
The Tip of the Four N-Terminal α-Helices of Clostridium sordellii Lethal Toxin Contains the Interaction Site with Membrane Phosphatidylserine Facilitating Small GTPases Glucosylation


 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Hydrophobic Patches on TcsL 1–93 and Mutation Selection
2.2. Interaction of TcsL-cat Mutants with Brain PS
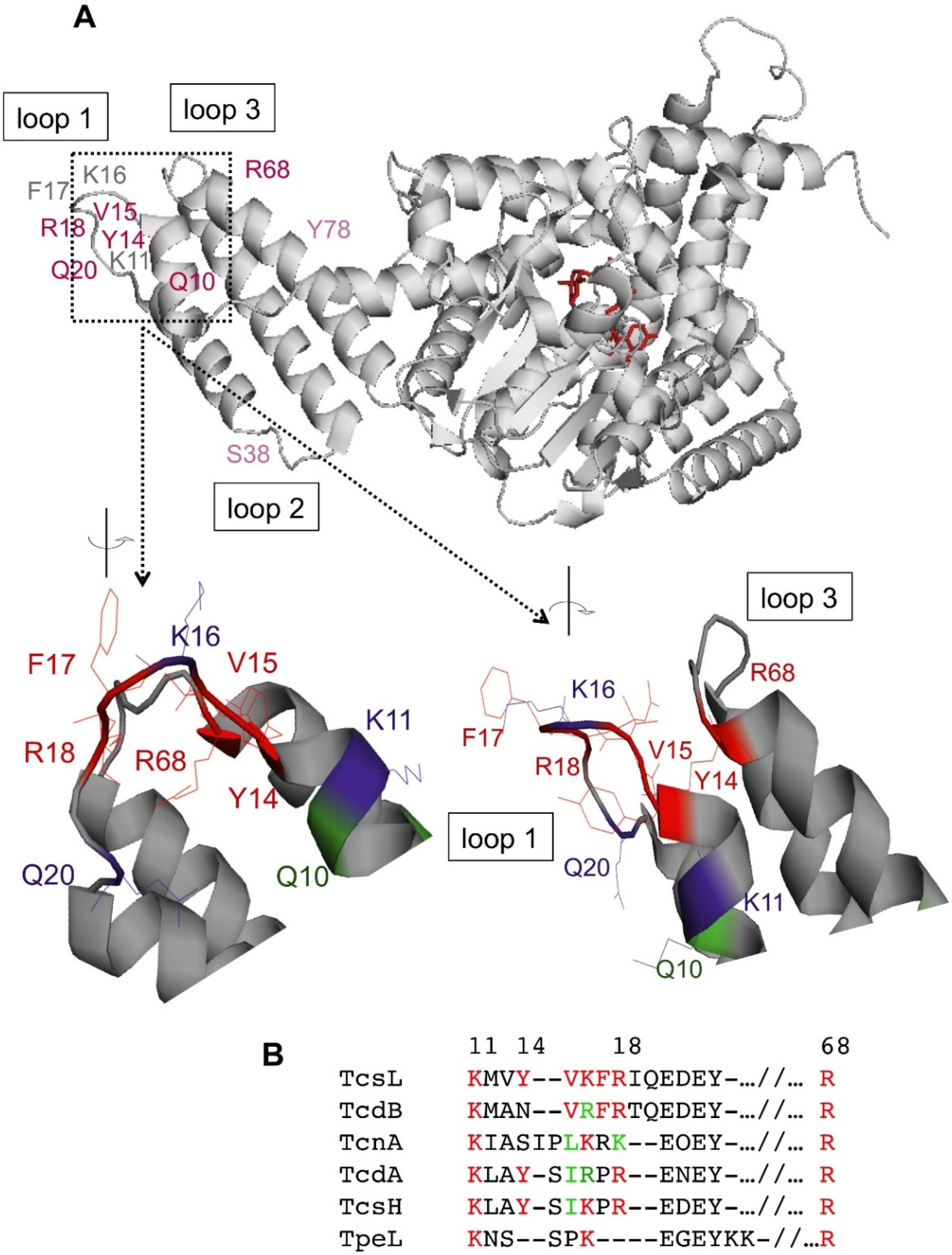
2.3. Localization of the PS Binding Site on TcsL
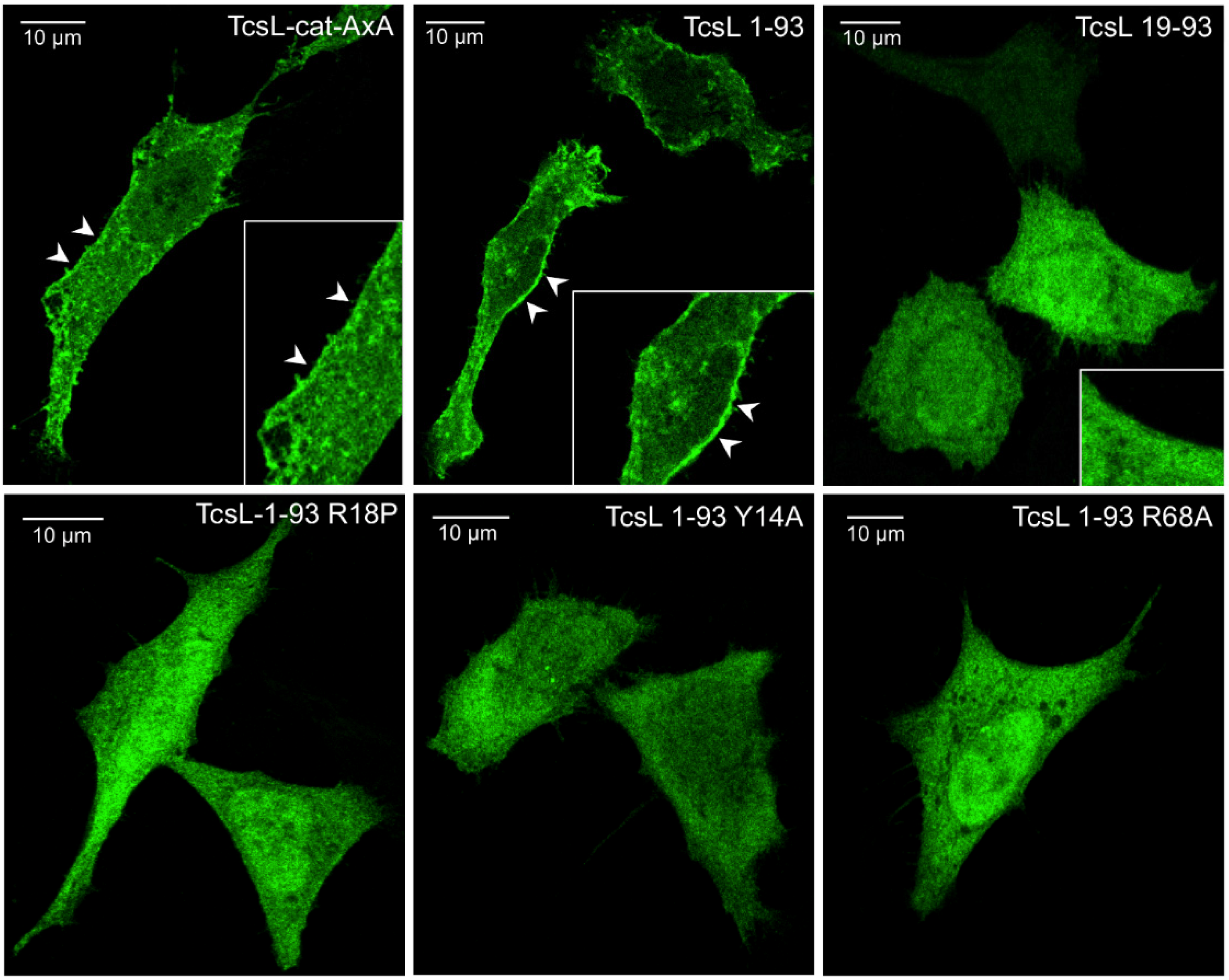
2.4. Binding Site to BPS Is Required for TcsL-cat Localization to Cell Membrane
2.5. Effects of TcsL-cat Mutants on Cell Viability
2.6. Glucosylation Activity of TcsL-cat Mutants
2.7. Model of TcsL-Cat Interaction with PS
3. Discussion
3.1. Identification of the PS Binding Site on the Tip of the N-Terminal Four-Helix Bundle
3.2. The TcsL-cat N-Terminal Domain Is Involved in Membrane Location and Modulates the Enzymatic Activity towards the Membrane-Bound Substrates
3.3. TcsL-cat Binding to Membrane in Appropriate Orientation Is Essential for Efficient Modification of Membrane Bound Ras and Subsequent Cytotoxicity
4. Conclusions
5. Experimental Procedures
5.1. Cloning and Protein Production
5.2. ELISA Test with Immobilized Phospholipids
5.3. Circular Dichroism (Far-UV) Spectroscopy
5.4. Construction of TcsL AXA, TcsL 1-93, TcsL 19-93, and TcsL 1-93 Mutants with GFP-Fusion
5.5. Cell Line, Transfection, and Microscopy
5.6. Delivery of TcsL-cat and Mutants into Cultured Cells by Electroporation
5.7. Proliferation Assay
5.8. In Vitro Glucosylation Assay
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Agrawal, P.; Garg, R. Fulminant leukemoid reaction due to postpartum Clostridium sordellii infection. J. Glob. Infect. Dis. 2012, 4, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Aldape, M.J.; Bryant, A.E.; Stevens, D.L. Clostridium sordellii infection: Epidemiology, clinical findings, and current perspectives on diagnosis and treatment. Clin. Infect. Dis. 2006, 43, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Aronoff, D.M.; Ballard, J.D. Clostridium sordellii toxic shock syndrome. Lancet Infect. Dis. 2009, 9, 725–726. [Google Scholar] [CrossRef]
- Bouvet, P.; Sautereau, J.; Le Coustumier, A.; Mory, F.; Bouchier, C.; Popoff, M.R. Foot infection by Clostridium sordellii: Case report and review of 15 cases in france. J. Clin. Microbiol. 2015, 53, 1423–1427. [Google Scholar] [CrossRef] [PubMed]
- Aktories, K. Bacterial protein toxins that modify host regulatory GTPases. Nat. Rev. Microbiol. 2011, 9, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Amimoto, K.; Noro, T.; Oishi, E.; Shimizu, M. A novel toxin homologous to large clostridial cytotoxins found in culture supernatant of Clostridium perfringens type C. Microbiology 2007, 153, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Just, I.; Gerhard, R. Large clostridial cytotoxins. Rev. Physiol. Biochem. Pharmacol. 2004, 152, 23–47. [Google Scholar] [PubMed]
- Genth, H.; Pauillac, S.; Schelle, I.; Bouvet, P.; Bouchier, C.; Varela-Chavez, C.; Just, I.; Popoff, M.R. Haemorrhagic toxin and lethal toxin from Clostridium sordellii strain vpi9048: Molecular characterization and comparative analysis of substrate specificity of the large clostridial glucosylating toxins. Cell Microbiol. 2014, 16, 1706–1721. [Google Scholar] [CrossRef] [PubMed]
- Thiele, T.L.; Stuber, T.P.; Hauer, P.J. Detection of Clostridium sordellii strains expressing hemorrhagic toxin (TcsH) and implications for diagnostics and regulation of veterinary vaccines. Vaccine 2013, 31, 5082–5087. [Google Scholar] [CrossRef] [PubMed]
- Popoff, M.R.; Chaves-Olarte, E.; Lemichez, E.; Von Eichel-Streiber, C.; Thelestam, M.; Chardin, P.; Cussac, D.; Antonny, B.; Chavrier, P.; Flatau, G.; et al. Ras, Rap, and Rac small GTP-binding proteins are targets for Clostridium sordellii lethal toxin glucosylation. J. Biol. Chem. 1996, 271, 10217–10224. [Google Scholar] [PubMed]
- Guttenberg, G.; Hornei, S.; Jank, T.; Schwan, C.; Lu, W.; Einsle, O.; Papatheodorou, P.; Aktories, K. Molecular characteristics of Clostridium perfringens TpeL toxin and consequences of mono-O-GlcNAcylation of ras in living cells. J. Biol. Chem. 2012, 287, 24929–24940. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, F.; Rex, G.; Aktories, K.; Just, I. The ras-related protein ral is monoglucosylated by Clostridium sordellii lethal toxin. Biochem. Biophys. Res. Commun. 1996, 227, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Just, I.; Selzer, J.; Wilm, M.; von Eichel-Streiber, C.; Mann, M.; Aktories, K. Glucosylation of Rho proteins by Clostridium difficile toxin B. Nature 1995, 375, 500–503. [Google Scholar] [CrossRef] [PubMed]
- Just, I.; Wilm, M.; Selzer, J.; Rex, G.; von Eichel-Streiber, C.; Mann, M.; Aktories, K. The enterotoxin from Clostridium difficile (ToxA) monoglucosylates the Rho proteins. J. Biol. Chem. 1995, 270, 13932–13936. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, M.; Ohkubo, A.; Oda, M.; Kobayashi, K.; Amimoto, K.; Miyamoto, K.; Sakurai, J. Clostridium perfringens TpeL glycosylates the Rac and Ras subfamily proteins. Infect. Immun. 2011, 79, 905–910. [Google Scholar] [CrossRef] [PubMed]
- Selzer, J.; Hofmann, F.; Rex, G.; Wilm, M.; Mann, M.; Just, I.; Aktories, K. Clostridium novyi α-toxin-catalyzed incorporation of GLCNAC into Rho subfamily proteins. J. Biol. Chem. 1996, 271, 25173–25177. [Google Scholar] [CrossRef] [PubMed]
- Zeiser, J.; Gerhard, R.; Just, I.; Pich, A. Substrate specificity of clostridial glucosylating toxins and their function on colonocytes analyzed by proteomics techniques. J. Proteome Res. 2013, 12, 1604–1618. [Google Scholar] [CrossRef] [PubMed]
- Genth, H.; Dreger, S.C.; Huelsenbeck, J.; Just, I. Clostridium difficile toxins: More than mere inhibitors of Rho proteins. Int. J. Biochem. Cell Biol. 2008, 40, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Genth, H.; Just, I. Large clostridial cytotoxins modifying small GTPases: Cellular aspects. In The Comprehensive Sourcebook of Bacterial Protein Toxins, 4th ed.; Alouf, J.E., Ladant, D., Popoff, M.R., Eds.; Elsevier: Amsterdam, The Netherland, 2015; pp. 426–440. [Google Scholar]
- Jank, T.; Aktories, K. Structure and mode of action of clostridial glucosylating toxins: The ABCD model. Trends Microbiol. 2008, 16, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Genth, H.; Just, I. Functional implications of lethal toxin-catalysed glucosylation of (H/K/N)Ras and Rac1 in Clostridium sordellii-associated disease. Eur. J. Cell Biol. 2011, 90, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Geny, B.; Grassart, A.; Manich, M.; Chicanne, G.; Payrastre, B.; Sauvonnet, N.; Popoff, M.R. Rac1 inactivation by lethal toxin from Clostridium sordellii modifies focal adhesions upstream of actin depolymerization. Cell Microbiol. 2010, 12, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Popoff, M.R.; Geny, B. Rho/Ras-GTPase-dependent and -independent activity of clostridial glucosylating toxins. J. Med. Microbiol. 2011, 60, 1057–1069. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, S.C.; Klose, I.; Reichenbach, M.; Huelsenbeck, J.; Genth, H. Distinct kinetics of (H/K/N)Ras glucosylation and Rac1 glucosylation catalysed by Clostridium sordellii lethal toxin. FEBS Lett. 2009, 583, 3133–3139. [Google Scholar] [CrossRef] [PubMed]
- LaFrance, M.E.; Farrow, M.A.; Chandrasekaran, R.; Sheng, J.; Rubin, D.H.; Lacy, D.B. Identification of an epithelial cell receptor responsible for Clostridium difficile TcdB-induced cytotoxicity. Proc. Natl. Acad. Sci. USA 2015, 112, 7073–7078. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Zhang, H.; Cai, C.; Zhu, S.; Zhou, Y.; Yang, X.; He, R.; Li, C.; Guo, S.; Li, S.; et al. Chondroitin sulfate proteoglycan 4 functions as the cellular receptor for Clostridium difficile toxin B. Cell Res. 2015, 25, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Genisyuerek, S.; Papatheodorou, P.; Guttenberg, G.; Schubert, R.; Benz, R.; Aktories, K. Structural determinants for membrane insertion, pore formation and translocation of Clostridium difficile toxin B. Mol. Microbiol. 2011, 79, 1643–1654. [Google Scholar] [CrossRef] [PubMed]
- Guttenberg, G.; Papatheodorou, P.; Genisyuerek, S.; Lu, W.; Jank, T.; Einsle, O.; Aktories, K. Inositol hexakisphosphate-dependent processing of Clostridium sordellii lethal toxin and Clostridium novyi alpha-toxin. J. Biol. Chem. 2011, 286, 14779–14786. [Google Scholar] [CrossRef] [PubMed]
- Papatheodorou, P.; Zamboglou, C.; Genisyuerek, S.; Guttenberg, G.; Aktories, K. Clostridial glucosylating toxins enter cells via clathrin-mediated endocytosis. PLoS One 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Prior, I.A.; Hancock, J.F. Ras trafficking, localization and compartmentalized signalling. Semin. Cell Dev. Biol. 2012, 23, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Cereijido, M.; Contreras, R.G.; Shoshani, L.; Garcia-Villegas, M.R. Membrane targeting. Prog. Biophys. Mol. Biol. 2003, 81, 81–115. [Google Scholar] [CrossRef]
- Kamitani, S.; Kitadokoro, K.; Miyazawa, M.; Toshima, H.; Fukui, A.; Abe, H.; Miyake, M.; Horiguchi, Y. Characterization of the membrane-targeting C1 domain in Pasteurella multocida toxin. J. Biol. Chem. 2010, 285, 25467–25475. [Google Scholar] [CrossRef] [PubMed]
- Geissler, B.; Ahrens, S.; Satchell, K.J. Plasma membrane association of three classes of bacterial toxins is mediated by a basic-hydrophobic motif. Cell Microbiol. 2012, 14, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Geissler, B.; Tungekar, R.; Satchell, K.J. Identification of a conserved membrane localization domain within numerous large bacterial protein toxins. Proc. Natl. Acad. Sci. USA 2010, 107, 5581–5586. [Google Scholar] [CrossRef] [PubMed]
- Satchell, K.J. Structure and function of MARTX toxins and other large repetitive RTX proteins. Annu. Rev. Microbiol. 2011, 65, 71–90. [Google Scholar] [CrossRef] [PubMed]
- Brothers, M.C.; Geissler, B.; Hisao, G.S.; Wilson, B.A.; Satchell, K.J.; Rienstra, C.M. Backbone and side-chain assignments of an effector membrane localization domain from Vibrio vulnificus MARTX toxin. Biomol. NMR Assign 2014, 8, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Chavez, C.V.; Hoos, S.; Haustant, G.M.; Chenal, A.; England, P.; Blondel, A.; Pauillac, S.; Lacy, D.B.; Popoff, M.R. The catalytic domains of Clostridium sordellii lethal toxin and related large clostridial glucosylating toxins specifically recognize the negatively charged phospholipids phosphatidylserine and phosphatidic acid. Cell Microbiol. 2015, 17, 1477–1493. [Google Scholar] [CrossRef] [PubMed]
- Mesmin, B.; Robbe, K.; Geny, B.; Luton, F.; Brandolin, G.; Popoff, M.R.; Antonny, B. A phosphatidylserine-binding site in the cytosolic fragment of Clostridium sordellii lethal toxin facilitates glucosylation of membrane-bound Rac and is required for cytotoxicity. J. Biol. Chem. 2004, 279, 49876–49882. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, M.O.; Jank, T.; Aktories, K.; Schulz, G.E. Conformational changes and reaction of clostridial glycosylating toxins. J. Mol. Biol. 2008, 377, 1346–1356. [Google Scholar] [CrossRef] [PubMed]
- Reinert, D.J.; Jank, T.; Aktories, K.; Schulz, G.E. Structural basis for the function of Clostridium difficile toxin B. J. Mol. Biol. 2005, 351, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Rarey, M.; Kramer, B.; Lengauer, T.; Klebe, G. A fast flexible docking method using an incremental construction algorithm. J. Mol. Biol. 1996, 261, 470–489. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, F.; Busch, C.; Aktories, K. Chimeric clostridial cytotoxins: Identification of the N-terminal region involved in protein substrate recognition. Infect. Immun. 1998, 66, 1076–1081. [Google Scholar] [PubMed]
- Jank, T.; Giesemann, T.; Aktories, K. Clostridium difficile glucosyltransferase toxin B-essential amino acids for substrate binding. J. Biol. Chem. 2007, 282, 35222–35231. [Google Scholar] [CrossRef] [PubMed]
- Karst, J.C.; Sotomayor Perez, A.C.; Guijarro, J.I.; Raynal, B.; Chenal, A.; Ladant, D. Calmodulin-induced conformational and hydrodynamic changes in the catalytic domain of Bordetella pertussis adenylate cyclase toxin. Biochemistry 2010, 49, 318–328. [Google Scholar] [CrossRef] [PubMed]









© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varela Chavez, C.; Haustant, G.M.; Baron, B.; England, P.; Chenal, A.; Pauillac, S.; Blondel, A.; Popoff, M.-R. The Tip of the Four N-Terminal α-Helices of Clostridium sordellii Lethal Toxin Contains the Interaction Site with Membrane Phosphatidylserine Facilitating Small GTPases Glucosylation. Toxins 2016, 8, 90. https://doi.org/10.3390/toxins8040090
Varela Chavez C, Haustant GM, Baron B, England P, Chenal A, Pauillac S, Blondel A, Popoff M-R. The Tip of the Four N-Terminal α-Helices of Clostridium sordellii Lethal Toxin Contains the Interaction Site with Membrane Phosphatidylserine Facilitating Small GTPases Glucosylation. Toxins. 2016; 8(4):90. https://doi.org/10.3390/toxins8040090
Chicago/Turabian StyleVarela Chavez, Carolina, Georges Michel Haustant, Bruno Baron, Patrick England, Alexandre Chenal, Serge Pauillac, Arnaud Blondel, and Michel-Robert Popoff. 2016. "The Tip of the Four N-Terminal α-Helices of Clostridium sordellii Lethal Toxin Contains the Interaction Site with Membrane Phosphatidylserine Facilitating Small GTPases Glucosylation" Toxins 8, no. 4: 90. https://doi.org/10.3390/toxins8040090
APA StyleVarela Chavez, C., Haustant, G. M., Baron, B., England, P., Chenal, A., Pauillac, S., Blondel, A., & Popoff, M. -R. (2016). The Tip of the Four N-Terminal α-Helices of Clostridium sordellii Lethal Toxin Contains the Interaction Site with Membrane Phosphatidylserine Facilitating Small GTPases Glucosylation. Toxins, 8(4), 90. https://doi.org/10.3390/toxins8040090