
Responses of Microcystis Colonies of Different Sizes to Hydrogen Peroxide Stress

Abstract
:1. Introduction
2. Results
2.1. Degradation of H2O2
2.2. Chlorophyll-a Content
2.3. Effects of H2O2 on Photosynthetic Activity
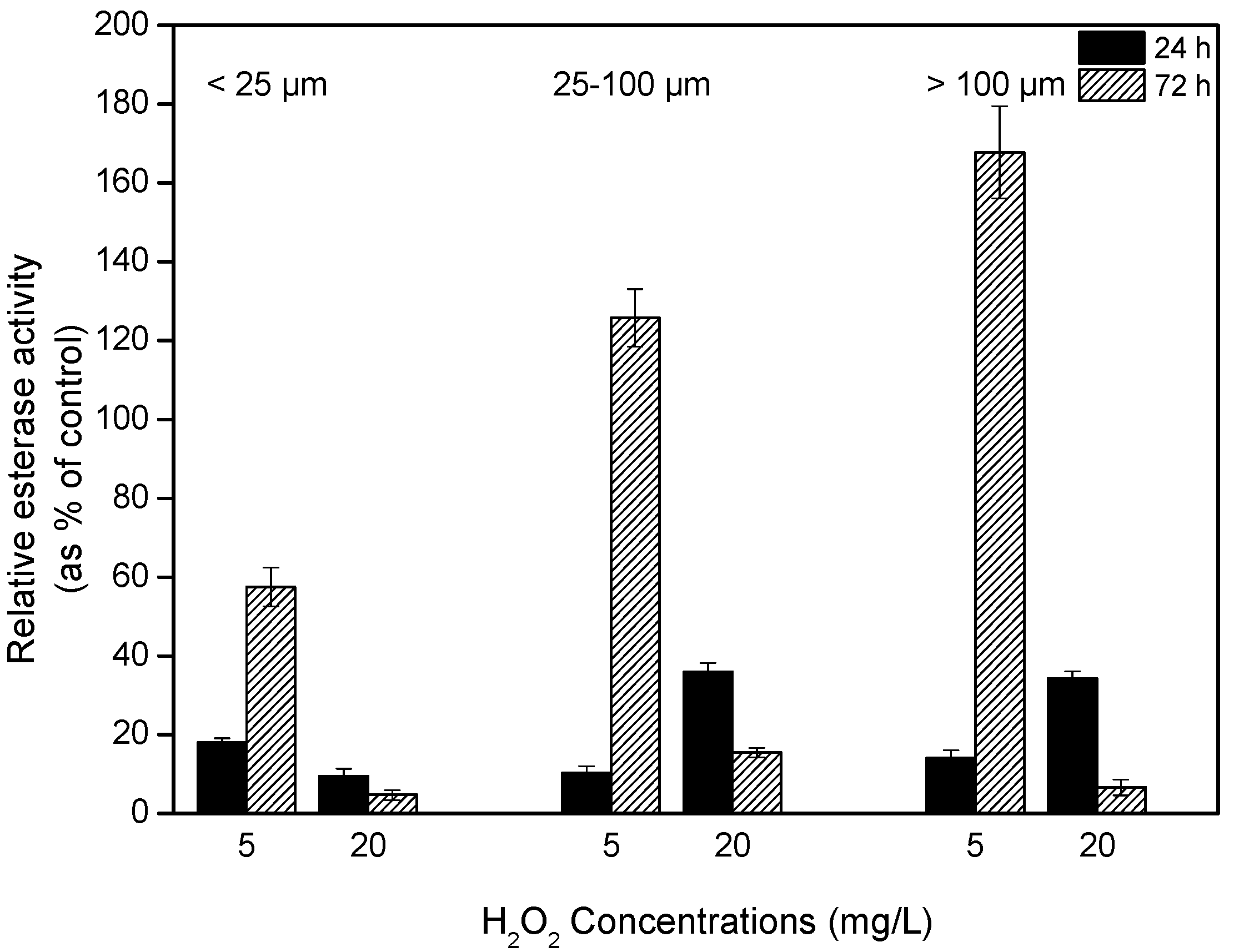
2.4. Effects of H2O2 on Esterase Activity
2.5. Antioxidant Responses
2.5.1. SOD and CAT Activities
2.5.2. GSH and MDA Contents
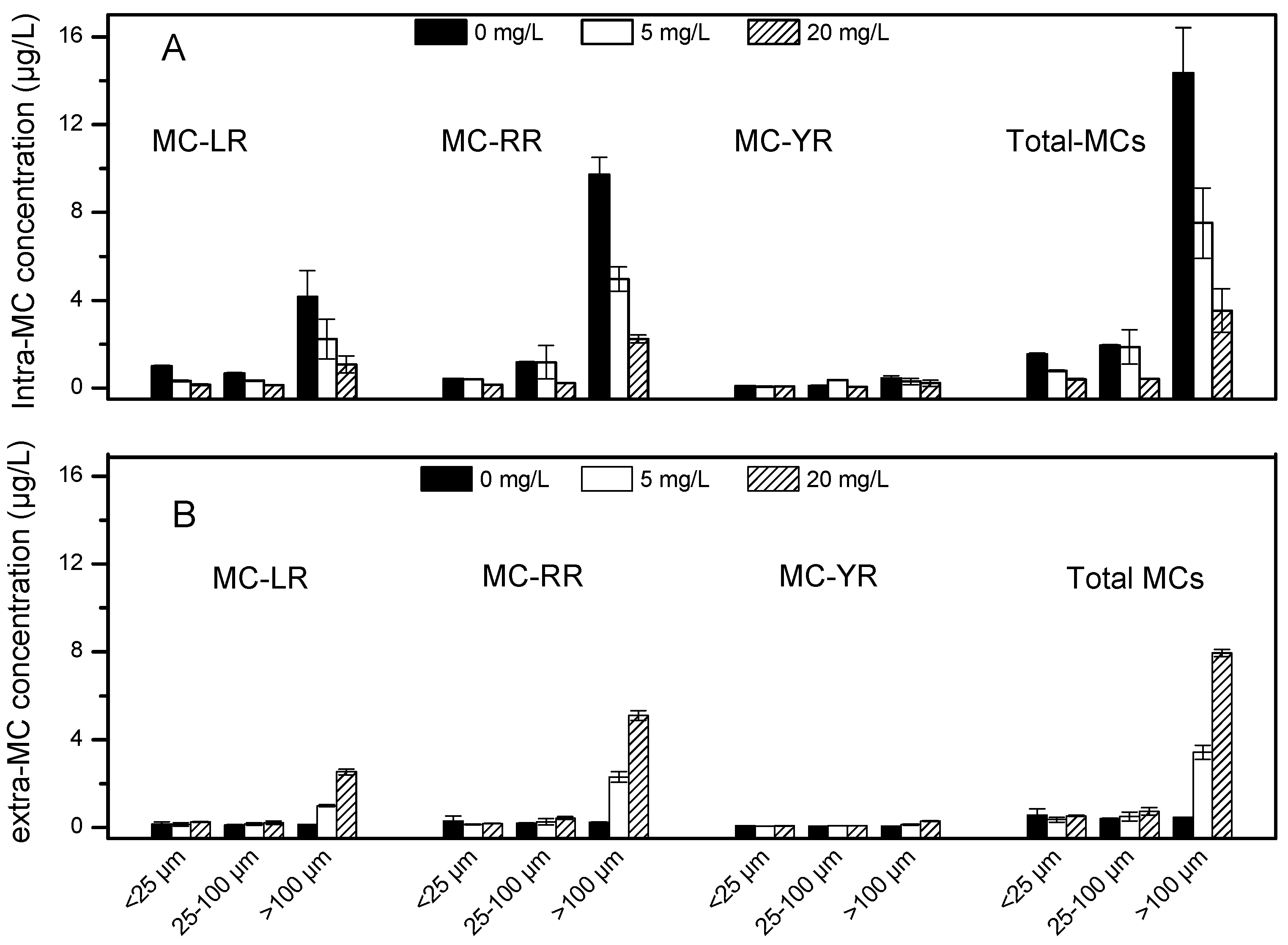
2.6. Microcystin Concentrations
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sampling
5.2. Experimental Procedures
5.3. Analytical Methods
5.3.1. Measurement of H2O2
5.3.2. Measurement of Chlorophyll-a
5.3.3. Photosynthesis Measurements
5.3.4. Esterase Activity Measurement
5.3.5. Array of Antioxidant Responses
5.3.6. Microcystin Analysis
5.3.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dokulil, M.T.; Teubner, K. Cyanobacterial dominance in lakes. Hydrobiologia 2000, 438, 1–12. [Google Scholar] [CrossRef]
- Paerl, H.W.; Fulton, R.S.; Moisander, P.H.; Dyble, J. Harmful freshwater algal blooms, with an emphasis on cyanobacteria. Sci. World J. 2001, 1, 76–113. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.X.; Gao, G. Hypothesis on cyanobacteria bloom-forming mechanism in large shallow eutrophic lakes. Acta Ecol. Sin. 2005, 25, 589–595. [Google Scholar]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Niedermeyer, T. Microcystin Congeners Described in the Literature. Available online: http://dx.doi.org/10.6084/m9.figshare.880756 (accessed on 21 October 2014).
- Rodríguez, E.M.; Acero, J.L.; Spoof, L.; Meriluoto, J. Oxidation of MC-LR and -RR with chlorine and potassium permanganate: Toxicity of the reaction products. Water Res. 2008, 42, 1744–1752. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Kong, F.; Zhang, M.; Yang, Z.; Shi, X.L.; Du, M.Y. The dynamics of Microcystis genotypes and microcystin production and associations with environmental factors during blooms in Lake Chaohu, China. Toxins 2014, 6, 3238–3257. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.H.; Yang, Z.; Su, W.; Johnson, D.; Kong, F.X. Controlling of cyanobacteria bloom during bottleneck stages of algal cycling in shallow Lake Taihu (China). J. Freshw. Ecol. 2013, 29, 129–140. [Google Scholar] [CrossRef]
- Huo, X.C.; Chang, D.W.; Tseng, J.H.; Burch, M.D.; Lin, T.F. Exposure of Microcystis aeruginosa to Hydrogen Peroxide under Light: Kinetic Modeling of Cell Rupture and Simultaneous Microcystin Degradation. Environ. Sci. Technol. 2015, 49, 5502–5510. [Google Scholar] [CrossRef] [PubMed]
- Drábková, M.; Matthijs, H.C.P.; Admiraal, W.; Maršálek, B. Selective effects of H2O2 on cyanobacterial photosynthesis. Photosynthetica 2007, 45, 363–369. [Google Scholar] [CrossRef]
- Drábková, M.; Admiraal, W.; Maršálek, B. Combined exposure to hydrogen peroxide and light - selective effects on cyanobacteria, green algae and diatoms. Environ. Sci. Technol. 2007, 41, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.C.; Li, D.H.; Qin, H.J.; Li, Y.X. An integrated method for removal of harmful cyanobacterial blooms in eutrophic lakes. Environ. Pollut. 2012, 160, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Pan, X.L.; Zhang, D.Y.; Mu, S.Y.; Lee, D.J.; Halik, U. Extracellular polymeric substances buffer against the biocidal effect of H2O2 on the bloom-forming cyanobacterium Microcystis aeruginosa. Water Res. 2015, 69, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.X.; Gan, N.Q.; Huang, Q.; Song, L.R. Response of Microcystis to copper stress: Do phenotypes of Microcystis make a difference in stress tolerance. Environ. Pollut. 2007, 147, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Li, D.H. Physiological variations of bloom-forming Microcystis (Cyanophyceae) related to colony size changes during blooms. Phycologia 2012, 51, 599–603. [Google Scholar] [CrossRef]
- Dummermuth, A.L.; Karsten, U.; Fisch, K.M.; König, G.M.; Wiencke, C. Responses of marine macroalgae to hydrogen-peroxide stress. J. Exp. Mar. Biol. Ecol. 2003, 289, 103–121. [Google Scholar] [CrossRef] [Green Version]
- Matthijs, H.C.P.; Visser, P.M.; Reeze, B.; Meeuse, J.; Slot, P.C.; Wijn, G.; Talens, R.; Huisman, J. Selective suppression of harmful cyanobacteria in an entire lake with hydrogen peroxide. Water Res. 2012, 46, 1460–1472. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Hu, H.Y.; Xie, X.; Li, F.M. Responses of enzymatic antioxidants and non-enzymatic antioxidants in the cyanobacterium Microcystis aeruginosa to the allelochemical ethyl 2-methyl acetoacetate (EMA) isolated from reed (Phragmites communis). J. Plant Physiol. 2008, 165, 1264–1273. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.L.; Zhang, B.; Dai, W.; Zhang, X.M. Oxidative damage and antioxidant responses in Microcystis aeruginosa exposed to the allelochemical berberine isolated from golden thread. J. Plant Physiol. 2011, 168, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.Q.; Shao, Y.S.; Gao, N.Y.; Deng, Y.; Qiao, J.L.; Ou, H.; Deng, J. Effects of different algaecides on the photosynthetic capacity, cell integrity and microcystin-LR release of Microcystis aeruginosa. Sci. Total Environ. 2013, 463–464, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Lurling, M.; Meng, D.; Faassen, E.J. Effects of hydrogen peroxide and ultrasound on biomass reduction and toxin release in the cyanobacterium, Microcystis aeruginosa. Toxins 2014, 6, 3260–3280. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Chen, C.; Ding, J.Q.; Hou, A.X.; Li, Y.; Niu, Z.B.; Su, X.Y.; Xu, Y.J.; Laws, E.A. The 2007 water crisis in Wuxi, China: Analysis of the origin. J. Hazard. Mater. 2010, 182, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, V.; Thakkar, M.; Wei, L.P. Effect of Aureococcus anophagefferens biomass on its inactivation by hydrogen peroxide: Culture study and empirical modeling. J. Appl. Phycol. 2013, 26, 349–355. [Google Scholar] [CrossRef]
- Chen, C.; Yang, Z.; Kong, F.X.; Zhang, M.; Yu, Y.; Shi, X.L. Growth, physiochemical and antioxidant responses of overwintering benthic cyanobacteria to hydrogen peroxide. Environ. Pollut. 2016, 219, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.J.; Ho, L.; Hobson, P.; Brookes, J. Evaluating the effectiveness of copper sulphate, chlorine, potassium permanganate, hydrogen peroxide and ozone on cyanobacterial cell integrity. Water Res. 2013, 47, 5153–5164. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Kong, F.X.; Shi, X.L.; Cao, H.S. Morphological response of Microcystis aeruginosa to grazing by different sorts of zooplankton. Hydrobiologia 2006, 563, 225–230. [Google Scholar] [CrossRef]
- Veldhuis, M.J.W.; Admiraal, W. Influence of phosphate depletion on the growth and colony formation of Phaeocystispouchetii. Mar. Biol. 1987, 95, 47–54. [Google Scholar] [CrossRef]
- Park, M.H.; Chung, I.M.; Ahmad, A.; Kim, B.H.; Hwang, S.J. Growth inhibition of unicellular and colonial Microcystis strains (Cyanophyceae) by compounds isolated from rice (Oryza sativa) hulls. Aquat. Bot. 2009, 90, 309–314. [Google Scholar] [CrossRef]
- Li, M.; Zhu, W.; Dai, X.X.; Li, X.Y. Effects of linear alkylbenzene sulfonate on extracellular polysaccharide content and cells per particle of Microcystis aeruginosa and Scenedesmus obliquus. Fresen. Environ. Bull. 2013, 22, 1189–1194. [Google Scholar]
- Liu, Y.; Guan, Y.T.; Gao, B.Y.; Yue, Q.Y. Antioxidant responses and degradation of two antibiotic contaminants in Microcystis aeruginosa. Ecotoxicol. Environ. Saf. 2012, 86, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Gomez, L.; Vanacker, H.; Foyer, C.H. Interactions between biosynthesis, compartmentation and transport in the control of glutathione homeostasis and signalling. J. Exp. Bot. 2002, 53, 1283–1304. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R.; Christiansen, G.; Chorus, I. The Abundance of Microcystin-Producing Genotypes Correlates Positively with Colony Size in Microcystis sp. and Determines Its Microcystin Net Production in Lake Wannsee. Appl. Environ. Microbiol. 2003, 69, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Borner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of Microcystis under oxidative stress conditions. PLoS ONE 2011, 6, e17615. [Google Scholar] [CrossRef] [PubMed]
- Dziallas, C.; Grossart, H.P. Increasing Oxygen Radicals and Water Temperature Select for Toxic Microcystis sp. PLoS ONE 2011, 6, e25569. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.W.; Qin, B.Q.; Teubner, K.; Dokulil, M.T. Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China. J. Plankton Res. 2003, 25, 445–453. [Google Scholar] [CrossRef]
- Qin, B.Q.; Li, W.; Zhu, G.W.; Zhang, Y.L.; Wu, T.F.; Gao, G. Cyanobacterial bloom management through integrated monitoring and forecasting in large shallow eutrophic Lake Taihu (China). J. Hazard. Mater. 2015, 287, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.P.; Lin, C.T.; Chang, C.M.; Wu, S.H.; Lo, L.C. Nitrophenylboronic Acids as Highly Chemoselective Probes To Detect Hydrogen Peroxide in Foods and Agricultural Products. J. Agric. Food Chem. 2011, 59, 11403–11406. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.S.; Kong, F.X.; Tan, J.K.; Zhang, X.F.; Tao, Y.; Yang, Z. Recruitment of Total Phytoplankton, Chlorophytes and Cyanobacteria from Lake Sediments Recorded by Photosynthetic Pigments in a Large, Shallow Lake (Lake Taihu, China). Int. Rev. Hydrobiol. 2005, 90, 347–357. [Google Scholar] [CrossRef]
- Yan, R.; Kong, F.X.; Han, X.B. Analysis of the Recruitment of the Winter Survival Algae on the Sediments of Lake Taihu by Fluorometry. J. Lake Sci. 2004, 16, 163–168. [Google Scholar]
- Yang, Z.; Kong, F.X.; Shi, X.L.; Yu, Y.; Zhang, M. Effects of UV-B radiation on microcystin production of a toxic strain of Microcystis aeruginosa and its competitiveness against a non-toxic strain. J. Hazard. Mater. 2015, 283, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yu, Y.; Yang, Z.; Kong, F.X. Photochemical responses of phytoplankton to rapid increasing-temperature process. Phycol. Res. 2012, 60, 199–207. [Google Scholar] [CrossRef]
- Latour, D.; Sabido, O.; Salenc, M.J.; Giraudet, H. Dynamics and metabolic activity of the benthic cyanobacterium Microcystis aeruginosa in the Grangent reservoir (France). J. Plankton Res. 2004, 26, 719–726. [Google Scholar] [CrossRef]
- Yu, Y.; Kong, F.X.; Wang, M.L.; Qian, L.L.; Shi, X.L. Determination of short-term copper toxicity in a multispecies microalgal population using flow cytometry. Ecotoxicol. Environ. Saf. 2007, 66, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Wayne, F.; Beyer, J.R.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar]
- Goth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–152. [Google Scholar] [CrossRef]
- Dogru, M.I.; Dogru, A.K.; Gul, M.; Esrefoglu, M.; Yurekli, M.; Erdogan, S.; Ates, B. The effect of adrenomedullin on rats exposed to lead. J. Appl. Toxicol. 2008, 28, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | H2O2 | Chlorophyll-a | Fv/Fm | Fv’/Fm’ | FDA | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Effects | F | p-Value | F | p-Value | F | p-Value | F | p-Value | F | p-Value |
| Time | 8832.23 | <0.01 * | 638.07 | <0.01* | 0.98 | 0.37 | 21.79 | <0.01 * | 274.71 | <0.01 * |
| Time * Concentration | 4492.83 | <0.01 * | 149.05 | <0.01* | 16.33 | <0.01 * | 21.16 | <0.01 * | 110.02 | <0.01 * |
| Time × Size | 0.47 | 0.83 | 5.63 | <0.01* | 6.26 | <0.01 * | 5.19 | <0.01 * | 23.06 | <0.01 * |
| Concentration | 9942.51 | <0.01 * | 2562.62 | <0.01* | 987.99 | <0.01 * | 953.76 | <0.01 * | 267.84 | <0.01 * |
| Size | 1.42 | 0.26 | 617.68 | <0.01* | 96.23 | <0.01 * | 98.72 | <0.01 * | 21.23 | <0.01 * |
| Concentration × Size | 1.10 | 0.38 | 45.06 | <0.01* | 42.08 | <0.01 * | 40.36 | <0.01 * | 17.91 | <0.01 * |
| Variable | SOD | CAT | GSH | MDA | ||||||
| Effects | F | p-Value | F | p-Value | F | p-Value | F | p-Value | ||
| Time | 17.97 | <0.01 * | 114.99 | <0.01 * | 102.51 | <0.01 * | 71.91 | <0.01* | ||
| Time × Concentration | 5.67 | <0.01 * | 25.73 | <0.01 * | 24.37 | <0.01 * | 8.90 | <0.01* | ||
| Time × Size | 21.96 | <0.01 * | 28.88 | <0.01 * | 40.28 | <0.01 * | 29.88 | <0.01* | ||
| Concentration | 170.31 | <0.01 * | 113.85 | <0.01 * | 225.26 | <0.01 * | 330.40 | <0.01* | ||
| Size | 70.11 | <0.01 * | 196.84 | <0.01 * | 469.17 | <0.01 * | 160.14 | <0.01* | ||
| Concentration × Size | 31.39 | <0.01 * | 11.45 | <0.01 * | 179.12 | <0.01 * | 44.76 | <0.01* | ||
| Variable | H2O2 Concentration | Colony Size | ||||
|---|---|---|---|---|---|---|
| 0 mg/L | 5 mg/L | 20 mg/L | <25 μm | 25–100 μm | >100 μm | |
| Intracellular-MCLR | a | b | c | a | a | b |
| Intracellular-MCRR | a | b | c | a | b | c |
| Intracellular-MCYR | a | a | b | a | b | c |
| Intracellular-MCs | a | b | c | a | a | b |
| Extracellular-MCLR | a | b | c | a | a | b |
| Extracellular-MCRR | a | b | c | a | a | b |
| Extracellular-MCYR | a | b | c | a | a | b |
| Extracellular-MCs | a | b | c | a | a | b |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Shi, X.; Chen, C.; Yu, L.; Sun, C. Responses of Microcystis Colonies of Different Sizes to Hydrogen Peroxide Stress. Toxins 2017, 9, 306. https://doi.org/10.3390/toxins9100306
Liu M, Shi X, Chen C, Yu L, Sun C. Responses of Microcystis Colonies of Different Sizes to Hydrogen Peroxide Stress. Toxins. 2017; 9(10):306. https://doi.org/10.3390/toxins9100306
Chicago/Turabian StyleLiu, Mixue, Xiaoli Shi, Chao Chen, Li Yu, and Chuang Sun. 2017. "Responses of Microcystis Colonies of Different Sizes to Hydrogen Peroxide Stress" Toxins 9, no. 10: 306. https://doi.org/10.3390/toxins9100306
APA StyleLiu, M., Shi, X., Chen, C., Yu, L., & Sun, C. (2017). Responses of Microcystis Colonies of Different Sizes to Hydrogen Peroxide Stress. Toxins, 9(10), 306. https://doi.org/10.3390/toxins9100306