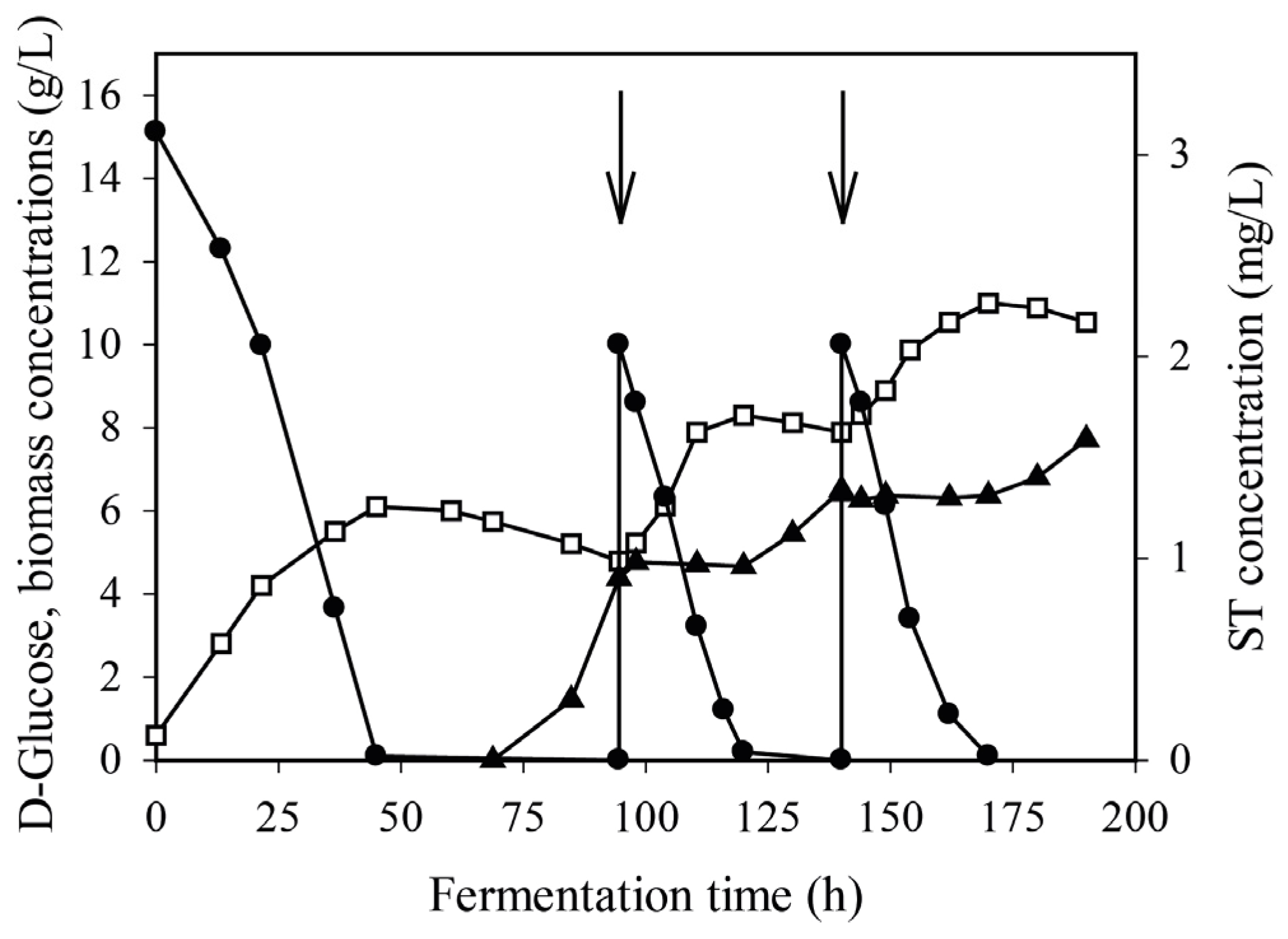
2.2. Kinetics of the Wild-Type Sterigmatocystin Production in Batch and Fed-Batch Cultures
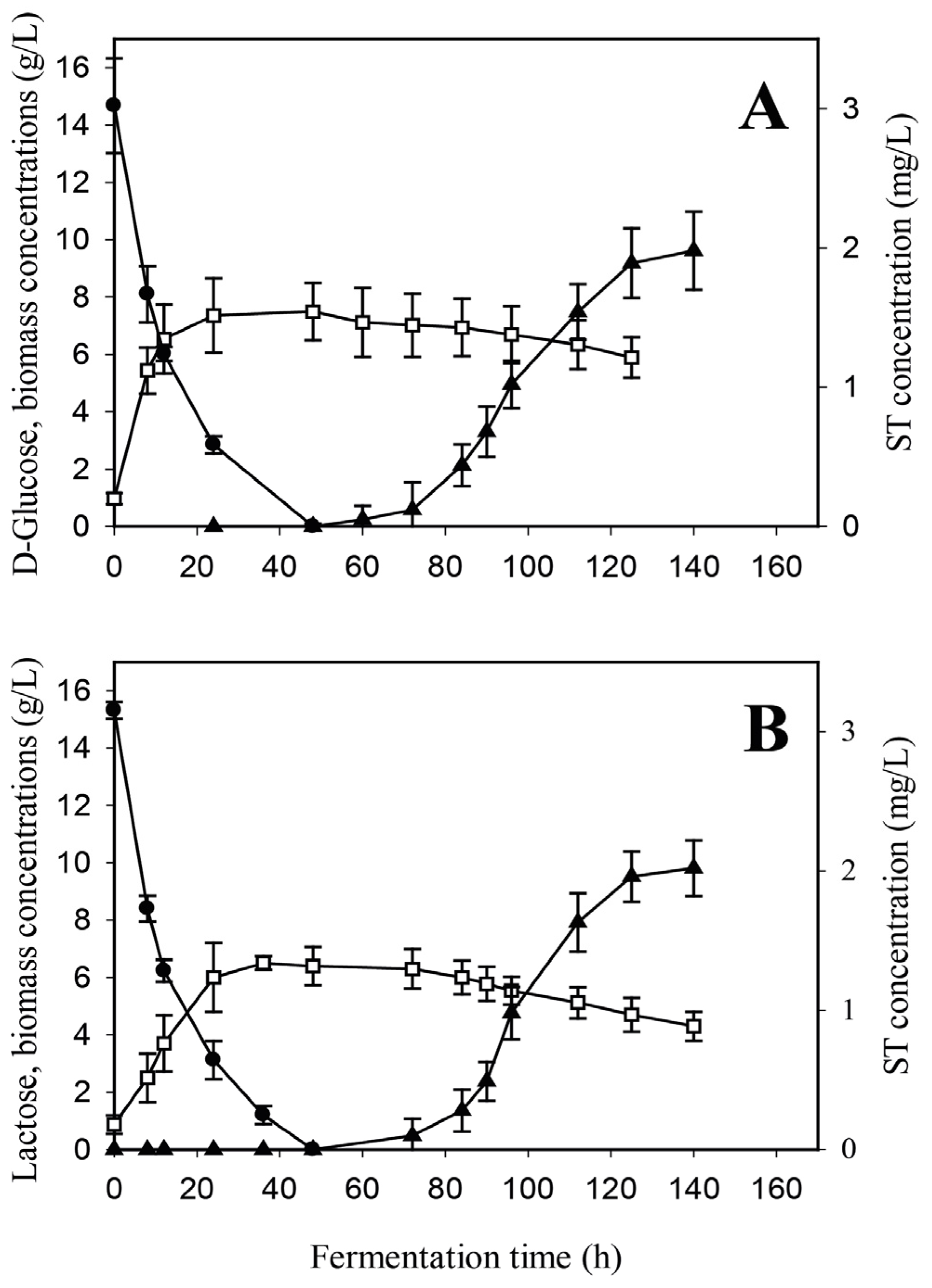
2.3. Kinetics of Sterigmatocystin Production by A. nidulans in a creA-Negative Background
2.4. Sterigmatocystin Formation of A. nidulans in Chemostat-Type Continuous Cultures
3. Discussion
4. Materials and Methods
4.1. Strains and Cultivation Conditions
4.2. Analytical Methods
4.3. Genomic DNA Isolation
4.4. Reproducibility
4.5. Chemicals
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rank, C.; Nielsen, K.F.; Larsen, T.O.; Varga, J.; Samson, R.B.A.; Frisvad, J.S. Distribution of sterigmatocystin in filamentous fungi. Fungal Biol. 2011, 115, 406–420. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, H.H.; Ramaswamy, A.; Sung, C.S.; Keller, N.P. Increased conidiation associated with progression along the sterigmatocystin biosynthetic pathway. Mycology 2004, 96, 1190–1198. [Google Scholar] [CrossRef]
- Wogan, G.N. Aflatoxins as risk for hepatocellular carcinoma in humans. Cancer Res. 1992, 52, 2114–2118. [Google Scholar]
- Hopwood, D.A.; Sherman, D.H. Molecular genetics of polyketides and its comparison to fatty acid biosynthesis. Annu. Rev. Genet. 1990, 24, 37–66. [Google Scholar] [CrossRef] [PubMed]
- Purchase, I.F.H.; van der Watt, J.J. Carcinogenicity of sterigmatocystin. Food Cosmet. Toxicol. 1970, 8, 291–295. [Google Scholar] [CrossRef]
- Barnes, S.E.; Dola, T.P.; Bennett, J.W.; Bhatnagar, D. Synthesis of sterigmatocystin on a chemically defined medium by species of Aspergillus and Chaetomium. Mycopathologia 1994, 125, 173–178. [Google Scholar] [CrossRef]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Albright, J.C.; Ye, R.; Mead, D.; Wagner, M.; Krerowicz, A.; Goering, A.W.; Kelleher, N.L.; Keller, N.P.; Wu, C.-C. Fungal artificial chromosomes for mining of the fungal secondary metabolome. BMC Genom. 2015, 16, 343. [Google Scholar] [CrossRef] [PubMed]
- Hodges, R.L.; Hodges, D.W.; Goggans, K.; Xuei, X.; Skatrud, P.; McGilvray, D. Genetic modification of an echinocandin B-producing strain of Aspergillus nidulans to produce mutants blocked in sterigmatocystin biosynthesis. J. Ind. Microbiol. 1994, 13, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Penalva, N.A.; Rowlands, R.T.; Turner, G. The optimization of penicillin biosynthesis in fungi. Trends Biotechnol. 1998, 16, 483–489. [Google Scholar] [CrossRef]
- Kennedy, J.; Auclair, K.; Kendrew, S.G.; Park, C.; Vederas, J.C.; Hutchinson, C.R. Modulation of polyketide synthase activity by accessory proteins during lovastatin biosynthesis. Science 1999, 284, 1368–1372. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B.G.; Mnich, E.; Nielsen, K.F.; Nielsen, J.B.; Nielsen, M.T.; Mortensen, U.H.; Larsen, T.O.; Patil, K.R. Involvement of a natural fusion of a cytochrome P450 and a hydrolase in mycophenolic acid biosynthesis. Appl. Environ. Microbiol. 2012, 78, 4908–4913. [Google Scholar] [CrossRef] [PubMed]
- Neely, F.L.; Emerson, C.S. Determination of sterigmatocystin in fermentation broths by reversed-phase high-performance liquid chromatography using post-column fluorescence enhancement. J. Chromatogr. 1990, 523, 305–311. [Google Scholar] [CrossRef]
- Hodges, R.L.; Kelkar, H.S.; Xuei, X.; Skatrud, P.L.; Keller, N.P.; Adams, T.H.; Kaiser, R.E.; Vinci, V.A.; McGilvray, D. Characterization of an echinocandin B-producing strain blocked for sterigmatocystin biosynthesis reveals a translocation in the stcW gene of the aflatoxin biosynthetic pathway. J. Ind. Microbiol. 2001, 25, 333–341. [Google Scholar] [CrossRef]
- Maggio-Hall, L.A.; Wilson, R.A.; Keller, N.P. Fundamental contribution of β-oxidation to polyketide mycotoxin production in planta. Mol. Plant Microbe Interact. 2005, 18, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Tsitsigiannis, D.I.; Kowieski, T.M.; Zarnowski, R.; Keller, N.P. Three putative oxylipin biosynthetic genes integrate sexual and asexual development in Aspergillus nidulans. Microbiology 2005, 151, 1809–1821. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M. The VeA regulatory system and its role in morphological and chemical development in fungi. Fungal Genet. Biol. 2008, 45, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Stinnett, S.M.; Espeso, E.A.; Cobeño, L.; Araújo-Bazán, L.; Calvo, A.M. Aspergillus nidulans VeA subcellular localization is dependent on the importin α carrier and on light. Mol. Microbiol. 2007, 63, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Septien, I.; Blanco, J.L.; Suarez, G.; Cutuli, M.T. Solubility and stability of sterigmatocystin in aqueous solution. Mycotoxin Res. 1994, 10, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.B.; Smith, J.E. Degradation of aflatoxin by Aspergillus flavus. J. Gen. Microbiol. 1987, 133, 2023–2029. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Jezkova, A.; Yuan, Z.; Pavlikova, L.; Dohnal, V.; Kuca, K. Biological degradation of aflatoxins. Drug Metab. Rev. 2009, 41, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ilyés, H.; Fekete, E.; Karaffa, L.; Fekete, É.; Sándor, E.; Szentirmai, A.; Kubicek, C.P. CreA-mediated carbon catabolite repression of β-galactosidase formation in Aspergillus nidulans is growth rate dependent. FEMS Microbiol. Lett. 2004, 235, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Brown, A. Fed-batch and continuous culture. In Fermentation: A Practical Approach; McNeil, B., Harvey, L.M., Eds.; Oxford University Press: Oxford, UK, 1990; pp. 113–130. [Google Scholar]
- Jorgensen, T.R.; van Kuyk, P.A.; Poulsen, B.R.; Ruijter, G.J.G.; Visser, J.; Iversen, J.J.L. Glucose uptake and growth of glucose-limited chemostat cultures of Aspergills niger and a disruptant lacking MstA, a high-affinity glucose transporter. Microbiology 2007, 153, 1963–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdollahi, A.; Buchanan, R.L. Regulation of aflatoxin biosynthesis: Induction of aflatoxin production by various carbohydrates. J. Food Sci. 1981, 46, 633–635. [Google Scholar] [CrossRef]
- Keller, N.P.; Nesbitt, C.; Sarr, B.; Philips, T.D.; Burow, G.B. pH regulation of sterigmatocystin and aflatoxin biosynthesis in Aspergillus spp. Phytopathology 1997, 87, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Price, M.S.; Conners, S.B.; Tachdjian, S.; Kelly, R.B.; Payne, G.A. Aflatoxin conducive and non-conducive growth conditions reveal new gene associations with aflatoxin production. Fungal Genet. Biol. 2005, 42, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Atoui, A.; Kastner, C.; Larey, C.M.; Thokala, R.; Etxebeste, O.; Espeso, E.A.; Fischer, R.; Calvo, A.M. Cross-talk between light and glucose regulation controls toxin production and morphogenesis in Aspergillus nidulans. Fungal Genet. Biol. 2010, 47, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Hicks, J.K.; Yu, J.H.; Keller, N.P.; Adams, T.H. Aspergillus sporulation and mycotoxin production both require inactivation of the FadA Gα protein-dependent signaling pathway. EMBO J. 1997, 16, 4916–4923. [Google Scholar] [CrossRef] [PubMed]
- Aharonowitz, Y.; Cohen, G.; Martin, J.F. Penicillin and cephalosporin biosynthetic genes: Structure, organization, regulation and evolution. Annu. Rev. Microbiol. 1992, 46, 461–495. [Google Scholar] [CrossRef] [PubMed]
- Brakhage, A.A. Molecular regulation of β-lactam biosynthesis in filamentous fungi. Microbiol. Mol. Biol. Rev. 1998, 62, 547–585. [Google Scholar] [PubMed]
- Espeso, E.A.; Peñalva, M.A. Carbon catabolite repression can account for the temporal pattern of expression of a penicillin biosynthetic gene in Aspergillus nidulans. Mol. Microbiol. 1992, 6, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhu, Y.; Bao, L.; Gao, L.; Yao, G.; Li, Y.; Yang, Z.; Li, Z.; Zhong, Y.; Li, F.; et al. Putative methyltransferase LaeA and transcription factor CreA are necessary for proper asexual development and controlling secondary metabolic gene cluster expression. Fungal Genet. Biol. 2016, 94, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Karaffa, L.; Fekete, E.; Gamauf, C.; Szentirmai, A.; Kubicek, C.P.; Seiboth, B. d-galactose induces cellulase gene expression in Hypocrea jecorina at low growth rates. Microbiology 2006, 152, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Portnoy, T.; Margeot, A.; Linke, R.; Atanasova, L.; Fekete, E.; Sándor, E.; Hartl, L.; Karaffa, L.; Druzhinina, I.S.; Seiboth, B.; et al. The CRE1 carbon catabolite repressor of the fungus Trichoderma reesei: A master regulator of carbon assimilation. BMC Genom. 2011, 12, 269. [Google Scholar] [CrossRef] [PubMed]
- Hicks, J.K.; Lockington, R.A.; Strauss, J.; Dieringer, D.; Kubicek, C.P.; Kelly, J.; Keller, N.P. RcoA has pleiotropic effects on Aspergillus nidulans cellular development. Mol. Microbiol. 2001, 39, 1482–1493. [Google Scholar] [CrossRef] [PubMed]
- Fekete, E.; Karaffa, L.; Seiboth, B.; Fekete, É.; Kubicek, C.P.; Flipphi, M. Identification of a permease gene involved in lactose utilisation in Aspergillus nidulans. Fungal Genet. Biol. 2012, 49, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Shih, C.N.; Marth, E.H. Some cultural conditions that control biosynthesis of lipid and aflatoxin by Aspergillus parasiticus. Appl. Microbiol. 1974, 27, 452–456. [Google Scholar] [PubMed]
- Fekete, E.; Orosz, A.; Kulcsár, L.; Kavalecz, N.; Flipphi, M.; Karaffa, L. Characterization of a second physiologically relevant lactose permease gene (lacpB) in Aspergillus nidulans. Microbiology 2016, 162, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Rebnegger, C.; Graf, A.B.; Valli, M.; Steiger, M.G.; Gasser, B.; Maurer, M.; Mattanovich, D. In Pichia pastoris, growth rate regulates protein synthesis and secretion, mating and stress response. Biotechnol. J. 2014, 9, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Barad, S.; Chen, Y.; Luo, X.; Tannous, J.; Dubey, A.; Glam, N.; Tian, S.; Li, B.; Keller, N.P.; et al. LaeA regulation of secondary metabolism modulates virulence in Penicillium expansum and is mediated by sucrose. Mol. Plant Pathol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Hanif, M.; Khattak, M.K.; Rehman, M.U.; Ramzan, M.; Amin, M.; Aamir, M.; Sher, S.S.; Hafizullah; Khan, S.; Saeed, M.; et al. Effect of drying temperature and natural preservatives on reducing aflatoxins in solar dried persimmon (Diospyros kaki L). Proc. Pak. Acad. Sci. 2015, 52, 361–365. [Google Scholar]
- Wellford, T.E.T.; Eadie, T.; Llewellyn, G.C. Evaluating, the inhibitory action of honey on fungal growth, sporulation, and aflatoxin production. Z. Lebensm. Unters. Forsch. 1978, 166, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Fekete, E.; Karaffa, L.; Sándor, E.; Seiboth, B.; Bíró, S.; Szentirmai, A.; Kubicek, C.P. Regulation of the intracellular β-galactosidase activity of Aspergillus nidulans. Arch. Microbiol. 2002, 179, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Clutterbuck, A.J. Aspergillus nidulans. In Handbook of Genetics, Bacteria, Bacteriophages, and Fungi; King, R.C., Ed.; Plenum Press: New York, NY, USA, 1974; Volume 1, pp. 447–510. [Google Scholar]
- Han, K.H.; Park, J.S.; Chae, K.S.; Han, D.M. Simple identification of veA1 mutation in Aspergillus nidulans. J. Microbiol. 2010, 48, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Shroff, R.A.; O’Connor, S.M.; Hynes, M.J.; Lockington, R.A.; Kelly, J.M. Null alleles of creA, the regulator of carbon catabolite repression in Aspergillus nidulans. Fungal Genet. Biol. 1997, 22, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Tsitsigiannis, D.I.; Zarnowski, R.; Keller, N.P. The lipid body protein, PpoA, coordinates sexual and asexual sporulation in Aspergillus nidulans. J. Biol. Chem. 2004, 279, 11344–11353. [Google Scholar] [CrossRef] [PubMed]
- Fantes, P.A.; Roberts, C.F. β-Galactosidase activity and lactose utilization in Aspergillus nidulans. J. Gen. Microbiol. 1973, 77, 471–486. [Google Scholar] [CrossRef]
- Pontecorvo, G.; Roper, J.A.; Chemmons, L.M.; Macdonald, K.D.; Bufton, A.W.J. The genetics of Aspergillus nidulans. Adv. Genet. 1953, 5, 141–238. [Google Scholar] [PubMed]
- Bayram, Ö.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain, Carbon Source | Carbon Utilization Rate (g/h) | Biomass Production Rate (gDCW/h) |
|---|---|---|
| Wild-type, d-glucose | 0.30 ± 0.03 | 0.15 ± 0.02 |
| Wild-type, lactose | 0.14 ± 0.02 | 0.07 ± 0.02 |
| CreA mutant, d-glucose | 0.29 ± 0.03 | 0.14 ± 0.01 |
| CreA mutant, lactose | 0.27 ± 0.03 | 0.13 ± 0.02 |
| Strain | Dilution Rate (1/h) | Steady-State Sterigmatocystin Concentration (mg/L) |
|---|---|---|
| Wild-type (veA+) | 0.09 | <0.1 |
| 0.02 | 0.5 ± 0.07 | |
| CreA mutant (veA+) | 0.09 | <0.1 |
| 0.02 | 0.4 ± 0.05 |
| Strain | Genotype | Reference |
|---|---|---|
| RDIT 9.32 | veA+ | Tsitsigiannis et al. [49] |
| (FGSC #A1252) | ||
| RDIT 2.3 | veA1 | Tsitsigiannis et al. [49] |
| (FGSC #A1830) | ||
| R21 | yA2, pabaA1; veA1 | Fantes and Roberts [50] |
| (FGSC #A1228) | ||
| V100 1 | creAΔ4; pantoB100; veA1 | Shroff et al. [48] 2 |
| RJMP 155.55 | wA3; riboB2, veA+ | Keller Lab (unpublished) |
| ZNEF 8.55 3 | creAΔ4; wA3; pantoB100; riboB2, veA+ | This study |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Németh, Z.; Molnár, Á.P.; Fejes, B.; Novák, L.; Karaffa, L.; Keller, N.P.; Fekete, E. Growth-Phase Sterigmatocystin Formation on Lactose Is Mediated via Low Specific Growth Rates in Aspergillus nidulans. Toxins 2016, 8, 354. https://doi.org/10.3390/toxins8120354
Németh Z, Molnár ÁP, Fejes B, Novák L, Karaffa L, Keller NP, Fekete E. Growth-Phase Sterigmatocystin Formation on Lactose Is Mediated via Low Specific Growth Rates in Aspergillus nidulans. Toxins. 2016; 8(12):354. https://doi.org/10.3390/toxins8120354
Chicago/Turabian StyleNémeth, Zoltán, Ákos P. Molnár, Balázs Fejes, Levente Novák, Levente Karaffa, Nancy P. Keller, and Erzsébet Fekete. 2016. "Growth-Phase Sterigmatocystin Formation on Lactose Is Mediated via Low Specific Growth Rates in Aspergillus nidulans" Toxins 8, no. 12: 354. https://doi.org/10.3390/toxins8120354
APA StyleNémeth, Z., Molnár, Á. P., Fejes, B., Novák, L., Karaffa, L., Keller, N. P., & Fekete, E. (2016). Growth-Phase Sterigmatocystin Formation on Lactose Is Mediated via Low Specific Growth Rates in Aspergillus nidulans. Toxins, 8(12), 354. https://doi.org/10.3390/toxins8120354