
Heterologous Expression of Toxins from Bacterial Toxin-Antitoxin Systems in Eukaryotic Cells: Strategies and Applications

 ,
, 
{kind=link}
{kind=link}
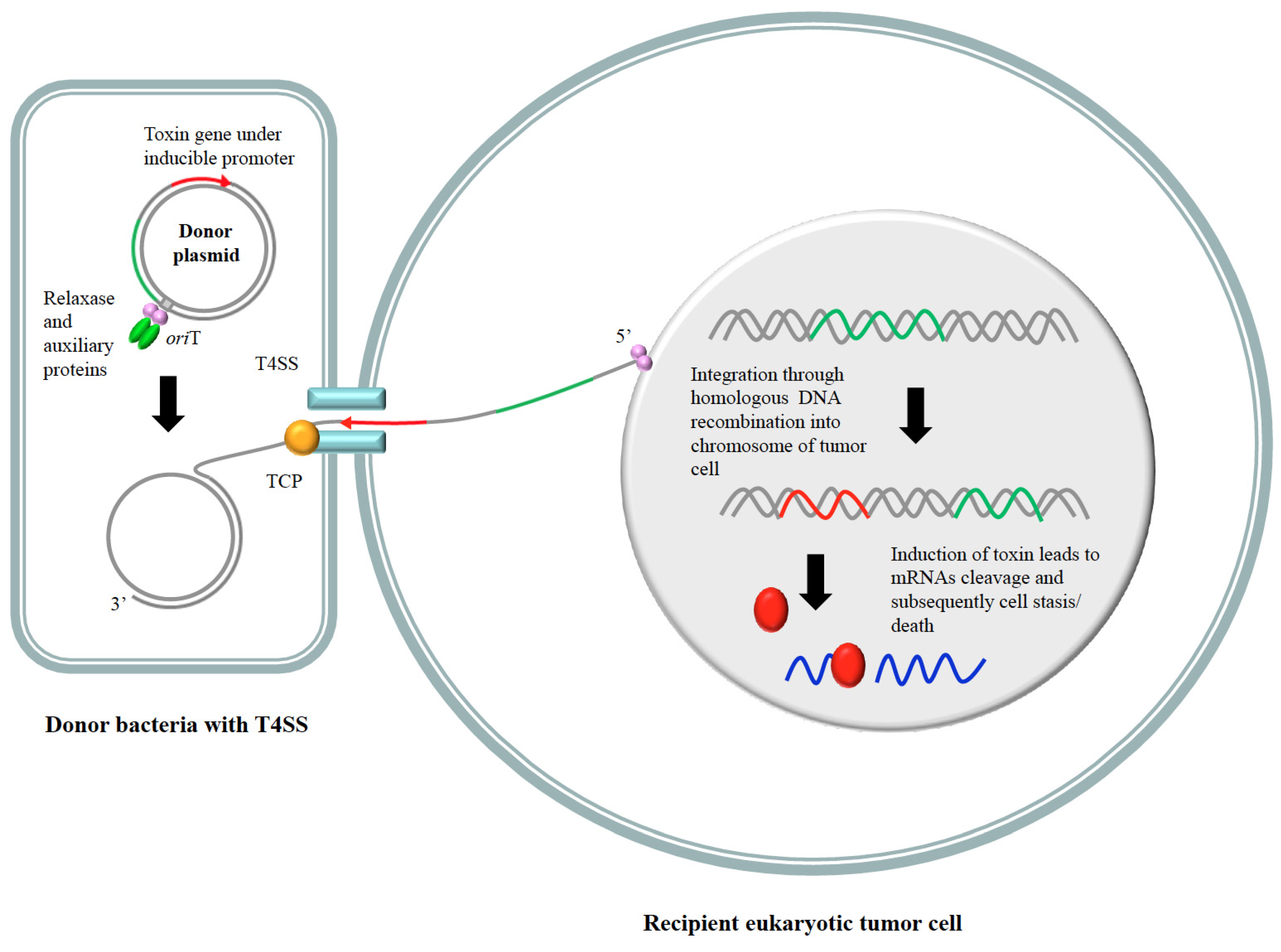{kind=link}
Abstract
:1. Introduction: An Overview of Bacterial Toxin-Antitoxin Systems
2. Expression of TA Systems in Yeasts: Applications
2.1. TA Systems as Tools for Containment in Yeasts
2.2. TA Systems as Tools for the Genetic Manipulation of Yeasts
3. TA Systems as Cell Ablation Tools
3.1. Cell Ablation for the Containment of Genetically Modified Plants
3.2. Cell Ablation in Developmental Biology Research of Higher Eukaryotes
4. TA Systems as Tools for Overproduction of Heterologous Proteins in Eukaryotic Cells
5. TA Systems in Gene Therapy
5.1. Antiviral Gene Therapy
5.2. Anticancer Gene Therapy
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CMV | cytomegalovirus |
| GMO | genetically-modified organism |
| HCV | hepatitis C virus |
| HIV | human immunodeficiency virus |
| IRES | internal ribosome entry site |
| TA | toxin-antitoxin |
| T4SS | type IV secretion system |
| UNAG | uridine diphosphate-N-acetylglucosamine |
References
- Gerdes, K.; Christensen, S.K.; Løbner-Olesen, A. Prokaryotic toxin-antitoxin stress response loci. Nat. Rev. Microbiol. 2005, 3, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Hayes, F.; van Melderen, L. Toxins-antitoxins: Diversity, evolution and function. Crit. Rev. Biochem. Mol. Biol. 2011, 46, 386–408. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Park, J.-H.; Inouye, M. Toxin-antitoxin systems in bacteria and archaea. Annu. Rev. Genet. 2011, 45, 61–79. [Google Scholar] [CrossRef] [PubMed]
- Goeders, N.; van Melderen, L. Toxin-antitoxin systems as multilevel interaction systems. Toxins 2014, 6, 304–324. [Google Scholar] [CrossRef] [PubMed]
- Hayes, F.; Kędzierska, B. Regulating toxin-antitoxin expression: Controlled detonation of intracellular molecular timebombs. Toxins 2014, 6, 337–358. [Google Scholar] [CrossRef] [PubMed]
- Yarmolinsky, M.B. Programmed cell death in bacterial populations. Science 1995, 267, 836–837. [Google Scholar] [CrossRef] [PubMed]
- Hayes, F. Toxins-antitoxins: Plasmid maintenance, programmed cell death, and cell cycle arrest. Science 2003, 301, 1496–1499. [Google Scholar] [CrossRef] [PubMed]
- Aizenman, E.; Engelberg-Kulka, H.; Glaser, G. An Escherichia coli chromosomal “addiction module” regulated by guanosine 3′,5′-bispyrophosphate: A model for programmed bacterial cell death. Proc. Natl. Acad. Sci. USA 1996, 93, 6059–6063. [Google Scholar] [CrossRef] [PubMed]
- Engelberg-Kulka, H.; Glaser, G. Addiction modules and programmed cell death and antideath in bacterial cultures. Annu. Rev. Microbiol. 1999, 53, 43–70. [Google Scholar] [CrossRef] [PubMed]
- Engelberg-Kulka, H.; Amitai, S.; Kolodkin-Gal, I.; Hazan, R. Bacterial programmed cell death and multicellular behavior in bacteria. PLoS Genet. 2006, 2. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.; Christensen, S.K.; Gerdes, K. Rapid induction and reversal of a bacteriostatic condition by controlled expression of toxins and antitoxins. Mol. Microbiol. 2002, 45, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.K.; Mikkelsen, M.; Pedersen, K.; Gerdes, K. RelE, a global inhibitor of translation, is activated during nutritional stress. Proc. Natl. Acad. Sci. USA 2001, 98, 14328–14333. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.K.; Gerdes, K. RelE toxins from bacteria and Archaea cleave mRNAs on translating ribosomes, which are rescued by tmRNA. Mol. Microbiol. 2003, 48, 1389–1400. [Google Scholar] [CrossRef] [PubMed]
- Keren, I.; Shah, D.; Spoering, A.; Kaldalu, N.; Lewis, K. Specialized persister cells and the mechanism of multidrug tolerance in Escherichia coli. J. Bacteriol. 2004, 186, 8172–8180. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef] [PubMed]
- De Bast, M.S.; Mine, N.; van Melderen, L. Chromosomal toxin-antitoxin systems may act as antiaddiction modules. J. Bacteriol. 2008, 190, 4603–4809. [Google Scholar] [CrossRef] [PubMed]
- Hazan, R.; Engelberg-Kulka, H. Escherichia coli mazEF-mediated cell death as a defense mechanism that inhibits the spread of phage P1. Mol. Genet. Genomics 2004, 272, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Blower, T.R.; Evans, T.J.; Przybilski, R.; Fineran, P.C.; Salmond, G.P.C. Viral evasion of a bacterial suicide system by RNA-based molecular mimicry enables infectious altruism. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Unterholzner, S.J.; Poppenberger, B.; Rozhon, W. Toxin-antitoxin Systems: Biology, identification, and application. Mob. Genet. Elem. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Szekeres, S.; Dauti, M.; Wilde, C.; Mazel, D.; Rowe-Magnus, D.A. Chromosomal toxin-antitoxin loci can diminish large-scale genome reductions in the absence of selection. Mol. Microbiol. 2007, 63, 1588–1605. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wood, T.K. Toxin-antitoxin systems influence biofilm and persister cell formation and the general stress response. Appl. Environ. Microbiol. 2011, 77, 5577–5583. [Google Scholar] [CrossRef] [PubMed]
- Mutschler, H.; Meinhart, A. ε/ζ Systems: Their role in resistance, virulence, and their potential for antibiotic development. J. Mol. Med. 2011, 89, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Bertram, R.; Schuster, C.F. Post-transcriptional regulation of gene expression in bacterial pathogens by toxin-antitoxin systems. Front. Cell. Infect. Microbiol. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Walker, A.N.; Daines, D.A. Toxin-antitoxin loci vapBC-1 and vapXD contribute to survival and virulence in nontypeable Haemophilus influenzae. BMC Microbiol. 2012, 12. [Google Scholar] [CrossRef] [PubMed]
- Fozo, E.M.; Hemm, M.R.; Storz, G. Small toxic proteins and the antisense RNAs that repress them. Microbiol. Mol. Biol. Rev. 2008, 72, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Blower, T.R.; Short, F.L.; Rao, F.; Mizuguchi, K.; Pei, X.Y.; Fineran, P.C.; Luisi, B.F.; Salmond, G.P.C. Identification and classification of bacterial Type III toxin-antitoxin systems encoded in chromosomal and plasmid genomes. Nucleic Acids Res. 2012, 40, 6158–6173. [Google Scholar] [CrossRef] [PubMed]
- Masuda, H.; Tan, Q.; Awano, N.; Wu, K.-P.; Inouye, M. YeeU enhances the bundling of cytoskeletal polymers of MreB and FtsZ, antagonizing the CbtA (YeeV) toxicity in Escherichia coli. Mol. Microbiol. 2012, 84, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lord, D.M.; Cheng, H.-Y.; Osbourne, D.O.; Hong, S.H.; Sanchez-Torres, V.; Quiroga, C.; Zheng, K.; Herrmann, T.; Peti, W.; et al. A new type V toxin-antitoxin system where mRNA for toxin GhoT is cleaved by antitoxin GhoS. Nat. Chem. Biol. 2012, 8, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Markovski, M.; Wickner, S. Preventing bacterial suicide: A novel toxin-antitoxin strategy. Mol. Cell 2013, 52, 611–612. [Google Scholar] [CrossRef] [PubMed]
- Aakre, C.D.; Phung, T.N.; Huang, D.; Laub, M.T. A bacterial toxin inhibits DNA replication elongation through a direct interaction with the β sliding clamp. Mol. Cell 2013, 52, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.T.; Balsa, D.; Espinosa, M. One cannot rule them all: Are bacterial toxins-antitoxins druggable? FEMS Microbiol. Rev. 2015, 39, 522–540. [Google Scholar] [CrossRef] [PubMed]
- Stieber, D.; Gabant, P.; Szpirer, C.Y. The art of selective killing: Plasmid toxin/antitoxin systems and their technological applications. Biotechniques 2008, 45, 344–346. [Google Scholar] [CrossRef] [PubMed]
- Kristoffersen, P.; Jensen, G.B.; Gerdes, K.; Piskur, J. Bacterial toxin-antitoxin gene system as containment control in yeast cells. Appl. Environ. Microbiol. 2000, 66, 5524–5526. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.M.; Gerdes, K.; Tunnacliffe, A. Bacterial toxin RelE induces apoptosis in human cells. FEBS Lett. 2002, 519, 191–194. [Google Scholar] [CrossRef]
- De la Cueva-Méndez, G.; Mills, A.D.; Clay-Farrace, L.; Díaz-Orejas, R.; Laskey, R.A. Regulatable killing of eukaryotic cells by the prokaryotic proteins Kid and Kis. EMBO J. 2003, 22, 246–251. [Google Scholar] [CrossRef] [PubMed]
- García, J.L.; Díaz, E. Plasmids as tools for containment. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef]
- Torres, B.; Jaenecke, S.; Timmis, K.N.; García, J.L.; Díaz, E. A dual lethal system to enhance containment of recombinant micro-organisms. Microbiology 2003, 149, 3595–3601. [Google Scholar] [CrossRef] [PubMed]
- Mandell, D.J.; Lajoie, M.J.; Mee, M.T.; Takeuchi, R.; Kuznetsov, G.; Norville, J.E.; Gregg, C.J.; Stoddard, B.L.; Church, G.M. Biocontainment of genetically modified organisms by synthetic protein design. Nature 2015, 518, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Rovner, A.J.; Haimovich, A.D.; Katz, S.R.; Li, Z.; Grome, M.W.; Gassaway, B.M.; Amiram, M.; Patel, J.R.; Gallagher, R.R.; Rinehart, J.; et al. Recoded organisms engineered to depend on synthetic amino acids. Nature 2015, 518, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Kroll, J.; Klinter, S.; Schneider, C.; Voss, I.; Steinbüchel, A. Plasmid addiction systems: Perspectives and applications in biotechnology. Microb. Biotechnol. 2010, 3, 634–657. [Google Scholar] [CrossRef] [PubMed]
- Zielenkiewicz, U.; Kowalewska, M.; Kaczor, C.; Ceglowski, P. In vivo interactions between toxin-antitoxin proteins epsilon and zeta of streptococcal plasmid pSM19035 in Saccharomyces cerevisiae. J. Bacteriol. 2009, 191, 3677–3684. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Gómez, A.J.; Lemonnier, M.; Santos-Sierra, S.; Berzal-Herranz, A.; Díaz-Orejas, R. RNase/anti-RNase activities of the bacterial parD toxin-antitoxin system. J. Bacteriol. 2005, 187, 3151–3157. [Google Scholar] [CrossRef] [PubMed]
- Mutschler, H.; Gebhardt, M.; Shoeman, R.L.; Meinhart, A. A novel mechanism of programmed cell death in bacteria by toxin-antitoxin systems corrupts peptidoglycan synthesis. PLoS Biol. 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- Balan, A.; Schenberg, A.C.G. A conditional suicide system for Saccharomyces cerevisiae relying on the intracellular production of the Serratia marcescens nuclease. Yeast 2005, 22, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Cabib, E.; Farkas, V.; Kosik, O.; Blanco, N.; Arroyo, J.; McPhie, P. Assembly of the yeast cell wall: Crh1p and Crh2p act as transglycosylases in vivo and in vitro. J. Biol. Chem. 2008, 283, 29859–29872. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.T.Y.; Lee, J.W.; Cameron, D.E.; Bashor, C.J.; Collins, J.J. “Deadman” and “Passcode” microbial kill switches for bacterial containment. Nat. Chem. Biol. 2016, 12, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Jiang, W.; Yang, S. MazF as a counter-selectable marker for unmarked genetic modification of Pichia pastoris. FEMS Yeast Res. 2009, 9, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Sun, H.; Li, P.; He, N.; Zhu, T.; Li, Y. Enhancement of the gene targeting efficiency of non-conventional yeasts by increasing genetic redundancy. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.J. Improving containment strategies in biopharming. Plant Biotechnol. J. 2007, 5, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Kamle, S.; Ali, S. Genetically modified crops: Detection strategies and biosafety issues. Gene 2013, 522, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Millwood, R.J.; Neal Stewart, C., Jr. Gene use restriction technologies for transgenic plant bioconfinement. Plant Biotechnol. J. 2013, 11, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, D.; Zhao, S.; Shen, Z. A double built-in containment strategy for production of recombinant proteins in transgenic rice. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, L. Genetic use restriction technologies: A review. Plant Biotechnol. J. 2014, 12, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Kempe, K.; Rubtsova, M.; Gils, M. Split-gene system for hybrid wheat seed production. Proc. Natl. Acad. Sci. USA 2014, 111, 9097–9102. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.; Roque, E.; Pineda, B.; Cañas, L.; Rodriguez-Concepción, M.; Beltrán, J.P.; Gómez-Mena, C. Early anther ablation triggers parthenocarpic fruit development in tomato. Plant Biotechnol. J. 2013, 11, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Ulyanova, V.; Vershinina, V.; Ilinskaya, O. Barnase and binase: Twins with distinct fates. FEBS J. 2011, 278, 3633–3643. [Google Scholar] [CrossRef] [PubMed]
- Mariani, C.; de Beuckeleer, M.; Truettner, J.; Leemans, J.; Goldberg, R.B. Induction of male sterility in plants by a chimaeric ribonuclease gene. Nature 1990, 347, 737–741. [Google Scholar] [CrossRef]
- Goldman, M.H.; Goldberg, R.B.; Mariani, C. Female sterile tobacco plants are produced by stigma-specific cell ablation. EMBO J. 1994, 13, 2976–2984. [Google Scholar]
- Beals, T.P.; Goldberg, R.B. A novel cell ablation strategy blocks tobacco anther dehiscence. Plant Cell 1997, 9, 1527–1545. [Google Scholar] [CrossRef] [PubMed]
- Gardner, N.; Felsheim, R.; Smith, A.G. Production of male- and female-sterile plants through reproductive tissue ablation. J. Plant Physiol. 2009, 166, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Munemura, I.; Hinata, K.; Yamamura, S. Bisexual sterility conferred by the differential expression of barnase and barstar: A simple and efficient method of transgene containment. Plant Cell Rep. 2006, 25, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Bisht, N.C.; Jagannath, A.; Augustine, R.; Burma, P.K.; Gupta, V.; Pradhan, A.K.; Pental, D. Effective restoration of male-sterile (barnase) lines requires overlapping and higher levels of barstar expression: A multi-generation field analysis in Brassica juncea. J. Plant Biochem. Biotechnol. 2014, 24, 393–399. [Google Scholar] [CrossRef]
- Millwood, R.J.; Moon, H.S.; Poovaiah, C.R.; Muthukumar, B.; Rice, J.H.; Abercrombie, J.M.; Abercrombie, L.L.; Green, W.D.; Stewart, C.N. Engineered selective plant male sterility through pollen-specific expression of the EcoRI restriction endonuclease. Plant Biotechnol. J. 2015. [Google Scholar] [CrossRef] [PubMed]
- Iida, S.; Terada, R. Modification of endogenous natural genes by gene targeting in rice and other higher plants. Plant Mol. Biol. 2005, 59, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Bakar, F.A.; Yeo, C.C.; Harikrishna, J.A. Expression of the Streptococcus pneumoniae yoeB chromosomal toxin gene causes cell death in the model plant Arabidopsis thaliana. BMC Biotechnol. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Brand, L.; Horler, M.; Nuesch, E.; Vassalli, S.; Barrell, P.; Yang, W.; Jefferson, R.A.; Grossniklaus, U.; Curtis, M.D. A versatile and reliable two-component system for tissue-specific gene induction in arabidopsis. Plant Physiol. 2006, 141, 1194–1204. [Google Scholar] [CrossRef] [PubMed]
- Borgen, B.H.; Thangstad, O.P.; Ahuja, I.; Rossiter, J.T.; Bones, A.M. Removing the mustard oil bomb from seeds: Transgenic ablation of myrosin cells in oilseed rape (Brassica napus) produces MINELESS seeds. J. Exp. Bot. 2010, 61, 1683–1697. [Google Scholar] [CrossRef] [PubMed]
- Nieto, C.; Cherny, I.; Khoo, S.K.; de Lacoba, M.G.; Chan, W.T.; Yeo, C.C.; Gazit, E.; Espinosa, M. The yefM-yoeB toxin-antitoxin systems of Escherichia coli and Streptococcus pneumoniae: Functional and structural correlation. J. Bacteriol. 2007, 189, 1266–1278. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Niu, Q.-W.; Chua, N.-H. An estrogen receptor-based transactivator XVE mediates highly inducible gene expression in transgenic plants. Plant J. 2000, 24, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Slanchev, K.; Stebler, J.; de la Cueva-Méndez, G.; Raz, E. Development without germ cells: The role of the germ line in zebrafish sex differentiation. Proc. Natl. Acad. Sci. USA 2005, 102, 4074–4079. [Google Scholar] [CrossRef] [PubMed]
- Shimazu, T.; Degenhardt, K.; Nur-E-Kamal, A.; Zhang, J.; Yoshida, T.; Zhang, Y.; Mathew, R.; White, E.; Inouye, M. NBK/BIK antagonizes MCL-1 and BCL-XL and activates BAK-mediated apoptosis in response to protein synthesis inhibition. Genes Dev. 2007, 21, 929–941. [Google Scholar] [CrossRef] [PubMed]
- Browne, S.M.; Al-Rubeai, M. Selection methods for high-producing mammalian cell lines. Trends Biotechnol. 2007, 25, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Nehlsen, K.; Herrmann, S.; Zauers, J.; Hauser, H.; Wirth, D. Toxin-antitoxin based transgene expression in mammalian cells. Nucleic Acids Res. 2010, 38. [Google Scholar] [CrossRef] [PubMed]
- Chono, H.; Matsumoto, K.; Tsuda, H.; Saito, N.; Lee, K.; Kim, S.; Shibata, H.; Ageyama, N.; Terao, K.; Yasutomi, Y.; et al. Acquisition of HIV-1 resistance in T lymphocytes using an ACA-specific E. coli mRNA interferase. Hum. Gene Ther. 2011, 22, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Hoeflich, K.P.; Ikura, M.; Qing, G.; Inouye, M. MazF cleaves cellular mRNAs specifically at ACA to block protein synthesis in Escherichia coli. Mol. Cell 2003, 12, 913–923. [Google Scholar] [CrossRef]
- Chono, H.; Saito, N.; Tsuda, H.; Shibata, H.; Ageyama, N.; Terao, K.; Yasutomi, Y.; Mineno, J.; Kato, I. In vivo safety and persistence of endoribonuclease gene-transduced CD4+ T cells in cynomolgus macaques for HIV-1 gene therapy model. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-H.; Yamaguchi, Y.; Inouye, M. Intramolecular regulation of the sequence-specific mRNA interferase activity of MazF fused to a MazE fragment with a linker cleavable by specific proteases. Appl. Environ. Microbiol. 2012, 78, 3794–3799. [Google Scholar] [CrossRef] [PubMed]
- Shapira, A.; Shapira, S.; Gal-Tanamy, M.; Zemel, R.; Tur-Kaspa, R.; Benhar, I. Removal of hepatitis C virus-infected cells by a zymogenized bacterial toxin. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- De la Cueva-Méndez, G. Systems and Methods for Diminishing Cell Growth and Inducing Selective Killing of Target Cells. WO2013037504 A4, 7 June 2013. [Google Scholar]
- Preston, M.A.; Pimentel, B.; Bermejo-Rodríguez, C.; Dionne, I.; Turnbull, A.; de la Cueva-Méndez, G. Repurposing a prokaryotic toxin-antitoxin system for the selective killing of oncogenically stressed human cells. ACS Synth. Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; de Torrontegui, G.; Díaz, R. Identification of components of a new stability system of plasmid R1, ParD, that is close to the origin of replication of this plasmid. Mol. Gen. Genet. 1987, 210, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Inouye, M. Type II toxin-antitoxin loci: The mazEF Family. In Prokaryotic Toxin-Antitoxins; Gerdes, K., Ed.; Springer-Verlag: Berlin, Germnay, 2013; pp. 107–136. [Google Scholar]
- Wieteska, Ł.; Skulimowski, A.; Cybula, M.; Szemraj, J. Toxins vapC and pasB from prokaryotic TA modules remain active in mammalian cancer cells. Toxins 2014, 6, 2948–2961. [Google Scholar] [CrossRef] [PubMed]
- De la Cueva-Méndez, G.; Pimentel, B. Biotechnological and medical exploitation of toxin-antitoxin genes and their components. In Prokaryotic Toxin-Antitoxins; Gerdes, K., Ed.; Springer: Berlin, Germany, 2013; pp. 341–360. [Google Scholar]
- Llosa, M.; Zupan, J.; Baron, C.; Zambryski, P. The N- and C-terminal portions of the agrobacterium VirB1 protein independently enhance tumorigenesis. J. Bacteriol. 2000, 182, 3437–3445. [Google Scholar] [CrossRef] [PubMed]
- Draper, O.; César, C.E.; Machón, C.; de la Cruz, F.; Llosa, M. Site-specific recombinase and integrase activities of a conjugative relaxase in recipient cells. Proc. Natl. Acad. Sci. USA 2005, 102, 16385–16390. [Google Scholar] [CrossRef] [PubMed]
- Llosa, M.; Roy, C.; Dehio, C. Bacterial type IV secretion systems in human disease. Mol. Microbiol. 2009, 73, 141–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Prieto, C.; Agúndez, L.; Linden, R.M.; Llosa, M. HUH site-specific recombinases for targeted modification of the human genome. Trends Biotechnol. 2013, 31, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, K. Prokaryotic Toxin-Antitoxins, 1st ed.; Springer-Verlag: Berlin, Germnay, 2013. [Google Scholar]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeo, C.C.; Abu Bakar, F.; Chan, W.T.; Espinosa, M.; Harikrishna, J.A. Heterologous Expression of Toxins from Bacterial Toxin-Antitoxin Systems in Eukaryotic Cells: Strategies and Applications. Toxins 2016, 8, 49. https://doi.org/10.3390/toxins8020049
Yeo CC, Abu Bakar F, Chan WT, Espinosa M, Harikrishna JA. Heterologous Expression of Toxins from Bacterial Toxin-Antitoxin Systems in Eukaryotic Cells: Strategies and Applications. Toxins. 2016; 8(2):49. https://doi.org/10.3390/toxins8020049
Chicago/Turabian StyleYeo, Chew Chieng, Fauziah Abu Bakar, Wai Ting Chan, Manuel Espinosa, and Jennifer Ann Harikrishna. 2016. "Heterologous Expression of Toxins from Bacterial Toxin-Antitoxin Systems in Eukaryotic Cells: Strategies and Applications" Toxins 8, no. 2: 49. https://doi.org/10.3390/toxins8020049
APA StyleYeo, C. C., Abu Bakar, F., Chan, W. T., Espinosa, M., & Harikrishna, J. A. (2016). Heterologous Expression of Toxins from Bacterial Toxin-Antitoxin Systems in Eukaryotic Cells: Strategies and Applications. Toxins, 8(2), 49. https://doi.org/10.3390/toxins8020049