
Gas Chromatography-Mass Spectrometry for Metabolite Profiling of Japanese Black Cattle Naturally Contaminated with Zearalenone and Sterigmatocystin

 ,
,
Abstract
:1. Introduction
2. Results
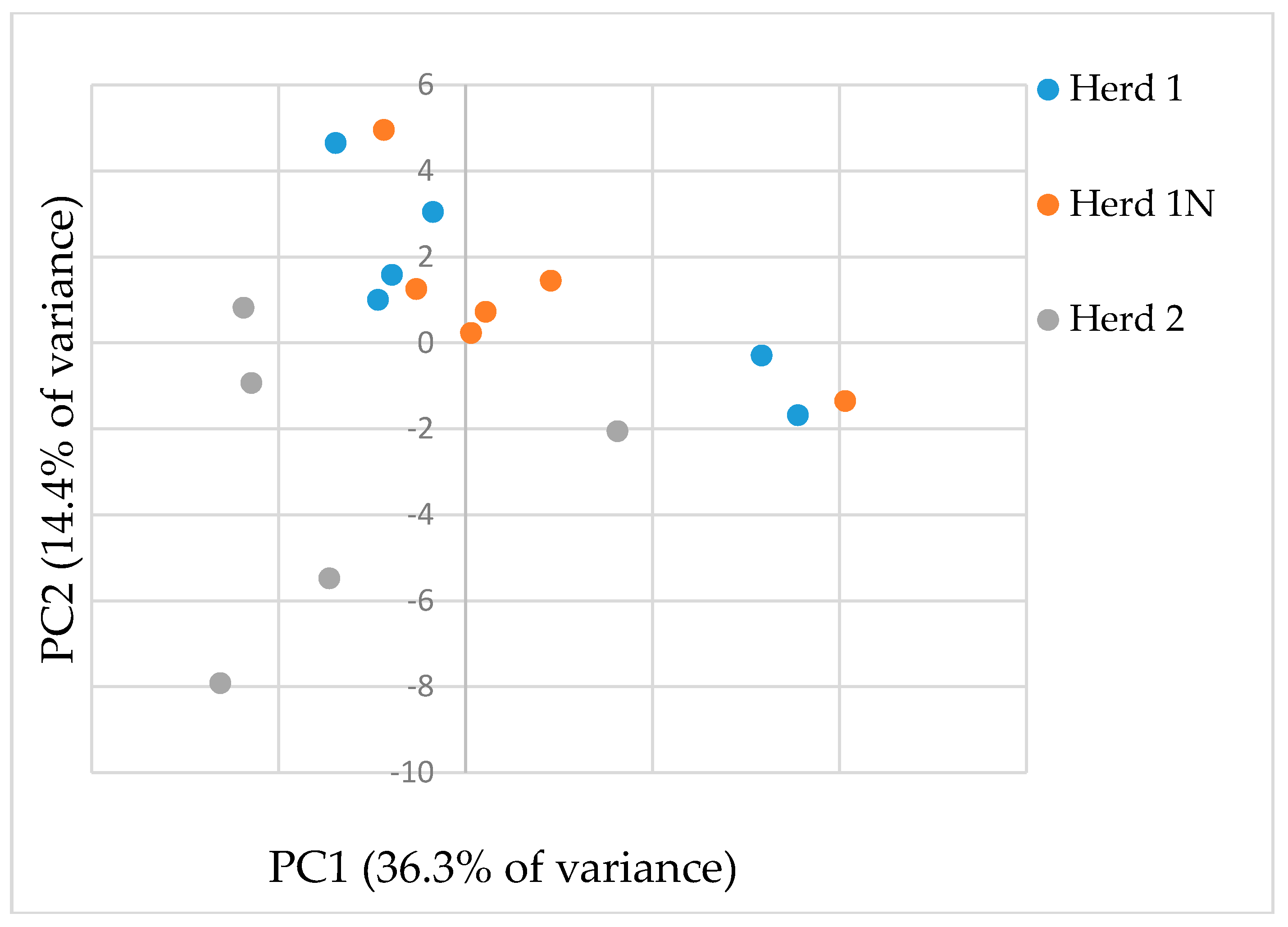
2.1. GC/MS Analysis of Urine Samples Derived from Cattle Herds
2.1.1. Original Chromatograms
2.1.2. Pattern Recognition
3. Discussion
3.1. Metabolism of ATP Generation
3.2. Amino Acid Metabolism
3.3. Glycinconjugation
3.4. Purine Base Metabolism
3.5. Metabolites Possibly Derived from Dietary Components
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Sample Collection and Processing
5.3. Preparation of Urine Samples for GC/MS Analyses
5.4. GC/MS Analysis and Creatinine Levels in the Urine
5.5. Data Processing
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Vet. J. 2008, 176, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yan, T.; Wang, J.; Huang, Z.; Chen, X.; Jia, G.; Wu, C.; Zhao, H.; Xue, B.; Xiao, L.; et al. Biological system responses to zearalenone mycotoxin exposure by integrated metabolomic studies. J. Agric. Food Chem. 2013, 61, 11212–11221. [Google Scholar] [CrossRef] [PubMed]
- Fushimi, Y.; Takagi, M.; Hasunuma, H.; Uno, S.; Kokushi, E.; Watanabe, U.; Liu, J.; Marey, M.; Miyamoto, A.; Otoi, T.; et al. Application of mycotoxin adsorbent to cattle feed naturally contaminated with zearalenone: Urinary zearalenone excretion and association with anti-Müllerian hormone. World Mycotoxin J. 2014, 7, 367–378. [Google Scholar] [CrossRef]
- Hasunuma, H.; Takagi, M.; Kawamura, O.; Taniguchi, C.; Nakamura, M.; Chuma, T.; Uno, S.; Kokushi, E.; Matsumoto, D.; Tshering, C.; et al. Natural contamination of dietary rice straw with zearalenone and urinary zearalenone concentrations in a cattle herd. J. Anim. Sci. 2012, 90, 1610–1616. [Google Scholar] [CrossRef] [PubMed]
- Fushimi, Y.; Takagi, M.; Uno, S.; Kokushi, E.; Nakamura, M.; Hasunuma, H.; Shinya, U.; Duguchi, E.; Fink-Gremmels, J. Measurement of sterigmatocystin concentrations in urine for monitoring the contamination of cattle feed. Toxins 2014, 6, 3117–3128. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Uno, S.; Kokushi, E.; Shiga, S.; Mukai, S.; Kuriyagawa, T.; Takagaki, K.; Hasunuma, H.; Matsumoto, D.; Okamoto, K.; et al. Measurement of urinary zearalenone concentrations for monitoring natural feed contamination in cattle herds—On farm trials. J. Anim. Sci. 2011, 89, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S. Metabolomics: The principles and potential applications to transplantation. Am. J. Transplant. 2005, 5, 2814–2820. [Google Scholar] [CrossRef] [PubMed]
- Madsen, R.; Lundstedt, T.; Trygg, J. Chemometrics in metabolomics- a review in human disease diagnosis. Anal. Chim. Acta 2010, 659, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Kind, T.; Tolstikov, V.; Fiehn, O.; Weiss, R.H. A comprehensive urinary metabolomic approach for identifying kidney cancer. Anal. Biochem. 2007, 363, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, S.; Gong, Y.; Mao, Y.; Liu, C.; Qu, H.; Cheng, Y. A metabonomic characterization of CCl4-induced acute liver failure using partial least square regression based on the GC/MS metabolic profiles of plasma in mice. J. Chromatogr. B 2008, 870, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Chorell, E.; Moritz, T.; Branth, S.; Antti, H.; Svensson, M.B. Predictive metabolomics evaluation of nutrition-modulated metabolic stress responses in human blood serum during the early recovery phase of strenuous physical exercise. J. Proteome. Res. 2009, 8, 2966–2977. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, H.; Kitoh, K.; Ohba, Y.; Kawashima, T.; Takahashi, H.; Yoshida, M.; Zhang, C.; Matsumoto, I. Urinary metabolic profiles of growth-retarded Japanese Black cattle by gas chromatography/mass spectrometry analysis. J. Jpn. Vet. Med. Assoc. 2003, 56, 445–449, (Japanese with English Abstract). [Google Scholar] [CrossRef]
- Bertram, H.C.; Yde, C.C.; Zhang, X.; Kristensen, N.B. Effect of dietary nitrogen content on the urine metabolite profile of dairy cows assessed by nuclear magnetic resonance (NMR)-based metabolomics. J. Agric. Food Chem. 2011, 59, 12499–12505. [Google Scholar] [CrossRef] [PubMed]
- Osorio, M.T.; Moloney, A.P.; Brennan, L.; Monahan, F.J. Authentication of beef production systems using a metabolomics-based approach. Animal 2012, 6, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Brand, B.; Hadlich, F.; Brandt, B.; Schauer, N.; Graunke, K.L.; Langgein, J.; Repsilber, D.; Ponsuksili, S.; Schwerin, M. Temperament type specific metabolite profiles of the prefrontal cortex and serum in cattle. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.Z.; Wang, D.M.; Wang, B.; Wang, J.K.; Liu, H.Y.; Guan, L.L.; Liu, J.X. Metabolomics of four biofluids from dairy cows: Potential biomarkers for milk production and quality. J. Proteome Res. 2015, 14, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Michell, A.W.; Mosedale, D.; Grainger, D.J.; Barker, R.A. Metabolomic analysis of urine and serum in Parkinson’s disease. Metabolomics 2008, 4, 191–201. [Google Scholar] [CrossRef]
- Vallejo, M.; García, A.; Tuñón, J.; García-Martínez, D.; Angulo, S.; Martin-Ventura, J.L.; Blanco-Colio, L.M.; Almeida, P.; Egido, J.; Barbas, C. Plasma fingerprinting with GC-MS in acute coronary syndrome. Anal. Bioanal. Chem. 2009, 394, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.; Groenke, K.; Takors, R.; Wandrey, C.; Oldiges, M. Simultaneous determination of multiple intracellular metabolites in glycolysis, pentose phosphate pathway and tricarboxylic acid cycle by liquid chromatography–mass spectrometry. J. Chromatogr. A 2007, 1147, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Bennett, B.D.; Rabinowitz, J.D. Analytical strategies for LCMS-based targeted metabolomics. J. Chromatogr. B 2008, 871, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Moraes, E.P.; Rupérez, F.J.; Plaza, M.; Herrero, M.; Barbas, C. Metabolomic assessment with CE-MS of the nutraceutical effect of Cystoseira spp extracts in an animal model. Electrophoresis 2011, 35, 2055–2062. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Zou, L.; Ong, C.N. Experiment-originated variations, and multi-peak and multi-origination phenomena in derivatizationbased GC-MS metabolomics. Trends Analyt. Chem. 2010, 29, 269–280. [Google Scholar] [CrossRef]
- Goodacre, R.; Vaidyanathan, S.; Dunn, W.B.; Harrigan, G.G.; Kell, D.B. Metabolomics by numbers: Acquiring and understanding global metabolite data. Trends Biotechnol. 2004, 22, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Seeling, K.; Lebzien, P.; Danicke, S.; Spilke, J.; Sudekum, K.H.; Flachowsky, G. Effects of level of feed intake and Fusarium toxin-contaminated wheat on rumen fermentation as well as on blood and milk parameters in cows. J. Anim. Physiol. Anim. Nutr. 2006, 90, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Wu, G.; He, Q.; Tang, H.; Wang, Y. The toxicity of acute exposure to T-2 toxin evaluated by the metabonomics technique. Mol. BioSyst. 2015, 11, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Uno, S.; Shintoyo, A.; Kokushi, E.; Yamamoto, M.; Nakayama, K.; Koyama, J. Gas chromatography-mass spectrometry for metabolite profiling of Japanese medaka (Oryzias latipes) juveniles exposed to melathion. Environ. Sci. Pollut. Res. 2012, 19, 2595–2605. [Google Scholar] [CrossRef] [PubMed]
- Kokushi, E.; Shintoyo, A.; Koyama, J.; Uno, S. Evaluation of 2,4-dichlorophenol exposure of Japanese medaka, Oryzias latipes, using a metabolomics approach. Environ. Sci. Pollut. Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.R.; Fink-Gremmels, J. Mycotoxin syndrome in dairy cattle: characterization and intervention results. World Mycotoxin J. 2014, 7, 357–366. [Google Scholar] [CrossRef]
- Saha, B.C. Hemicellulose bioconversion. J. Ind. Microbiol. Biotechnol. 2003, 30, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Conkova, E.; Laciakova, A.; Pastorova, B.; Seidel, H.; Kovac, G. The effect of zearalenone on some enzymatic parameters in rabbits. Toxicol. Lett. 2001, 121, 145–149. [Google Scholar] [CrossRef]
- Lasram, M.M.; Dhouib, I.B.; Annabi, A.; El Fazaa, S.; Gharbi, N. A review on the molecular mechanisms involved in insulin resistance induced by organophosphorus pesticides. Toxicilogy 2014, 322, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ye, Y.; An, Y.; Tian, Y.; Wang, Y.; Tang, H. Systems responses of rats to aflatoxin B1 exposure revealed with metabonomic changes in multiple biological matrices. J. Proteome Res. 2010, 10, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Novelli, G.D.; Kaplan, N.O.; Lipmann, F. The liberation of pantothenic acid from coenzyme A. J. Biol. Chem. 1949, 177, 97–107. [Google Scholar] [PubMed]
- Fisher, S.K.; Heacock, A.M.; Agranoff, B.W. Inositol lipids and signal transduction in the nervous system: An update. J. Neurochem. 1992, 58, 18–38. [Google Scholar] [CrossRef] [PubMed]
- Landaas, S. Increased urinary excretion of 3-hydroxyisovaleric acid in patients with ketoacidosis. Clin. Chim. Acta 1974, 54, 39–46. [Google Scholar] [CrossRef]
- Abid-Essefi, S.; Quanes, Z.; Hassen, W.; Baudrimont, L.; Creppy, E.; Bacha, H. Cytotoxicity, inhibition of DNA and protein syntheses and oxidative damage in cultured cells exposed to zearalenone. Toxicol. In Vitro 2004, 18, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Schlessinger, J. Cell signaling by receptor tyrosine kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.N.; Rulquin, H.; Andrade, A.; Faverdin, P.; Peyraud, J.L.; Lemosquet, S. Milk proteins synthesis in response to the provision of an “ideal” amino acid profile at 2 levels of metabolizable protein supply in dairy cows. J. Dairy Sci. 2012, 95, 5876–5887. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.L.; Miller, G.M. Advances in tryptophan hydroxylase-2 gene expression regulation: New insights into serotonin-stress interaction and clinical implication. Am. J. Med. Genet. B Neuropsychiatr Genet. B 2012, 159, 152–171. [Google Scholar] [CrossRef] [PubMed]
- Hale, M.W.; Shekhar, A.; Lowry, C.A. Development by environment interactions controlling tryptophan hydroxylase expression. J. Chem. Neuroanat. 2011, 41, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Boudra, H.; Doreau, M.; Noziere, P. Simultaneous analysis of the main markers of nitrogen status in dairy cow’s urine using hydrophilic interaction chromatography and tandem mass spectrometry detection. J. Chromatogr. A 2012, 1256, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Bomalaski, J.S.; Clark, M.A. Serum uric acid-lowering therapies: where are we heading in management of hyperuricemia and the potential role of uricase. Curr. Rheumat. Rep. 2004, 6, 240–247. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Offer, N.W. Evaluation of the spot urine sampling technique to assess urinary pirine derivates excretion in lactating dairy cows. Anim. Sci. 1998, 66, 557–568. [Google Scholar] [CrossRef]
- Chen, X.B.; Mejia, A.T.; Kyle, D.J.; Orskov, E.R. Evaluation of the use of purine derivative-creatinine ratio in spot urine and plasma samples as an index of microbial protein supply in ruminants-studies in sheep. J. Agric. Sci. 1995, 125, 137–143. [Google Scholar] [CrossRef]
- Shingfield, K.J. Estimation of microbial protein supply in ruminant animals based on renal and mammary purine excretion: A review. J. Anim. Feed Sci. 2000, 9, 169–212. [Google Scholar] [CrossRef]
- Sharma, R.P.; Olson, L.E.; Stowe, C.M. Excretion of benzoate in bovine urine after the administration of thiopental. Biochem. Pharmacol. 1972, 21, 181–191. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Metabolite | PC1 | PC2 | p Value | |
|---|---|---|---|---|
| ATP generation | xylitol | 0.88 | 0.12 | |
| pantothenic acid | 0.79 | 0.63 | ||
| galactose | 0.59 | 0.79 | ||
| myo-inositol | 0.56 | 0.51 | ||
| aconitic acid | 0.54 | 0.55 | ||
| glucose | 0.53 | 0.78 | ||
| isocitric acid | 0.53 | 0.50 | ||
| lactate | −0.87 | 0.038 | ||
| lactose | −0.84 | 0.03 | ||
| lyxose | −0.59 | 0.47 | ||
| xylitol | −0.50 | 0.80 | ||
| Organic acid | hydroxyisovaleric acid | 0.64 | 0.48 | |
| methyl succinic acid | 0.66 | 0.32 | ||
| ethylhydracrylic acid | −0.69 | 0.19 | ||
| phenaceturic acid | −0.63 | 0.35 | ||
| Amino acid | threonine | 0.78 | 0.08 | |
| lysine | 0.64 | 0.31 | ||
| taurine | 0.52 | 0.028 | ||
| tyrosine | 0.86 | 0.36 | ||
| oxoproline (pyroglutamic acid) | 0.70 | 0.38 | ||
| tryptophan | 0.74 | 0.32 | ||
| phosphocolamine | 0.73 | 0.0008 | ||
| serine | 0.62 | −0.61 | 0.09 | |
| Glycin-conjugate | butyrylglycine | 0.76 | 0.70 | |
| hippurate | 0.75 | 0.66 | ||
| methylbutyrylglycine | 0.62 | 0.82 | ||
| Purine base | ribofuranose | 0.76 | 0.003 | |
| allantoin | −0.56 | 0.011 | ||
| β-pseudouridine | 0.85 | 0.87 | ||
| uric acid | −0.60 | −0.58 | 0.23 | |
| Dietary plant-derived | adonitol | 0.88 | 0.87 | |
| threitol | 0.88 | 0.87 | ||
| cinnamate | 0.82 | 0.30 | ||
| hydroxyphenyllactate | 0.82 | 0.48 | ||
| glycolic acid | 0.56 | −0.48 | 0.75 | |
| indol-3-acetic acid | 0.54 | 0.54 | ||
| galacturonic acid | −0.66 | 0.002 | ||
| hippuric acid | −0.67 | 0.0002 | ||
| deoxytetronic acid | 0.95 | 0.53 | ||
| threonic acid | 0.90 | 0.68 | ||
| gluconic acid | 0.88 | 0.77 |
| Forage Feed, kg | Formula Feed | |||||||
|---|---|---|---|---|---|---|---|---|
| Herd | Total, kg | Bran, % | Cereal, % | Oil Seed Meal, % | Other, % | TDN, % | CP, % | |
| Herd 1 | Straw, 2 | 9 | 17 | 77 | 5 | 1 | >75 | >14 |
| Herd 2 | Straw, 4 | 9 | 17 | 77 | 5 | 1 | >75 | >14 |
| Experimental Group | Straw (mg/kg) | Urine Samples (Mean ± SEM; pg/mg Creatinine) | ||||
|---|---|---|---|---|---|---|
| ZEN | STC | ZEN (n = 6) | α-ZOL (n = 6) | β-ZOL (n = 6) | STC (n = 6) | |
| Herd 1 | 7.6 | 0.24 | 3702 ± 747 | 859 ± 178 | 5503 ± 1130 | 569 ± 111 |
| (−42 days) * | 7.5 | 0.17 | 2444 ± 394 | 759 ± 148 | 5496 ± 1197 | 209 ± 22 |
| Herd 1N | ND | 0.04 | 63 ± 12 | 20 ± 3 | 119 ± 18 | 33 ± 8 |
| Herd 2 | 0.2 | <0.01 | 50 ± 8 | ND | 82 ± 27 | 47 ± 19 |
| (−34 days) ** | 0.2 | 0.03 | 68 ± 10 | 21 ± 9 | 215 ± 51 | 147 ± 39 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toda, K.; Kokushi, E.; Uno, S.; Shiiba, A.; Hasunuma, H.; Fushimi, Y.; Wijayagunawardane, M.P.B.; Zhang, C.; Yamato, O.; Taniguchi, M.; et al. Gas Chromatography-Mass Spectrometry for Metabolite Profiling of Japanese Black Cattle Naturally Contaminated with Zearalenone and Sterigmatocystin. Toxins 2017, 9, 294. https://doi.org/10.3390/toxins9100294
Toda K, Kokushi E, Uno S, Shiiba A, Hasunuma H, Fushimi Y, Wijayagunawardane MPB, Zhang C, Yamato O, Taniguchi M, et al. Gas Chromatography-Mass Spectrometry for Metabolite Profiling of Japanese Black Cattle Naturally Contaminated with Zearalenone and Sterigmatocystin. Toxins. 2017; 9(10):294. https://doi.org/10.3390/toxins9100294
Chicago/Turabian StyleToda, Katsuki, Emiko Kokushi, Seiichi Uno, Ayaka Shiiba, Hiroshi Hasunuma, Yasuo Fushimi, Missaka P. B. Wijayagunawardane, Chunhua Zhang, Osamu Yamato, Masayasu Taniguchi, and et al. 2017. "Gas Chromatography-Mass Spectrometry for Metabolite Profiling of Japanese Black Cattle Naturally Contaminated with Zearalenone and Sterigmatocystin" Toxins 9, no. 10: 294. https://doi.org/10.3390/toxins9100294
APA StyleToda, K., Kokushi, E., Uno, S., Shiiba, A., Hasunuma, H., Fushimi, Y., Wijayagunawardane, M. P. B., Zhang, C., Yamato, O., Taniguchi, M., Fink-Gremmels, J., & Takagi, M. (2017). Gas Chromatography-Mass Spectrometry for Metabolite Profiling of Japanese Black Cattle Naturally Contaminated with Zearalenone and Sterigmatocystin. Toxins, 9(10), 294. https://doi.org/10.3390/toxins9100294