
Combined Venom Gland Transcriptomic and Venom Peptidomic Analysis of the Predatory Ant Odontomachus monticola


Abstract
:1. Introduction
2. Results and Discussion
2.1. Ilumina Hiseq 2500 Deep RNA Sequencing, Assembly, and Annotation
2.2. Toxin-Like Peptides and Proteins
2.2.1. Pilosulin-Like Peptides
2.2.2. Chitinase-Like Protein
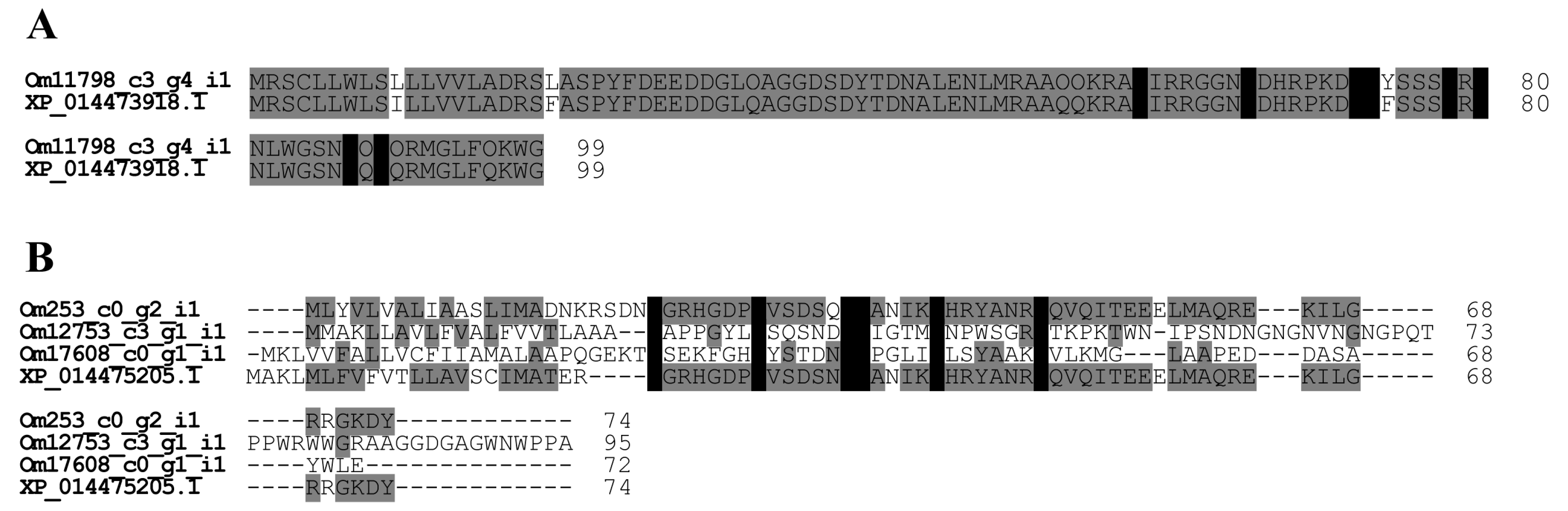
2.2.3. Icarapin-Like Peptide
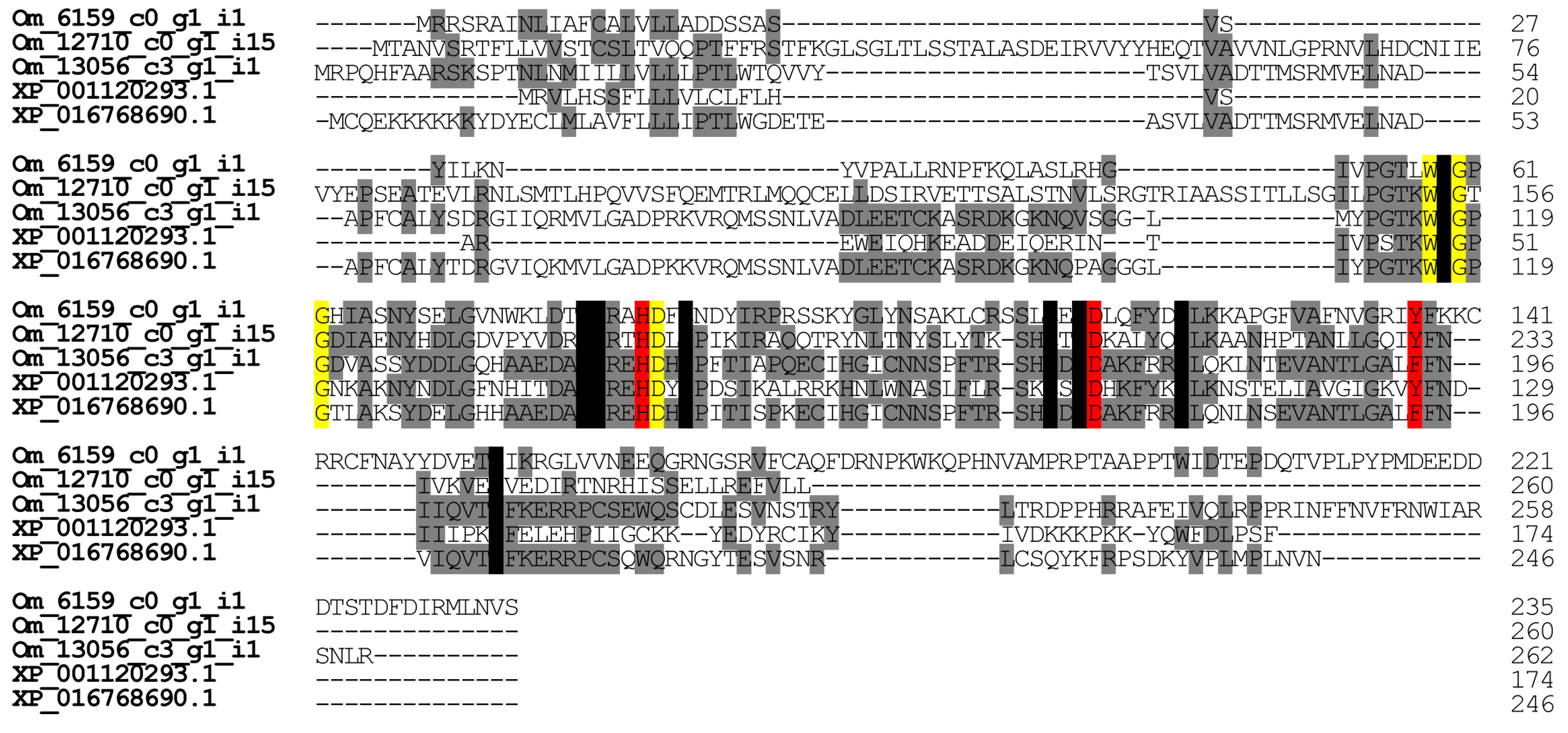
2.2.4. Hyaluronidase
2.2.5. ICK-Like Peptides
2.2.6. Dipeptidyl Peptidase 4
2.2.7. Phospholipase A2-Like Proteins
2.2.8. Other Low-Abundance Transcripts of Toxin-Like Peptides and Proteins
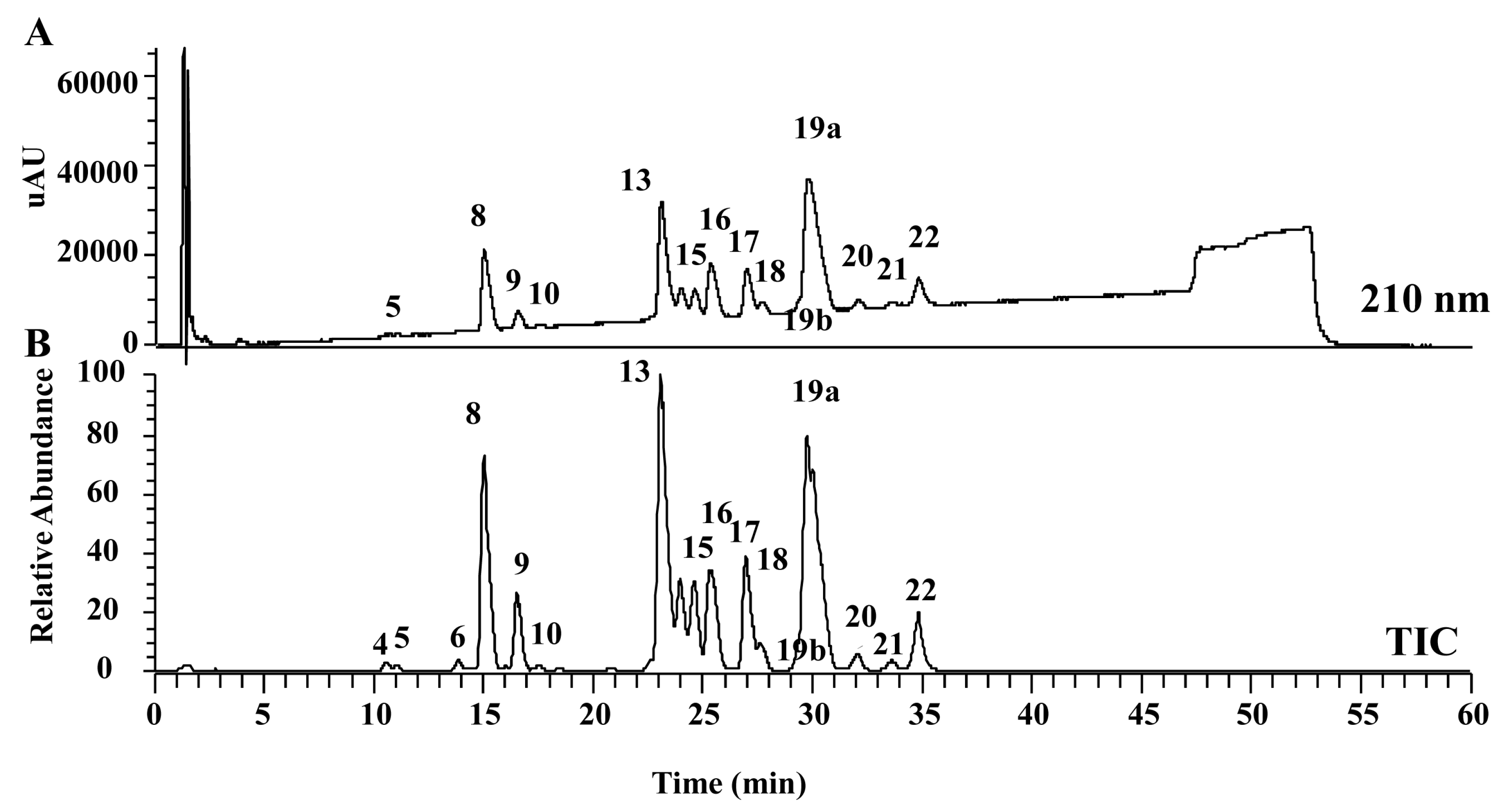
2.3. Peptidomic Analysis
3. Conclusions
4. Materials and Methods
4.1. Ants
4.2. Transcriptome Analysis
4.3. Pilosulin-Like Peptide 1 cDNA Cloning and Sequencing
4.4. Liquid Chromatography-Mass Spectrometry (LC-MS) Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Inagaki, H.; Akagi, M.; Imai, H.T.; Taylor, R.W.; Kubo, T. Molecular cloning and biological characterization of novel antimicrobial peptides, pilosulin 3 and pilosulin 4, from a species of the Australian ant genus Myrmecia. Arch. Biochem. Biophys. 2004, 428, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, H.; Akagi, M.; Imai, H.T.; Taylor, R.W.; Wiese, M.D.; Davies, N.W.; Kubo, T. Pilosulin 5, a novel histamine-releasing peptide of the Australian ant, Myrmecia pilosula (Jack Jumper Ant). Arch. Biochem. Biophys. 2008, 477, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, H.; Masuko, K.; Kudo, T. SKTXs: Peptides identified from the ant Strumigenys kumadori that block sodium channels. In Proceedings of the 8th-Asia-Pacific Congress on Animal, Plant and Microbial Toxins, Hanoi, Vietnam, 2–6 December 2008; p. 75. [Google Scholar]
- Torres, A.F.; Huang, C.; Chong, C.M.; Leung, S.W.; Prieto-da-Silva, A.R.; Havt, A.; Quinet, Y.P.; Martins, A.M.; Lee, S.M.; Radis-Baptista, G. Transcriptome analysis in venom gland of the predatory giant ant Dinoponera quadriceps: Insights into the polypeptide toxin arsenal of hymenopterans. PLoS ONE 2014, 9, e87556. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, W.; Verdenaud, M.; Klopp, C.; Ducancel, F.; Noirot, C.; Vetillard, A. De Novo sequencing and transcriptome analysis for Tetramorium bicarinatum: A comprehensive venom gland transcriptome analysis from an ant species. BMC Genom. 2014, 15, 987. [Google Scholar] [CrossRef] [PubMed]
- Kubota, M.; Imai, H.T.; Kondo, M.; Onoyama, K.; Ogata, K.; Terayama, M.; Yoshimura, M. Japanese Ant Image Database. Available online: http://ant.miyakyo-u.ac.jp/ (accessed on 13 September 2017).
- Aili, S.R.; Touchard, A.; Escoubas, P.; Padula, M.P.; Orivel, J.; Dejean, A.; Nicholson, G.M. Diversity of peptide toxins from stinging ant venoms. Toxicon 2014, 92, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Aili, S.R.; Touchard, A.; Koh, J.M.; Dejean, A.; Orivel, J.; Padula, M.P.; Escoubas, P.; Nicholson, G.M. Comparisons of Protein and Peptide Complexity in Poneroid and Formicoid Ant Venoms. J. Proteome Res. 2016, 15, 3039–3054. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.A.; McCutcheon, J.P.; Nakabachi, A. Genomics and evolution of heritable bacterial symbionts. Annu. Rev. Genet. 2008, 42, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.W.; Xiong, S.J.; Xu, G.; Gan, S.Y.; Chen, X.; Stanley, D.; Yan, Z.C.; Ye, G.Y.; Fang, Q. Protein Discovery: Combined Transcriptomic and Proteomic Analyses of Venom from the Endoparasitoid Cotesia chilonis (Hymenoptera: Braconidae). Toxins (Basel) 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Shi, M.; Ye, X.Q.; Li, F.; Wang, X.W.; Chen, X.X. Comparative transcriptome analysis of venom glands from Cotesia vestalis and Diadromus collaris, two endoparasitoids of the host Plutella xylostella. Sci. Rep. 2017, 7, 1298. [Google Scholar] [CrossRef] [PubMed]
- Habermann, E.; Jentsch, J. [Sequence analysis of melittin from tryptic and peptic degradation products]. Hoppe Seyler's Z. Physiol. Chem. 1967, 348, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Shinada, T.; Ohfune, Y.; Hisada, M.; Yasuda, A.; Naoki, H.; Nakajima, T. Novel mastoparan and protonectin analogs isolated from a solitary wasp, Orancistrocerus drewseni drewseni. Amino Acids 2009, 37, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Donovan, G.R.; Baldo, B.A.; Sutherland, S. Molecular cloning and characterization of a major allergen (Myr p I) from the venom of the Australian jumper ant, Myrmecia pilosula. Biochim. Biophys. Acta 1993, 1171, 272–280. [Google Scholar] [CrossRef]
- Hsieh, S.K.; Tzen, J.T.; Wu, T.Y.; Chen, Y.J.; Yang, W.H.; Huang, C.F.; Hsieh, F.C.; Jinn, T.R. Functional expression and characterization of dipeptidyl peptidase IV from the black-bellied hornet Vespa basalis in Sf21 insect cells. Biosci. Biotechnol. Biochem. 2011, 75, 2371–2375. [Google Scholar] [CrossRef] [PubMed]
- Merkler, D.J. C-terminal amidated peptides: Production by the in vitro enzymatic amidation of glycine-extended peptides and the importance of the amide to bioactivity. Enzym. Microb. Technol. 1994, 16, 450–456. [Google Scholar] [CrossRef]
- Krishnan, A.; Nair, P.N.; Jones, D. Isolation, cloning, and characterization of new chitinase stored in active form in chitin-lined venom reservoir. J. Biol. Chem. 1994, 269, 20971–20976. [Google Scholar] [PubMed]
- Key, L.N.; Boyle, P.R.; Jaspars, M. Novel activities of saliva from the octopus Eledone cirrhosa (Mollusca; Cephalopoda). Toxicon 2002, 40, 677–683. [Google Scholar] [CrossRef]
- Tetreau, G.; Cao, X.; Chen, Y.R.; Muthukrishnan, S.; Jiang, H.; Blissard, G.W.; Kanost, M.R.; Wang, P. Overview of chitin metabolism enzymes in Manduca sexta: Identification, domain organization, phylogenetic analysis and gene expression. Insect Biochem. Mol. Biol. 2015, 62, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Peiren, N.; de Graaf, D.C.; Brunain, M.; Bridts, C.H.; Ebo, D.G.; Stevens, W.J.; Jacobs, F.J. Molecular cloning and expression of icarapin, a novel IgE-binding bee venom protein. FEBS Lett. 2006, 580, 4895–4899. [Google Scholar] [CrossRef] [PubMed]
- Bordon, K.C.; Wiezel, G.A.; Amorim, F.G.; Arantes, E.C. Arthropod venom Hyaluronidases: Biochemical properties and potential applications in medicine and biotechnology. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 43. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhu, S. Comparative genomics analysis of five families of antimicrobial peptide-like genes in seven ant species. Dev. Comp. Immunol. 2012, 38, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.O.; Blum, M.S.; Overal, W.L. Comparative enzymology of venoms from stinging Hymenoptera. Toxicon 1986, 24, 907–921. [Google Scholar] [CrossRef]
- Six, D.A.; Dennis, E.A. The expanding superfamily of phospholipase A2 enzymes: Classification and characterization. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2000, 1488, 1–19. [Google Scholar] [CrossRef]
- Dos Santos Pinto, J.R.; Fox, E.G.; Saidemberg, D.M.; Santos, L.D.; da Silva Menegasso, A.R.; Costa-Manso, E.; Machado, E.A.; Bueno, O.C.; Palma, M.S. Proteomic view of the venom from the fire ant Solenopsis invicta Buren. J. Proteome Res. 2012, 11, 4643–4653. [Google Scholar] [CrossRef] [PubMed]
- DDBJ. Available online: http://p.ddbj.nig.ac.jp/pipeline/ (accessed on 22 November 2016).
- UniProtKB. Available online: http://www.uniprot.org/ (accessed on 29 November 2016).
- MS-Product in ProteinProspector Program. Available online: http://prospector.ucsf.edu/prospector/cgi-bin/msform.cgi?form=msproduct (accessed on 1 September 2017).












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total number of reads | 44,087,058 |
| Average base pairs | 100 bp |
| Total number of contigs | 49,639 |
| (after cd-hit-est) | 43,662 |
| (-rRNA, -Bacterial transcripts, +Pilosulin-like peptide 1) | 41,764 |
| N50 contig size | 1466 bp |
| Fraction No. | Retention Time (min) | Amino Acid Sequence | Observed Molecular Mass a | Derivative |
|---|---|---|---|---|
| 9 | 8.15 | KVMDWLKDKM-NH2 | 1291.677 | PLP1 |
| 11 | 8.55 | VMDWLKDKM-NH2 | 1163.582 | PLP1 |
| 11 | 8.75 | AAANAAASEALSAMASQNE | 1776.795 | PLP3 |
| 12 | 9.08 | GLLDWGK | 787.423 | PLP1 |
| 12 | 9.21 | GWGSLFK | 793.412 | PLP2 |
| 12 | 9.40 | GWGSLFKT | 894.458 | PLP2 |
| 13 | 9.54 | GWGSLFKTVGKM | 1309.685 | PLP2 |
| 14 | 10.95 | GWGSLFKTVGKMLAKAAVK | 1991.142 | PLP2 |
| 14 | 11.08 | KTALEAAANAAASEALSAMASQNE | 2319.098 | PLP3 |
| 15 | 11.86 | GLLDWGKKVMDWLKDKm-NH2 | 2077.083 | PLP1 |
| 16 | 12.75 | GLLDWGKKVMDWLKDKM-NH2 | 2061.090 | PLP1 |
| Peak | ID | Retention Time (min) | Area (%) | Observed Molecular Mass a | Transcriptome Analysis Assisted Estimation of Mature Peptides | |||
|---|---|---|---|---|---|---|---|---|
| Possible Amino Acid Sequence | Molecular Formula | Theoretical Molecular Mass a | Derivative | |||||
| 4 | Om3264 | 10.58 | 0.41 | 3263.957 | ||||
| 5 | Om4117 | 11.04 | 0.25 | 4117.238 | ||||
| 6 | Om3489 | 13.84 | 0.46 | 3488.881 | ||||
| 7b | Om4229 | 14.45 | <0.1 | 4229.334 | KIKWGKIFKKGGKLIGKTALEAAANAAASEAISAMASQNEK | C188H317N53O55S | 4229.336 | PLP3 |
| 7a | Om3101 | 14.45 | <0.1 | 3100.615 | ||||
| 8 | Om4101 | 15.04 | 12.94 | 4101.244 | KIKWGKIFKKGGKLIGKTALEAAANAAASEAISAMASQNE | C182H305N51O54S | 4101.241 | PLP3 |
| 9 | Om3704 | 16.51 | 4.29 | 3704.235 | IKGKKIMKNMGKAMKIAGKVAKAMAPIVVPLIVSAA-NH2 | C168H306N46O38S4 | 3704.231 | PLP6 |
| 10 | Om4401 | 17.51 | 0.26 | 4401.364 | ||||
| 13 | Om6368 | 23.07 | 20.85 | 6367.557 | ||||
| 14 | Om6350 | 23.99 | 5.27 | 6349.589 | ||||
| 15 | Om6332 | 24.62 | 5.42 | 6331.631 | GVKELFGKAWGLVKKHLPKACGLLGYVKQ | C300H484N78O68S2 | 6331.625 | PLP4 |
| | | ||||||||
| GVKELFGKAWGLVKKHLPKACGLLGYVKQ | ||||||||
| 16 | Om3362 | 25.37 | 7.78 | 3361.756 | GWGSIFKTVGKMIAKAAVKAAPEAISAMASQNE | C149H224N40O44S2 | 3361.753 | PLP2 |
| 17 | Om3325 | 26.94 | 6.97 | 3324.856 | ||||
| 18 | Om2247 | 27.66 | 1.52 | 2247.188 | GILDWGKKVMDWIKDKMGK | C103H166N26O26S2 | 2247.191 | PLP1 |
| 19b | Om2119 | 29.26 | <1.0 | 2119.094 | GILDWGKKVMDWIKDKMG | C97H154N24O25S2 | 2119.096 | PLP1 |
| 19a | Om2061 | 29.76 | 27.41 | 2061.089 | GILDWGKKVMDWIKDKM-NH2 | C95H152N24O23S2 | 2061.090 | PLP1 |
| 20 | Om1782 | 32.05 | 1.08 | 1782.019 | ||||
| 21 | Om1812 | 33.62 | 0.67 | 1812.030 | ||||
| 22 | Om1837 | 34.86 | 3.65 | 1836.995 | IWGALLGTLIPAITSAIQ-NH2 | C87H144N20O23 | 1837.071 | PLP5 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazuma, K.; Masuko, K.; Konno, K.; Inagaki, H. Combined Venom Gland Transcriptomic and Venom Peptidomic Analysis of the Predatory Ant Odontomachus monticola. Toxins 2017, 9, 323. https://doi.org/10.3390/toxins9100323
Kazuma K, Masuko K, Konno K, Inagaki H. Combined Venom Gland Transcriptomic and Venom Peptidomic Analysis of the Predatory Ant Odontomachus monticola. Toxins. 2017; 9(10):323. https://doi.org/10.3390/toxins9100323
Chicago/Turabian StyleKazuma, Kohei, Keiichi Masuko, Katsuhiro Konno, and Hidetoshi Inagaki. 2017. "Combined Venom Gland Transcriptomic and Venom Peptidomic Analysis of the Predatory Ant Odontomachus monticola" Toxins 9, no. 10: 323. https://doi.org/10.3390/toxins9100323
APA StyleKazuma, K., Masuko, K., Konno, K., & Inagaki, H. (2017). Combined Venom Gland Transcriptomic and Venom Peptidomic Analysis of the Predatory Ant Odontomachus monticola. Toxins, 9(10), 323. https://doi.org/10.3390/toxins9100323