
Relationship between vacA Types and Development of Gastroduodenal Diseases



{kind=link}
Abstract
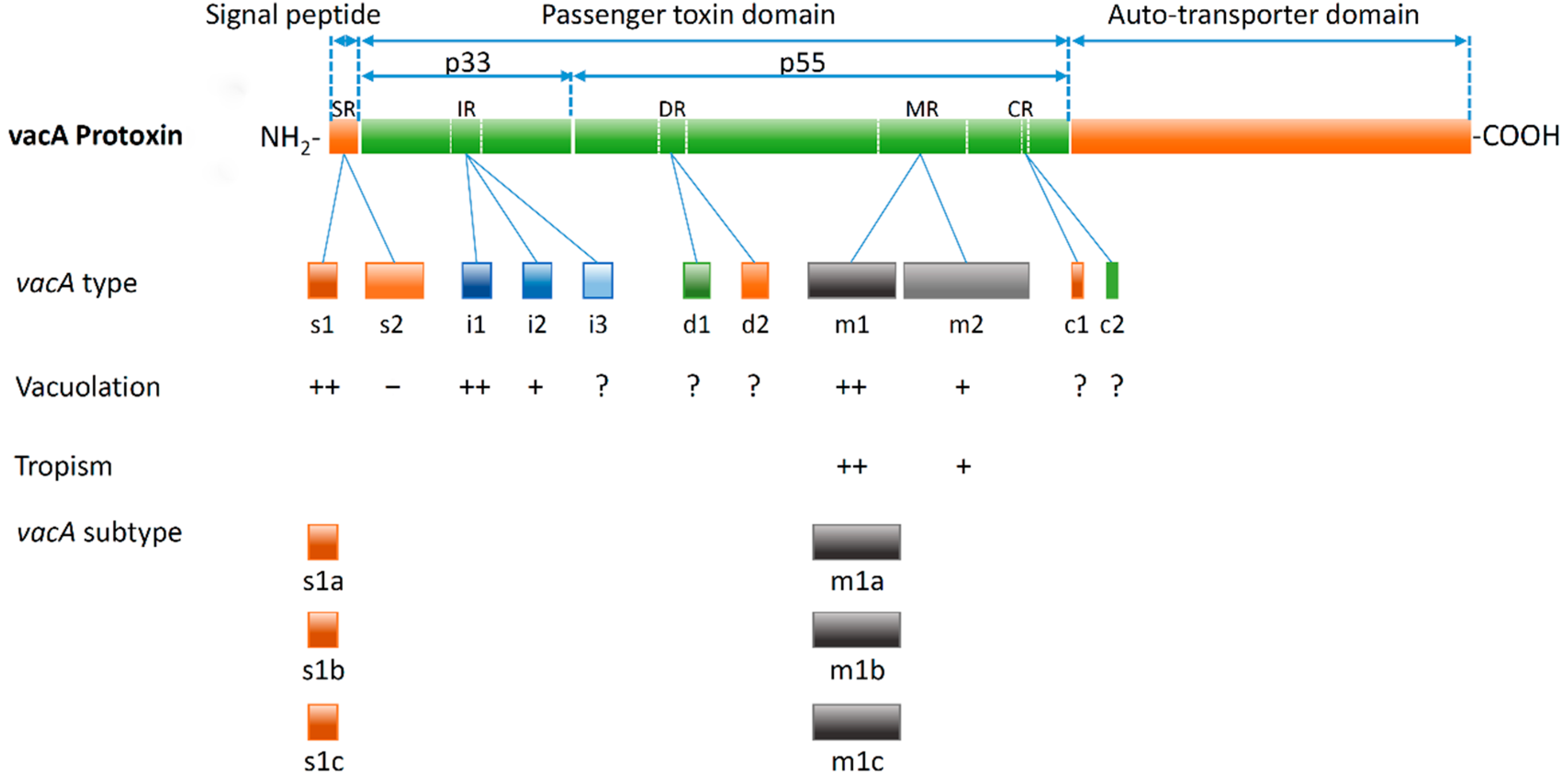
:1. VacA and Allelic Diversity of vacA
2. The Association of vacA Genotypes and Cytotoxin
3. The Relationship between vacA Polymorphism and H. pylori Persistence
4. Role of the vacA Genotype in Gastroduodenal Diseases
5. Role of vacA Genotype in Extragastroduodenal Diseases
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Ferreira, R.M.; Machado, J.C.; Letley, D.; Atherton, J.C.; Pardo, M.L.; Gonzalez, C.A.; Carneiro, F.; Figueiredo, C. A novel method for genotyping the Helicobacter pylori vacA intermediate region directly in gastric biopsy specimens. J. Clin. Microbiol. 2012, 50, 3983–3989. [Google Scholar] [CrossRef] [PubMed]
- Memon, A.A.; Hussein, N.R.; Miendje Deyi, V.Y.; Burette, A.; Atherton, J.C. Vacuolating cytotoxin genotypes are strong markers of gastric cancer and duodenal ulcer-associated Helicobacter pylori strains: A matched case-control study. J. Clin. Microbiol. 2014, 52, 2984–2989. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, T.; Yoshimura, T.; Mikami, T.; Fukuda, S.; Crabtree, J.E.; Munakata, A. Evaluation of Helicobacter pylori vacA genotype in Japanese patients with gastric cancer. J. Clin. Pathol. 1998, 51, 299–301. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Tummuru, M.K.; Cao, P.; Thompson, S.A.; Blaser, M.J. Divergence of genetic sequences for the vacuolating cytotoxin among Helicobacter pylori strains. J. Biol. Chem. 1994, 269, 10566–10573. [Google Scholar] [PubMed]
- Atherton, J.C.; Cao, P.; Peek, R.M., Jr.; Tummuru, M.K.; Blaser, M.J.; Cover, T.L. Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori. Association of specific vacA types with cytotoxin production and peptic ulceration. J. Biol. Chem. 1995, 270, 17771–17777. [Google Scholar] [PubMed]
- Forsyth, M.H.; Atherton, J.C.; Blaser, M.J.; Cover, T.L. Heterogeneity in levels of vacuolating cytotoxin gene (vacA) transcription among Helicobacter pylori strains. Infect. Immun. 1998, 66, 3088–3094. [Google Scholar] [PubMed]
- Letley, D.P.; Atherton, J.C. Natural diversity in the n terminus of the mature vacuolating cytotoxin of Helicobacter pylori determines cytotoxin activity. J. Bacteriol. 2000, 182, 3278–3280. [Google Scholar] [CrossRef] [PubMed]
- Strobel, S.; Bereswill, S.; Balig, P.; Allgaier, P.; Sonntag, H.G.; Kist, M. Identification and analysis of a new vacA genotype variant of Helicobacter pylori in different patient groups in germany. J. Clin. Microbiol. 1998, 36, 1285–1289. [Google Scholar] [PubMed]
- Van Doorn, L.J.; Figueiredo, C.; Sanna, R.; Pena, S.; Midolo, P.; Ng, E.K.; Atherton, J.C.; Blaser, M.J.; Quint, W.G. Expanding allelic diversity of Helicobacter pylori vacA. J. Clin. Microbiol. 1998, 36, 2597–2603. [Google Scholar] [PubMed]
- Rhead, J.L.; Letley, D.P.; Mohammadi, M.; Hussein, N.; Mohagheghi, M.A.; Eshagh Hosseini, M.; Atherton, J.C. A new Helicobacter pylori vacuolating cytotoxin determinant, the intermediate region, is associated with gastric cancer. Gastroenterology 2007, 133, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.; Olivares, A.; Torres, E.; Yilmaz, O.; Cohen, H.; Perez-Perez, G. Diversity of vacA intermediate region among Helicobacter pylori strains from several regions of the world. J. Clin. Microbiol. 2010, 48, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Torres, V.J.; Ivie, S.E.; McClain, M.S.; Cover, T.L. Functional properties of the p33 and p55 domains of the Helicobacter pylori vacuolating cytotoxin. J. Biol. Chem. 2005, 280, 21107–21114. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Yamaoka, Y. The association of vacA genotype and Helicobacter pylori-related disease in Latin American and African populations. Clin. Microbiol. Infect. 2009, 15, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Zali, M.R.; Yamaoka, Y. The association of vacA genotypes and Helicobacter pylori-related gastroduodenal diseases in the middle east. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.J.; Blanke, S.R. Remodeling the host environment: Modulation of the gastric epithelium by the Helicobacter pylori vacuolating toxin (vacA). Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Basso, D.; Zambon, C.F.; Letley, D.P.; Stranges, A.; Marchet, A.; Rhead, J.L.; Schiavon, S.; Guariso, G.; Ceroti, M.; Nitti, D.; et al. Clinical relevance of Helicobacter pylori cagA and vacA gene polymorphisms. Gastroenterology 2008, 135, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Atherton, J.C.; Sharp, P.M.; Cover, T.L.; Gonzalez-Valencia, G.; Peek, R.M., Jr.; Thompson, S.A.; Hawkey, C.J.; Blaser, M.J. Vacuolating cytotoxin (vacA) alleles of Helicobacter pylori comprise two geographically widespread types, m1 and m2, and have evolved through limited recombination. Curr. Microbiol. 1999, 39, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Pagliaccia, C.; de Bernard, M.; Lupetti, P.; Ji, X.; Burroni, D.; Cover, T.L.; Papini, E.; Rappuoli, R.; Telford, J.L.; Reyrat, J.M. The m2 form of the Helicobacter pylori cytotoxin has cell type-specific vacuolating activity. Proc. Natl. Acad. Sci. USA 1998, 95, 10212–10217. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Fernandez, T.; Burroni, D.; Pagliaccia, C.; Atherton, J.C.; Reyrat, J.M.; Rappuoli, R.; Telford, J.L. Cell specificity of Helicobacter pylori cytotoxin is determined by a short region in the polymorphic midregion. Infect. Immun. 2000, 68, 3754–3757. [Google Scholar] [CrossRef] [PubMed]
- Ogiwara, H.; Sugimoto, M.; Ohno, T.; Vilaichone, R.K.; Mahachai, V.; Graham, D.Y.; Yamaoka, Y. Role of deletion located between the intermediate and middle regions of the Helicobacter pylori vacA gene in cases of gastroduodenal diseases. J. Clin. Microbiol. 2009, 47, 3493–3500. [Google Scholar] [CrossRef] [PubMed]
- Bakhti, S.Z.; Latifi-Navid, S.; Mohammadi, S.; Zahri, S.; Bakhti, F.S.; Feizi, F.; Yazdanbod, A.; Siavoshi, F. Relevance of Helicobacter pylori vacA 3′-end region polymorphism to gastric cancer. Helicobacter 2015. [Google Scholar] [CrossRef] [PubMed]
- Papini, E.; de Bernard, M.; Milia, E.; Bugnoli, M.; Zerial, M.; Rappuoli, R.; Montecucco, C. Cellular vacuoles induced by Helicobacter pylori originate from late endosomal compartments. Proc. Natl. Acad. Sci. USA 1994, 91, 9720–9724. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Blanke, S.R. Helicobacter pylori vacA, a paradigm for toxin multifunctionality. Nat. Rev. Microbiol. 2005, 3, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Molinari, M.; Salio, M.; Galli, C.; Norais, N.; Rappuoli, R.; Lanzavecchia, A.; Montecucco, C. Selective inhibition of Ii-dependent antigen presentation by Helicobacter pylori toxin vacA. J. Exp. Med. 1998, 187, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.Y.; Jones, N.L. Helicobacter pylori strains expressing the vacuolating cytotoxin interrupt phagosome maturation in macrophages by recruiting and retaining taco (coronin 1) protein. Cell Microbiol. 2003, 5, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wandinger-Ness, A.; Goldenring, J.R.; Cover, T.L. Clustering and redistribution of late endocytic compartments in response to Helicobacter pylori vacuolating toxin. Mol. Biol. Cell 2004, 15, 1946–1959. [Google Scholar] [CrossRef] [PubMed]
- McClain, M.S.; Cao, P.; Iwamoto, H.; Vinion-Dubiel, A.D.; Szabo, G.; Shao, Z.; Cover, T.L. A 12-amino-acid segment, present in type s2 but not type s1 Helicobacter pylori VacA proteins, abolishes cytotoxin activity and alters membrane channel formation. J. Bacteriol. 2001, 183, 6499–6508. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.C.; Wang, H.J.; Kuo, C.H. Two distinctive cell binding patterns by vacuolating toxin fused with glutathione s-transferase: One high-affinity m1-specific binding and the other lower-affinity binding for variant m forms. Biochemistry 2001, 40, 11887–11896. [Google Scholar] [CrossRef] [PubMed]
- Letley, D.P.; Rhead, J.L.; Twells, R.J.; Dove, B.; Atherton, J.C. Determinants of non-toxicity in the gastric pathogen Helicobacter pylori. J. Biol. Chem. 2003, 278, 26734–26741. [Google Scholar] [CrossRef] [PubMed]
- Atherton, J.C.; Peek, R.M., Jr.; Tham, K.T.; Cover, T.L.; Blaser, M.J. Clinical and pathological importance of heterogeneity in vacA, the vacuolating cytotoxin gene of Helicobacter pylori. Gastroenterology 1997, 112, 92–99. [Google Scholar] [CrossRef]
- Ferreira, R.M.; Machado, J.C.; Figueiredo, C. Clinical relevance of Helicobacter pylori vacA and cagA genotypes in gastric carcinoma. Best Pract. Res. Clin. Gastroenterol. 2014, 28, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Salama, N.R.; Otto, G.; Tompkins, L.; Falkow, S. Vacuolating cytotoxin of Helicobacter pylori plays a role during colonization in a mouse model of infection. Infect. Immun. 2001, 69, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Ayraud, S.; Janvier, B.; Fauchere, J.L. Experimental colonization of mice by fresh clinical isolates of Helicobacter pylori is not influenced by the cagA status and the vacA genotype. FEMS Immunol. Med. Microbiol. 2002, 34, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.A.; Letley, D.P.; Cook, K.W.; Rhead, J.L.; Zaitoun, A.A.; Ingram, R.J.; Amilon, K.R.; Croxall, N.J.; Kaye, P.V.; Robinson, K.; et al. A role for the vacuolating cytotoxin, vacA, in colonization and Helicobacter pylori-induced metaplasia in the stomach. J. Infect. Dis. 2014, 210, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, F.; Campbell, B.J.; Alfizah, H.; Varro, A.; Zahra, R.; Yamaoka, Y.; Pritchard, D.M. Analysis of clinical isolates of Helicobacter pylori in pakistan reveals high degrees of pathogenicity and high frequencies of antibiotic resistance. Helicobacter 2014, 19, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Rahimian, G.; Sanei, M.H.; Shirzad, H.; Azadegan-Dehkordi, F.; Taghikhani, A.; Salimzadeh, L.; Hashemzadeh-Chaleshtori, M.; Rafieian-Kopaei, M.; Bagheri, N. Virulence factors of Helicobacter pylori VacA increase markedly gastric mucosal TFG-β1-beta1 mRNA expression in gastritis patients. Microb. Pathog. 2014, 67–68, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Neu, B.; Rad, R.; Reindl, W.; Neuhofer, M.; Gerhard, M.; Schepp, W.; Prinz, C. Expression of tumor necrosis factor-α-related apoptosis-inducing ligand and its proapoptotic receptors is down-regulated during gastric infection with virulent cagA+/vacAs1+ Helicobacter pylori strains. J. Infect. Dis. 2005, 191, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Toller, I.M.; Neelsen, K.J.; Steger, M.; Hartung, M.L.; Hottiger, M.O.; Stucki, M.; Kalali, B.; Gerhard, M.; Sartori, A.A.; Lopes, M.; et al. Carcinogenic bacterial pathogen Helicobacter pylori triggers DNA double-strand breaks and a DNA damage response in its host cells. Proc. Natl. Acad. Sci. USA 2011, 108, 14944–14949. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, C.; Figueiredo, C.; Carneiro, F.; Gomes, A.T.; Barreira, R.; Figueira, P.; Salgado, C.; Belo, L.; Peixoto, A.; Bravo, J.C.; et al. Helicobacter pylori genotypes may determine gastric histopathology. Am. J. Pathol. 2001, 158, 647–654. [Google Scholar] [CrossRef]
- Correa, P. Human gastric carcinogenesis: A multistep and multifactorial process—First American cancer society award lecture on cancer epidemiology and prevention. Cancer Res. 1992, 52, 6735–6740. [Google Scholar] [PubMed]
- Jones, N.L.; Day, A.S.; Jennings, H.; Shannon, P.T.; Galindo-Mata, E.; Sherman, P.M. Enhanced disease severity in Helicobacter pylori-infected mice deficient in fas signaling. Infect. Immun. 2002, 70, 2591–2597. [Google Scholar] [CrossRef] [PubMed]
- Peek, R.M., Jr.; Wirth, H.P.; Moss, S.F.; Yang, M.; Abdalla, A.M.; Tham, K.T.; Zhang, T.; Tang, L.H.; Modlin, I.M.; Blaser, M.J. Helicobacter pylori alters gastric epithelial cell cycle events and gastrin secretion in mongolian gerbils. Gastroenterology 2000, 118, 48–59. [Google Scholar] [CrossRef]
- Fan, X.; Crowe, S.E.; Behar, S.; Gunasena, H.; Ye, G.; Haeberle, H.; Van Houten, N.; Gourley, W.K.; Ernst, P.B.; Reyes, V.E. The effect of class II major histocompatibility complex expression on adherence of Helicobacter pylori and induction of apoptosis in gastric epithelial cells: A mechanism for T helper cell type 1-mediated damage. J. Exp. Med. 1998, 187, 1659–1669. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Blaser, M.J. Helicobacter pylori in health and disease. Gastroenterology 2009, 136, 1863–1873. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Jones, K.R.; Olsen, C.H.; Joo, Y.M.; Yoo, Y.J.; Chung, I.S.; Cha, J.H.; Merrell, D.S. Epidemiological link between gastric disease and polymorphisms in vacA and cagA. J. Clin. Microbiol. 2010, 48, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, Y.; Kato, M.; Asaka, M. Geographic differences in gastric cancer incidence can be explained by differences between Helicobacter pylori strains. Intern. Med. 2008, 47, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Zambon, C.F.; Navaglia, F.; Basso, D.; Rugge, M.; Plebani, M. Helicobacter pylori babA2, cagA, and s1 vacA genes work synergistically in causing intestinal metaplasia. J. Clin. Pathol. 2003, 56, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Gunn, M.C.; Stephens, J.C.; Stewart, J.A.; Rathbone, B.J.; West, K.P. The significance of cagA and vacA subtypes of Helicobacter pylori in the pathogenesis of inflammation and peptic ulceration. J. Clin. Pathol. 1998, 51, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Soltermann, A.; Koetzer, S.; Eigenmann, F.; Komminoth, P. Correlation of Helicobacter pylori virulence genotypes vacA and cagA with histological parameters of gastritis and patient’s age. Mod. Pathol. 2007, 20, 878–883. [Google Scholar] [CrossRef] [PubMed]
- Siddique, I.; Al-Qabandi, A.; Al-Ali, J.; Alazmi, W.; Memon, A.; Mustafa, A.S.; Junaid, T.A. Association between Helicobacter pylori genotypes and severity of chronic gastritis, peptic ulcer disease and gastric mucosal interleukin-8 levels: Evidence from a study in the Middle East. Gut Pathog. 2014, 6, 41. [Google Scholar] [CrossRef] [PubMed]
- Trang, T.T.; Shiota, S.; Matsuda, M.; Binh, T.T.; Suzuki, R.; Vilaichone, R.K.; Mahachai, V.; Tshering, L.; Dung, H.D.; Uchida, T.; et al. The prevalence of Helicobacter pylori virulence factors in Bhutan, Vietnam, and Myanmar is related to gastric cancer incidence. Biomed. Res. Int. 2015, 2015, 830813. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C.A.; Figueiredo, C.; Lic, C.B.; Ferreira, R.M.; Pardo, M.L.; Ruiz Liso, J.M.; Alonso, P.; Sala, N.; Capella, G.; Sanz-Anquela, J.M. Helicobacter pylori cagA and vacA genotypes as predictors of progression of gastric preneoplastic lesions: A long-term follow-up in a high-risk area in Spain. Am. J. Gastroenterol. 2011, 106, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.M.; Figueiredo, C.; Bonet, C.; Pardo, M.L.; Liso, J.M.; Alonso, P.; Sala, N.; Capella, G.; Sanz-Anquela, J.M.; Gonzalez, C.A. Helicobacter pylori vacA intermediate region genotyping and progression of gastric preneoplastic lesions. Am. J. Gastroenterol. 2012, 107, 145–146. [Google Scholar] [CrossRef] [PubMed]
- Figura, N.; Valassina, M.; Moretti, E.; Vindigni, C.; Collodel, G.; Iacoponi, F.; Giordano, N.; Roviello, F.; Marrelli, D. Histological variety of gastric carcinoma and Helicobacter pylori cagA and vacA polymorphism. Eur. J. Gastroenterol. Hepatol. 2015, 27, 1017–1021. [Google Scholar] [PubMed]
- Biernat, M.M.; Gosciniak, G.; Iwanczak, B. Prevalence of Helicobacter pylori cagA, vacA, iceA, babA2 genotypes in polish children and adolescents with gastroduodenal disease. Postepy Hig. Med. Doswiadczalnej (Online) 2014, 68, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Almeida, N.; Donato, M.M.; Romaozinho, J.M.; Luxo, C.; Cardoso, O.; Cipriano, M.A.; Marinho, C.; Fernandes, A.; Sofia, C. Correlation of Helicobacter pylori genotypes with gastric histopathology in the central region of a South-European country. Dig. Dis. Sci. 2015, 60, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wu, X.; Liu, Z.; Bu, G.; Li, X.; Qu, N.; Peng, J.; Xu, C.; Shen, S.; Yuan, Y. Association between virulence factors and TRAF1/4-1BB/Bcl-xL expression in gastric mucosa infected with Helicobacter pylori. Gastroenterol. Res. Pract. 2015, 2015, 648479. [Google Scholar] [CrossRef] [PubMed]
- Breurec, S.; Michel, R.; Seck, A.; Brisse, S.; Come, D.; Dieye, F.B.; Garin, B.; Huerre, M.; Mbengue, M.; Fall, C.; et al. Clinical relevance of cagA and vacA gene polymorphisms in Helicobacter pylori isolates from senegalese patients. Clin. Microbiol. Infect. 2012, 18, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Ogiwara, H.; Graham, D.Y.; Yamaoka, Y. vacA i-region subtyping. Gastroenterology 2008, 134, 1267. [Google Scholar] [CrossRef] [PubMed]
- Yordanov, D.; Boyanova, L.; Markovska, R.; Gergova, G.; Mitov, I. Significance of Helicobacter pylori vacA intermediate region genotyping-a Bulgarian study. Diagn. Microbiol. Infect. Dis. 2012, 74, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Mottaghi, B.; Safaralizadeh, R.; Bonyadi, M.; Latifi-Navid, S.; Somi, M.H. Helicobacter pylori vacA i region polymorphism but not babA2 status associated to gastric cancer risk in Northwestern Iran. Clin. Exp. Med. 2014, 16, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rivera, C.; Algood, H.M.; Radin, J.N.; McClain, M.S.; Cover, T.L. The intermediate region of Helicobacter pylori vacA is a determinant of toxin potency in a jurkat T cell assay. Infect. Immun. 2012, 80, 2578–2588. [Google Scholar] [CrossRef] [PubMed]
- Basiri, Z.; Safaralizadeh, R.; Bonyadi, M.J.; Somi, M.H.; Mahdavi, M.; Latifi-Navid, S. Helicobacter pylori vacA d1 genotype predicts risk of gastric adenocarcinoma and peptic ulcers in Northwestern Iran. Asian Pac. J. Cancer Prev. 2014, 15, 1575–1579. [Google Scholar] [CrossRef] [PubMed]
- Latifi-Navid, S.; Mohammadi, S.; Maleki, P.; Zahri, S.; Yazdanbod, A.; Siavoshi, F.; Massarrat, S. Helicobacter pylori vacA d1/-i1 genotypes and geographic differentiation between high and low incidence areas of gastric cancer in Iran. Arch. Iran. Med. 2013, 16, 330–337. [Google Scholar] [PubMed]
- Ladeira, M.S.; Bueno, R.C.; Dos Santos, B.F.; Pinto, C.L.; Prado, R.P.; Silveira, M.G.; Rodrigues, M.A.; Bartchewsky, W., Jr.; Pedrazzoli, J., Jr.; Ribeiro, M.L.; et al. Relationship among oxidative DNA damage, gastric mucosal density and the relevance of cagA, vacA and iceA genotypes of Helicobacter pylori. Dig. Dis. Sci. 2008, 53, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Sze, M.A.; Chen, Y.W.; Tam, S.; Tashkin, D.; Wise, R.A.; Connett, J.E.; Man, S.P.; Sin, D.D. The relationship between Helicobacter pylori seropositivity and COPD. Thorax 2015. [Google Scholar] [CrossRef] [PubMed]
- Kraus, J.; Nartova, E.; Pavlik, E.; Katra, R.; Sterzl, I.; Astl, J. Prevalence of Helicobacter pylori in adenotonsillar hypertrophy in children. Acta Oto-Laryngol. 2014, 134, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, S.; Kakugawa, T.; Yura, H.; Tomonaga, M.; Harada, T.; Hara, A.; Hara, S.; Nakano, M.; Yamasaki, E.; Sakamoto, N.; et al. Identification of Helicobacter pylori vacA in human lung and its effects on lung cells. Biochem. Biophys. Res. Commun. 2015, 460, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Santibanez, M.; Aguirre, E.; Belda, S.; Aragones, N.; Saez, J.; Rodriguez, J.C.; Galiana, A.; Sola-Vera, J.; Ruiz-Garcia, M.; Paz-Zulueta, M.; et al. Relationship between tobacco, cagA and vacA i1 virulence factors and bacterial load in patients infected by Helicobacter pylori. PLoS ONE 2015, 10, e0120444. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thi Huyen Trang, T.; Thanh Binh, T.; Yamaoka, Y. Relationship between vacA Types and Development of Gastroduodenal Diseases. Toxins 2016, 8, 182. https://doi.org/10.3390/toxins8060182
Thi Huyen Trang T, Thanh Binh T, Yamaoka Y. Relationship between vacA Types and Development of Gastroduodenal Diseases. Toxins. 2016; 8(6):182. https://doi.org/10.3390/toxins8060182
Chicago/Turabian StyleThi Huyen Trang, Tran, Tran Thanh Binh, and Yoshio Yamaoka. 2016. "Relationship between vacA Types and Development of Gastroduodenal Diseases" Toxins 8, no. 6: 182. https://doi.org/10.3390/toxins8060182
APA StyleThi Huyen Trang, T., Thanh Binh, T., & Yamaoka, Y. (2016). Relationship between vacA Types and Development of Gastroduodenal Diseases. Toxins, 8(6), 182. https://doi.org/10.3390/toxins8060182