
Shiga Toxin Glycosphingolipid Receptors in Human Caco-2 and HCT-8 Colon Epithelial Cell Lines

Abstract
:1. Introduction
2. Results
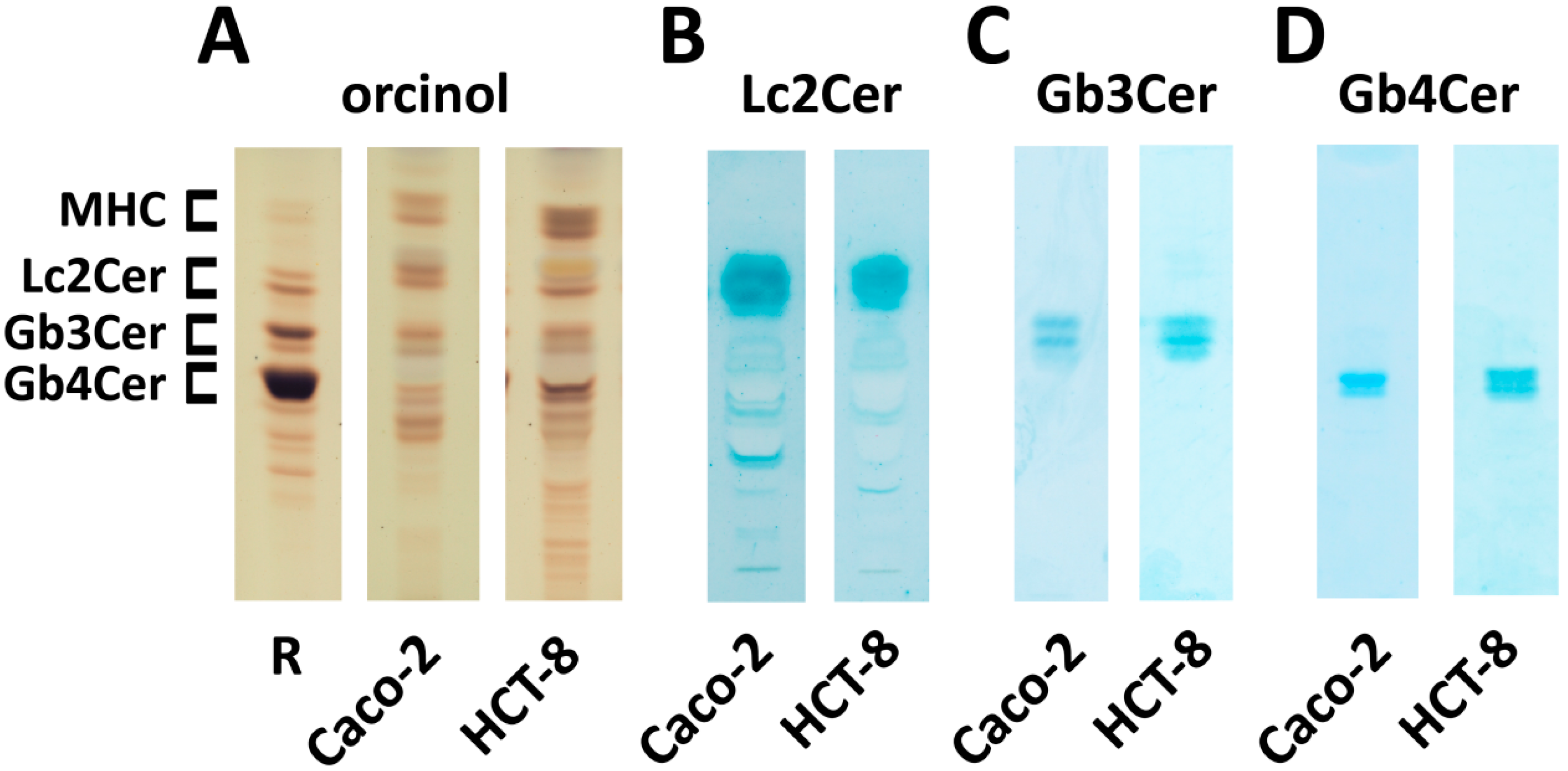
2.1. Identification of Neutral GSLs of the Globo-Series in Caco-2 and HCT-8 Cells
2.2. Structural Characterization of Sphingolipids from the Neutral GSL Fraction of Caco-2 and HCT-8 Cells
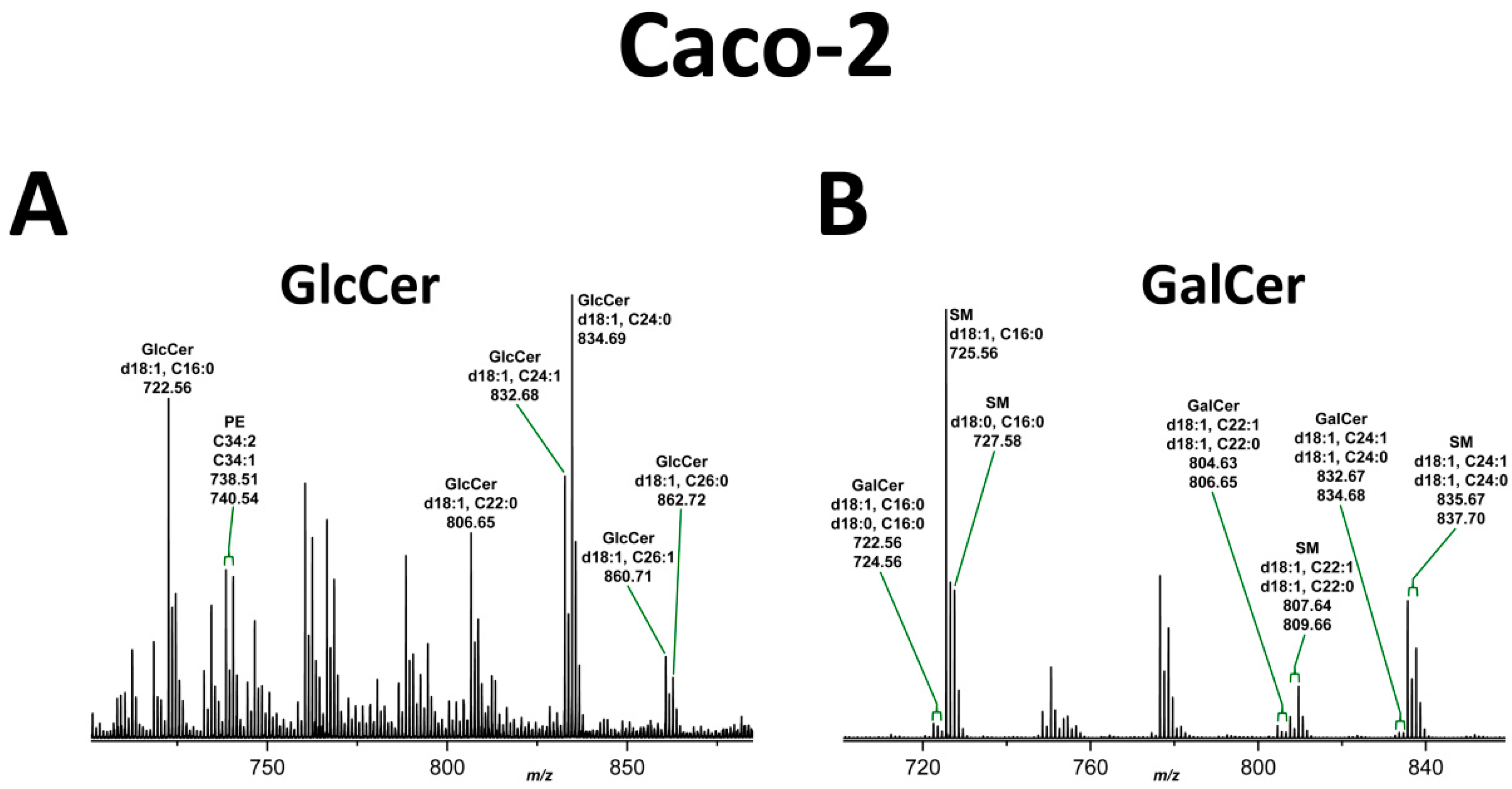
2.3. Structural Characterization of GlcCer and GalCer Lipoforms Detected in the Neutral GSL Fraction of Caco-2 and HCT-8 Cells
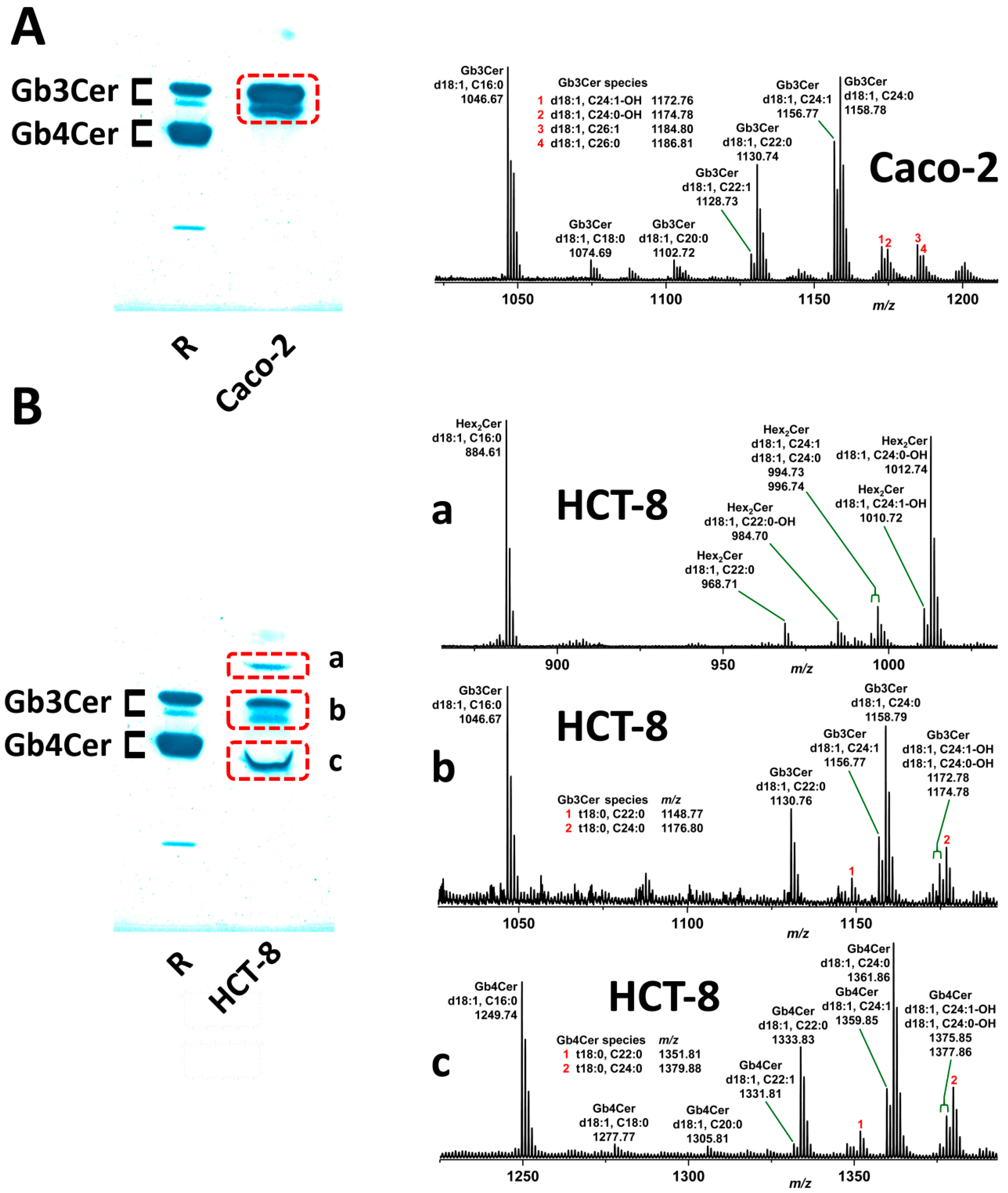
2.4. Relative Content of the Various Gb3Cer and Gb4Cer Lipoforms of Caco-2 and HCT-8 Cells
2.5. Identification and Structural Characterization of Stx2a-Binding GSLs
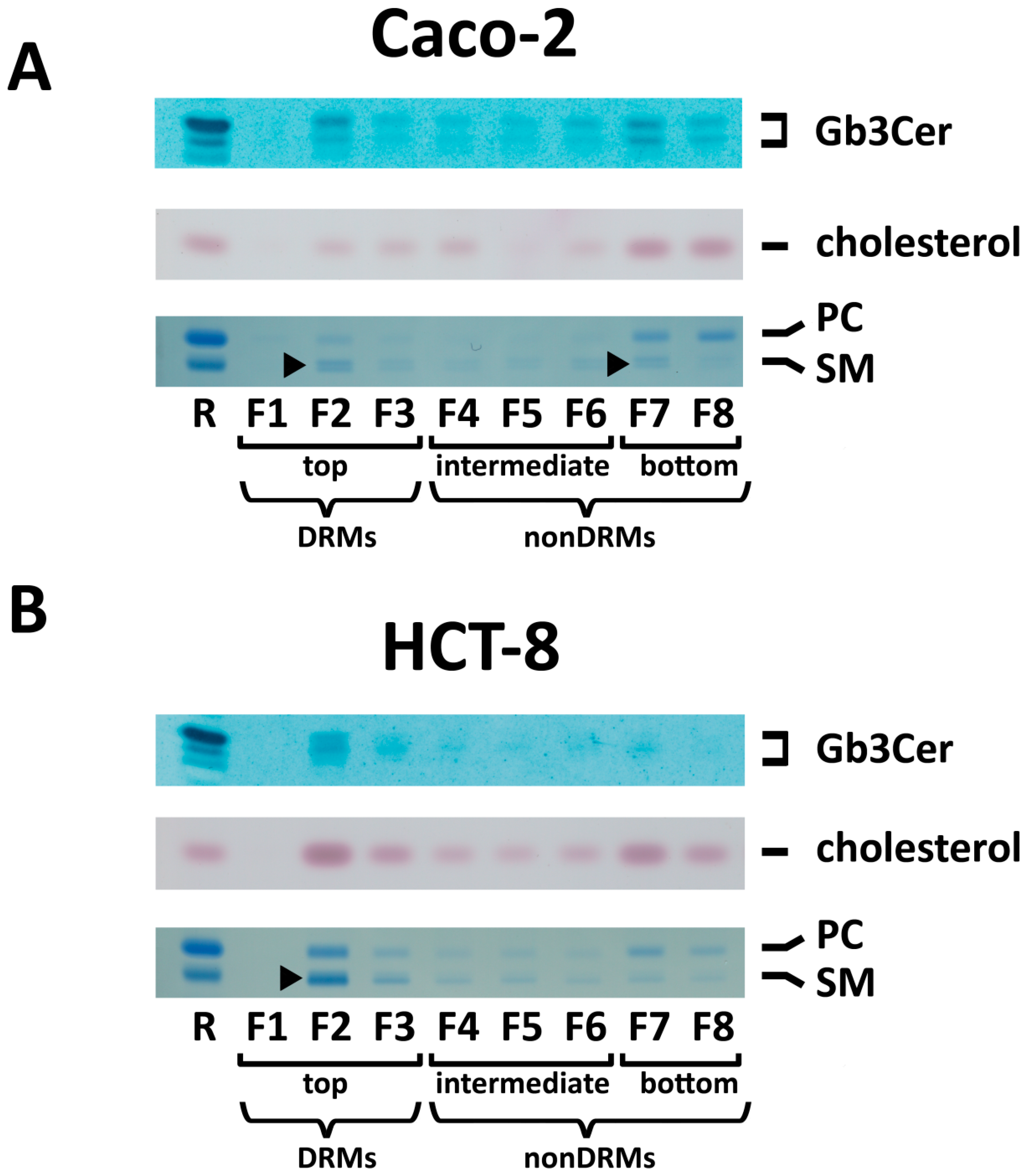
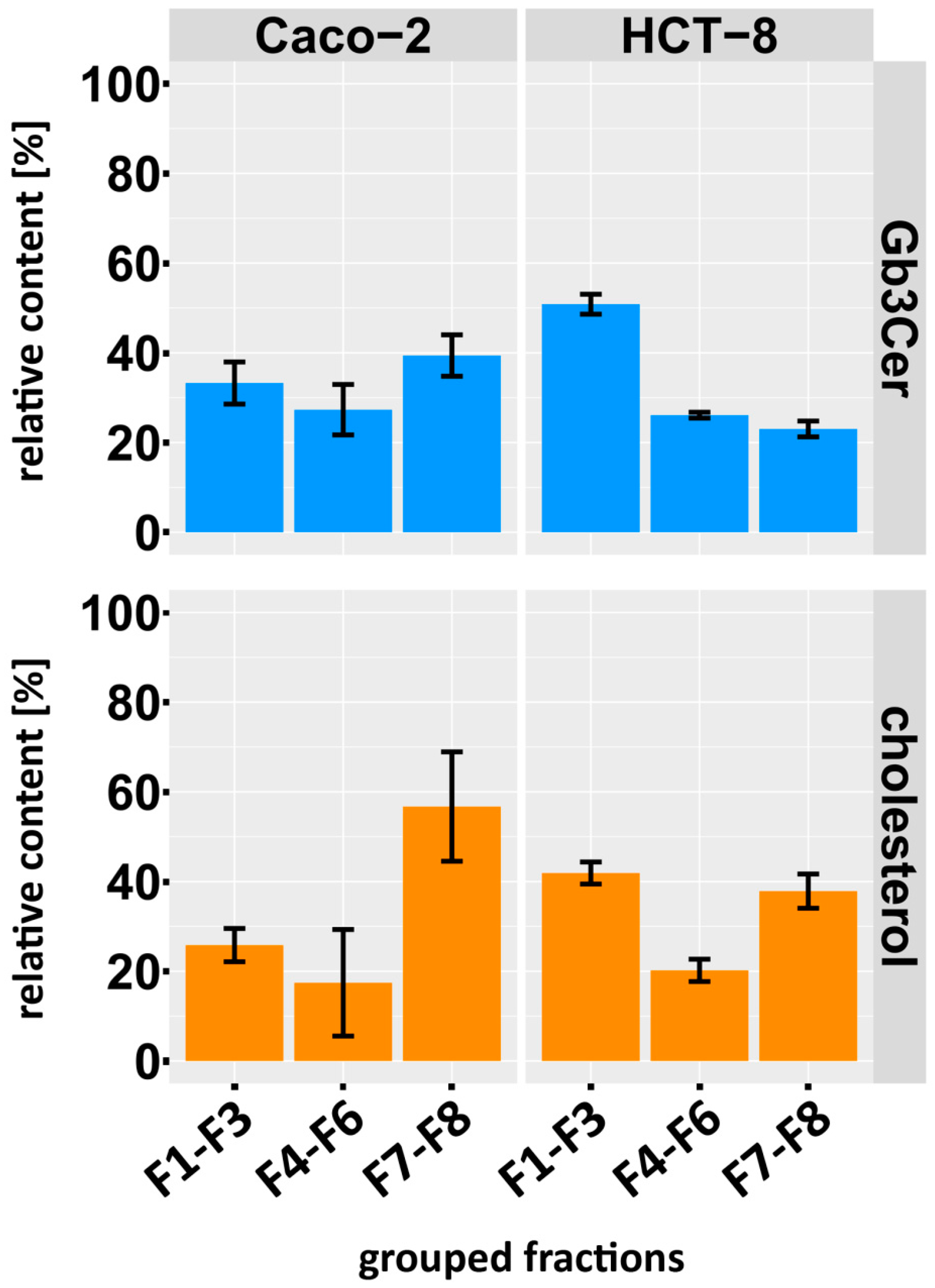
2.6. Distribution of Gb3Cer and Gb4Cer to DRM and NonDRM Fractions
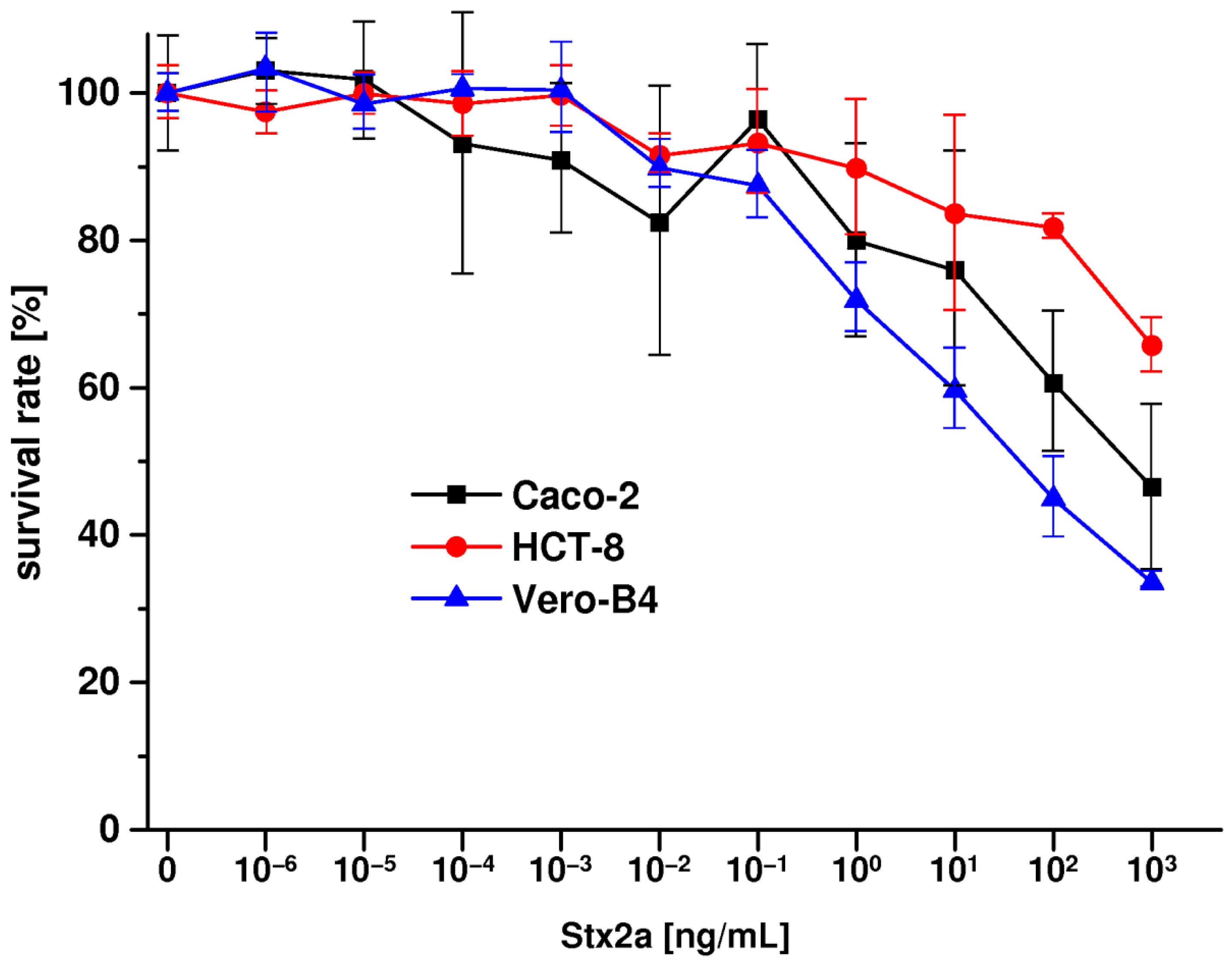
2.7 Stx2a-Mediated Cellular Damage of Caco-2 and HCT-8 Cells
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cultivation of Caco-2, HCT-8 and Vero-B4 Cell Lines and Stx2a Cytotoxicity Assay
5.2. Isolation of Lipids and Purification of GSLs from Cultured Cell Lines
5.3. Preparation of Sucrose Density Gradient Fractions by Ultracentrifugation
5.4. Isolation of Lipids from Sucrose Density Gradient Fractions
5.5. Lipid References, Antibodies and Stx2a
5.6. Thin-Layer Chromatography and Lipid Staining
5.7. TLC Overlay Assay and Lipid Semiquantification
5.8. Mass Spectrometry of GSLs
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Karch, H.; Tarr, P.I.; Bielaszewska, M. Enterohaemorrhagic Escherichia coli in human medicine. Int. J. Med. Microbiol. 2005, 295, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Tarr, P.I.; Gordon, C.A.; Chandler, W.L. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 2005, 365, 1073–1086. [Google Scholar] [CrossRef]
- Obrig, T.G. Escherichia coli Shiga toxin mechanisms of action in renal disease. Toxins (Basel) 2010, 2, 2769–2794. [Google Scholar] [CrossRef] [PubMed]
- Melton-Celsa, A.R. Shiga toxin (Stx) classification, structure, and function. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Koo, S.; Tesh, V.L. Shiga toxins as multi-functional proteins: Induction of host cellular stress responses, role in pathogenesis and therapeutic applications. Toxins (Basel) 2016, 8, 77. [Google Scholar] [CrossRef] [PubMed]
- Karpman, D.; Loos, S.; Tati, R.; Arvidsson, I. Haemolytic uraemic syndrome. J. Intern. Med. 2017, 281, 123–148. [Google Scholar] [CrossRef] [PubMed]
- Bergan, J.; Dyve Lingelem, A.B.; Simm, R.; Skotland, T.; Sandvig, K. Shiga toxins. Toxicon 2012, 60, 1085–1107. [Google Scholar] [CrossRef] [PubMed]
- Lainhart, W.; Stolfa, G.; Koudelka, G.B. Shiga toxin as a bacterial defense against eukaryotic predator, Tetrahymena thermophila. J. Bacteriol. 2009, 191, 5116–5122. [Google Scholar] [CrossRef] [PubMed]
- Mauro, S.A.; Koudelka, G.B. Shiga toxin: Expression, distribution, and its role in the environment. Toxins (Basel) 2011, 3, 608–625. [Google Scholar] [CrossRef] [PubMed]
- Arnold, J.W.; Koudelka, G.B. The Trojan Horse of the microbial arms race: Phage-encoded toxins as a defence against eukaryotic predators. Environ. Microbiol. 2014, 16, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Nakao, H.; Takeda, T. Escherichia coli Shiga toxin. J. Nat. Toxins 2000, 9, 299–313. [Google Scholar] [PubMed]
- Karmali, M.A. Prospects for preventing serious systemic toxemic complications of Shiga toxin producing Escherichia coli infections using Shiga toxin receptor analogues. J. Infect. Dis. 2004, 189, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Karch, H.; Friedrich, A.W.; Gerber, A.; Zimmerhackl, L.B.; Schmidt, M.A.; Bielaszewska, M. New aspects in the pathogenesis of enteropathic hemolytic uremic syndrome. Semin. Thromb. Hemost. 2006, 32, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Te Loo, D.M.; Monnens, L.A.; van der Velden, T.J.; Vermeer, M.A.; Preyers, F.; Demacker, P.N.; van den Heuvel, L.P.; van Hinsbergh, V.W. Binding and transfer of verocytotoxin by polymorphonuclear leukocytes in hemolytic uremic syndrome. Blood 2000, 95, 3396–3402. [Google Scholar] [PubMed]
- Brigotti, M.; Carnicelli, D.; Ravanelli, E.; Barbieri, S.; Ricci, F.; Bontadini, A.; Tozzi, A.E.; Scavia, G.; Caprioloi, A.; Tazzari, P.L. Interactions between Shiga toxins and human polymorphonuclear leukocytes. J. Leukoc. Biol. 2008, 84, 1019–1027. [Google Scholar] [CrossRef] [PubMed]
- Brigotti, M.; Carnicelli, D.; Arfilli, V.; Tamassia, N.; Borsetti, F.; Fabbri, E.; Tazzari, P.L.; Ricci, F.; Pagliaro, P.; Spisni, E.; et al. Identification of TLR4 as the receptor that recognizes Shiga toxin in human neutrophils. J. Immunol. 2013, 191, 4748–4758. [Google Scholar] [CrossRef] [PubMed]
- Arfilli, V.; Carnicelli, D.; Ardissino, G.; Torresani, E.; Scavia, G.; Brigotti, M. A rapid and sensitive method to measure the functional activity of Shiga toxins in human serum. Toxins (Basel) 2015, 7, 4564–4576. [Google Scholar] [CrossRef] [PubMed]
- Yagi, H.; Narita, N.; Matsumoto, M.; Sakurai, Y.; Ikari, H.; Yoshioka, A.; Kita, E.; Ikeda, Y.; Titani, K.; Fujimura, Y. Enhanced low shear stress induced platelet aggregation by Shiga-like toxin 1 purified from Escherichia coli O157. Am. J. Hematol. 2001, 66, 105–115. [Google Scholar] [CrossRef]
- Ghosh, S.A.; Polanowska-Grabowska, R.K.; Fujii, J.; Obrig, T.; Gear, A.R. Shiga toxin binds to activated platelets. J. Thromb. Haemost. 2004, 2, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Geelen, J.M.; van der Velden, T.J.; van den Heuvel, L.P.; Monnens, L.A. Interactions of Shiga-like toxin with human peripheral blood monocytes. Pediatr. Nephrol. 2007, 22, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Lee, M.S.; Cherla, R.P.; Tesh, V.L. Shiga toxin 1 induces apoptosis through the endoplasmic reticulum stress response in human monocytic cells. Cell. Microbiol. 2008, 10, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Arfilli, V.; Carnicelli, D.; Rocchi, L.; Ricci, F.; Pagliaro, P.; Tazzari, P.L.; Brigotti, M. Shiga toxin 1 and ricin A chain bind to human polymorphonuclear leucocytes through a common receptor. Biochem. J. 2010, 432, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Brigotti, M.; Tazzari, P.L.; Ravanelli, E.; Carnicelli, D.; Barbieri, S.; Rocchi, L.; Arfilli, V.; Scavia, G.; Ricci, F.; Bontadini, A.; et al. Endothelial damage induced by Shiga toxins delivered by neutrophils during transmigration. J. Leukoc. Biol. 2010, 88, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Schweppe, C.H.; Hoffmann, P.; Nofer, J.R.; Pohlentz, G.; Mormann, M.; Karch, H.; Friedrich, A.W.; Müthing, J. Neutral glycosphingolipids in human blood: A precise mass spectrometry analysis with special reference to lipoprotein-associated Shiga toxin receptors. J. Lipid Res. 2010, 51, 2282–2294. [Google Scholar] [CrossRef] [PubMed]
- Kouzel, I.U.; Pohlentz, G.; Storck, W.; Radamm, L.; Hoffmann, P.; Bielaszewska, M.; Bauwens, A.; Cichon, C.; Schmidt, M.A.; Mormann, M.; et al. Association of Shiga toxin glycosphingolipid receptors with membrane microdomains of toxin-sensitive lymphoid and myeloid cells. J. Lipid Res. 2013, 51, 692–710. [Google Scholar] [CrossRef] [PubMed]
- Ståhl, A.L.; Arvidsson, I.; Johansson, K.E.; Chromek, M.; Rebetz, J.; Loos, S.; Kristoffersson, A.C.; Békássy, Z.D.; Mörgelin, M.; Karpman, D. A novel mechanism of bacterial transfer within host blood cell-derived microvesicles. PLoS Pathog. 2015, 11, e1004619. [Google Scholar] [CrossRef] [PubMed]
- Karpman, D.; Ståhl, A.L.; Arvidsson, I. Extracellular vesicles in renal disease. Nat. Rev. Nephrol. 2017, 13, 545–562. [Google Scholar] [CrossRef] [PubMed]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Piérard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.S.; Ng, T.B. Shiga toxins: From structure and mechanism to applications. Appl. Microbiol. Biotechnol. 2016, 100, 1597–1610. [Google Scholar] [CrossRef] [PubMed]
- Müthing, J.; Meisen, I.; Zhang, W.; Bielaszewska, M.; Mormann, M.; Bauerfeind, R.; Schmidt, M.A.; Friedrich, A.W.; Karch, H. Promiscuous Shiga toxin 2e and its intimate relationship to Forssman. Glycobiology 2012, 22, 849–862. [Google Scholar] [CrossRef] [PubMed]
- DeGrandis, S.; Law, H.; Brunton, J.; Gyles, C.; Lingwood, C.A. Globotetraosylceramide is recognized by the pig edema disease toxin. J. Biol. Chem. 1989, 264, 12502–12505. [Google Scholar]
- Steil, D.; Schepers, C.L.; Pohlentz, G.; Legros, N.; Runde, J.; Humpf, H.U.; Karch, H.; Müthing, J. Shiga toxin glycosphingolipid receptors of Vero-B4 kidney epithelial cells and their membrane microdomain lipid environment. J. Lipid Res. 2015, 56, 2322–2336. [Google Scholar] [CrossRef] [PubMed]
- Römer, W.; Berland, L.; Chambon, V.; Gaus, K.; Windschiegl, B.; Tenza, D.; Aly, M.R.; Fraisier, V.; Florent, J.C.; Perrais, D.; et al. Shiga toxin induces tubular membrane invaginations for its uptake into cells. Nature 2007, 450, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, K.; Bergan, J.; Dyve, A.B.; Skotland, T.; Torgersen, M.L. Endocytosis and retrograde transport of Shiga toxin. Toxicon 2010, 56, 1181–1185. [Google Scholar] [CrossRef] [PubMed]
- Johannes, L.; Römer, W. Shiga toxins—From cell biology to biomedical applications. Nat. Rev. Microbiol. 2010, 8, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Römer, W.; Pontani, L.L.; Sorre, B.; Rentero, C.; Berland, L.; Chambon, V.; Lamaze, C.; Bassereau, P.; Sykes, C.; Gaus, K.; et al. Actin dynamics drive membrane reorganization and scission in clathrin-independent endocytosis. Cell 2010, 140, 540–553. [Google Scholar] [CrossRef] [PubMed]
- Kavaliauskiene, S.; Dyve Lingelem, A.B.; Skotland, T.; Sandvig, K. Protection against Shiga toxins. Toxins (Basel) 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Brigotti, M.; Alfieri, R.; Sestili, P.; Bonelli, M.; Petronini, P.G.; Guidarelli, A.; Barbieri, L.; Stirpe, F.; Sperti, S. Damage to nuclear DNA induced by Shiga toxin 1 and ricin in human endothelial cells. FASEB J. 2002, 16, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Sestili, P.; Alfieri, R.; Carnicelli, D.; Martinelli, C.; Barbieri, L.; Stirpe, F.; Bonelli, M.; Petronini, P.G.; Brigotti, M. Shiga toxin 1 and ricin inhibit the repair of H2O2-induced DNA single strand breaks in cultured mammalian cells. DNA Repair 2005, 4, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Brigotti, M.; Arfilli, V.; Carnicelli, D.; Rocchi, L.; Calcabrini, C.; Ricci, F.; Pagliaro, P.; Tazzari, P.L.; Alfieri, R.R.; Petronini, P.G.; et al. Shiga toxin 1, as DNA repair inhibitor, synergistically potentiates the activity of the anticancer drug, mafosfamide, on Raji cells. Toxins (Basel) 2013, 5, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Tesh, V.L. Activation of cell stress response pathways by Shiga toxins. Cell. Microbiol. 2012, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Karch, H. Consequences of enterohaemorrhagic Escherichia coli infection for the vascular endothelium. Thromb. Haemost. 2005, 94, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Müthing, J.; Schweppe, C.H.; Karch, H.; Friedrich, A.W. Shiga toxins, glycosphingolipid diversity, and endothelial cell injury. Thromb. Haemost. 2009, 101, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Zoja, C.; Buelli, S.; Morigi, M. Shiga toxin-associated hemolytic uremic syndrome: Pathophysiology of endothelial dysfunction. Pediatr. Nephrol. 2010, 25, 2231–2240. [Google Scholar] [CrossRef] [PubMed]
- Bauwens, A.; Betz, J.; Meisen, I.; Kemper, B.; Karch, H.; Müthing, J. Facing glycosphingolipid-Shiga toxin interaction: Dire straits for endothelial cells of the human vasculature. Cell. Mol. Life Sci. 2013, 70, 425–457. [Google Scholar] [CrossRef] [PubMed]
- Obrig, T.G.; Louise, C.B.; Lingwood, C.A.; Boyd, B.; Barley-Malony, L.; Daniel, T.O. Endothelial heterogeneity in Shiga toxin receptors and responses. J. Biol. Chem. 1993, 268, 15484–15488. [Google Scholar] [PubMed]
- Van Setten, P.A.; van Hinsbergh, V.W.; van der Velden, T.J.; van de Kar, N.C.; Vermeer, M.; Mahan, J.D.; Assmann, K.J.; van den Heuvel, L.P.; Monnens, L.A. Effects of TNFα on verocytotoxin cytotoxicity in purified human glomerular microvascular endothelial cells. Kidney Int. 1997, 51, 1245–1256. [Google Scholar] [CrossRef] [PubMed]
- Müthing, J.; Duvar, S.; Heitmann, D.; Hanisch, F.G.; Neumann, U.; Lochnit, G.; Geyer, R.; Peter-Katalinić, J. Isolation and structural characterization of glycosphingolipids of in vitro propagated human umbilical vein endothelial cells. Glycobiology 1999, 9, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Eisenhauer, P.B.; Chaturvedi, P.; Fine, R.E.; Ritchie, A.J.; Pober, J.S.; Cleary, T.G.; Newburg, D.S. Tumor necrosis factor alpha increases human cerebral endothelial cell Gb3 and sensitivity to Shiga toxin. Infect. Immun. 2001, 69, 1889–1894. [Google Scholar] [CrossRef] [PubMed]
- Stricklett, P.K.; Hughes, A.K.; Ergonul, Z.; Kohan, D.E. Molecular basis for up-regulation by inflammatory cytokines of Shiga toxin 1 cytotoxicity and globotriaosylceramide expression. J. Infect. Dis. 2002, 186, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Obrig, T.G.; Seaner, R.M.; Bentz, M.; Lingwood, C.A.; Boyd, B.; Smith, A.; Narrow, W. Induction by sphingomyelinase of Shiga toxin receptor and Shiga toxin 2 sensitivity in human microvascular endothelial cells. Infect. Immun. 2003, 71, 845–849. [Google Scholar] [CrossRef] [PubMed]
- Schweppe, C.H.; Bielaszewska, M.; Pohlentz, G.; Friedrich, A.W.; Büntemeyer, H.; Schmidt, M.A.; Kim, K.S.; Peter-Katalinić, J.; Karch, H.; Müthing, J. Glycosphingolipids in vascular endothelial cells: Relationship of heterogeneity in Gb3Cer/CD77 receptor expression with differential Shiga toxin 1 cytotoxicity. Glycoconj. J. 2008, 25, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Nakakita, S.I.; Nakayama, K.I. Structural characterization and dynamics of globotetraosylceramide in vascular endothelial cells under TNF-α stimulation. Glycoconj. J. 2010, 27, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Betz, J.; Bielaszewska, M.; Thies, A.; Humpf, H.U.; Dreisewerd, K.; Karch, H.; Kim, K.S.; Friedrich, A.W.; Müthing, J. Shiga toxin glycosphingolipid receptors in microvascular and macrovascular endothelial cells: Differential association with membrane lipid raft microdomains. J. Lipid Res. 2011, 52, 618–634. [Google Scholar] [CrossRef] [PubMed]
- Betz, J.; Bauwens, A.; Kunsmann, L.; Bielaszewska, M.; Mormann, M.; Humpf, H.U.; Karch, H.; Friedrich, A.W.; Müthing, J. Uncommon membrane distribution of Shiga toxin glycosphingolipid receptors in toxin-sensitivie human glomerular microvascular endothelial cells. Biol. Chem. 2012, 393, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Legros, N.; Dusny, S.; Humpf, H.U.; Pohlentz, G.; Karch, H.; Müthing, J. Shiga toxin glycosphingolipid receptors and their lipid membrane ensemble in primary human blood-brain-barrier endothelial cells. Glycobiology 2017, 27, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Legros, N.; Pohlentz, G.; Runde, J.; Dusny, S.; Humpf, H.U.; Karch, H.; Müthing, J. Colocalization of receptors for Shiga toxins with lipid rafts in primary human renal glomerular endothelial cells and influence of D-PDMP on synthesis and distribution of glycosphingolipid receptors. Glycobiology 2017, 27, 947–965. [Google Scholar] [CrossRef] [PubMed]
- Schüller, S. Shiga toxin interaction with human intestinal epithelium. Toxins (Basel) 2011, 3, 626–639. [Google Scholar] [CrossRef] [PubMed]
- Björk, S.; Breimer, M.E.; Hansson, G.C.; Karlsson, K.A.; Leffler, H. Structures of blood group glycosphingolipids of human small intestine. A relation between the expression of fucolipids of epithelial cells and the ABO, Le and Se phenotype of the donor. J. Biol. Chem. 1987, 262, 6758–6765. [Google Scholar] [PubMed]
- Holgersson, J.; Jovall, P.Å.; Breimer, M.E. Glycosphingolipids of human large intestine: Detailed structural characterization with special reference to blood group compounds and bacterial receptor structures. J. Biochem. 1991, 110, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Schüller, S.; Frankel, G.; Phillips, A.D. Interaction of Shiga toxin from Escherichia coli with human intestinal epithelial cell lines and explants: Stx2 induces epithelial damage in organ culture. Cell. Microbiol. 2004, 6, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Zumbrun, S.D.; Hanson, L.; Sinclair, J.F.; Freedy, J.; Melton-Celsa, A.R.; Rodriguez-Canales, J.; Hanson, J.C.; O’Brien, A.D. Human intestinal tissue and cultured colonic cells contain globotriaosylceramide synthase mRNA and the alternate Shiga toxin receptor globotetraosylceramide. Infect. Immun. 2010, 78, 4488–4499. [Google Scholar] [CrossRef] [PubMed]
- Kovbasnjuk, O.; Mourtazina, R.; Baibakov, B.; Wang, T.; Elowsky, C.; Choti, M.A.; Kane, A.; Donowitz, M. The glycosphingolipid globotriaosylceramide in the metastatic transformation of colon cancer. Proc. Natl. Acad. Sci. USA 2005, 102, 19087–19092. [Google Scholar] [CrossRef] [PubMed]
- Falguières, T.; Maak, M.; von Weyhern, C.; Sarr, M.; Sastre, X.; Poupon, M.F.; Robine, S.; Johannes, L.; Janssen, K.P. Human colorectal tumors and metastases express Gb3 and can be targeted by an intestinal pathogen-based delivery tool. Mol. Cancer Ther. 2008, 7, 2498–2508. [Google Scholar] [CrossRef] [PubMed]
- Distler, U.; Souady, J.; Hülsewig, M.; Drmić-Hofman, I.; Haier, J.; Friedrich, A.W.; Karch, H.; Senninger, N.; Dreisewerd, K.; Berkenkamp, S.; et al. Shiga toxin receptor Gb3Cer/CD77: Tumor association and promising therapeutic target in pancreas and colon cancer. PLoS ONE 2009, 4, e6813. [Google Scholar] [CrossRef] [PubMed]
- Engedal, N.; Skotland, T.; Torgersen, M.L.; Sandvig, K. Shiga toxin and its use in targeted cancer therapy and imaging. Microb. Biotechnol. 2011, 4, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Behnam-Motlagh, P.; Tyler, A.; Grankvist, K.; Johansson, A. Verotoxin-1 treatment or manipulation of its receptor globotriaosylceramide (gb3) for reversal of multidrug resistance to cancer chemotherapy. Toxins (Basel) 2010, 2, 2467–2477. [Google Scholar] [CrossRef] [PubMed]
- Maak, M.; Nitsche, U.; Keller, L.; Wolf, P.; Sarr, M.; Thiebaud, M.; Rosenberg, R.; Langer, R.; Kleeft, J.; Friess, H.; et al. Tumor-specific targeting of pancreatic cancer with Shiga toxin B-subunit. Mol. Cancer Ther. 2011, 10, 1918–1928. [Google Scholar] [CrossRef] [PubMed]
- Storck, W.; Meisen, I.; Gianmoena, K.; Pläger, I.; Kouzel, I.U.; Bielaszewska, M.; Haier, J.; Mormann, M.; Humpf, H.U.; Karch, H.; et al. Shiga toxin glycosphingolipid receptor expression and toxin susceptibility of human pancreatic ductal adenocarcinomas of differing origin and differentiation. Biol. Chem. 2012, 393, 785–799. [Google Scholar] [CrossRef] [PubMed]
- Stimmer, L.; Dehay, S.; Nemati, F.; Massonnet, G.; Richon, S.; Decaudin, D.; Klijanienko, J.; Johannes, L. Human breast cancer and lymph node metastases express Gb3 and can be targeted by StxB-vectorized chemotherapeutic compounds. BMC Cancer 2014, 14, 916. [Google Scholar] [CrossRef] [PubMed]
- Geyer, P.E.; Maak, M.; Nitsche, U.; Perl, M.; Novotny, A.; Slotta-Huspenina, J.; Dransart, E.; Holtorf, A.; Johannes, L.; Janssen, K.P. Gastric adenocarcinomas express the glycosphingolipid Gb3/CD77: Targeting of gastric cancer cells with Shiga toxin B-subunit. Mol. Cancer Ther. 2016, 15, 1008–1017. [Google Scholar] [CrossRef] [PubMed]
- Jacewicz, M.S.; Acheson, D.W.K.; Mobassaleh, M.; Donohue-Rolfe, A.; Balasubramanian, K.A.; Keusch, G.T. Maturational regulation of globotriaosylceramide, the Shiga-like toxin 1 receptor, in cultured human gut epithelial cells. J. Clin. Investig. 1995, 96, 1328–1335. [Google Scholar] [CrossRef] [PubMed]
- Miller-Podraza, H. Polyglycosylceramides, poly-N-acetyllactosamine-containing glycosphingolipids: Methods of analysis, structure, and presumable biological functions. Chem. Rev. 2000, 100, 4663–4682. [Google Scholar] [CrossRef] [PubMed]
- Levery, S.B. Glycosphingolipid structural analysis and glycosphingolipidomics. Methods Enzymol. 2005, 405, 300–369. [Google Scholar] [PubMed]
- Müthing, J.; Distler, U. Advances on the compositional analysis of glycosphingolipids combining thin-layer chromatography with mass spectrometry. Mass Spectrom. Rev. 2010, 29, 425–479. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, G.; Capasso, S.; Sticco, L.; Russo, D. Glycosphingolipids: Synthesis and functions. FEBS J. 2013, 280, 6338–6353. [Google Scholar] [CrossRef] [PubMed]
- Ilver, D.; Johansson, P.; Miller-Podraza, H.; Nyholm, P.G.; Teneberg, S.; Karlsson, K.A. Bacterium-host protein-carbohydrate interactions. Methods Enzymol. 2003, 363, 134–157. [Google Scholar] [PubMed]
- Stroud, M.R.; Stapleton, A.E.; Levery, S.B. The P histo-blood group-related glycosphingolipid sialosyl galacrosyl globoside as a preferred binding receptor for uropathogenic Escherichia coli: Isolation and structural characterization from human kidney. Biochemistry 1998, 37, 17420–17428. [Google Scholar] [CrossRef] [PubMed]
- Dodson, K.W.; Pinkner, J.S.; Rose, T.; Magnusson, G.; Hultgren, S.J.; Waksman, G. Structural basis of the interaction of the pyelonephritic E. coli adhesin to its human kidney receptor. Cell 2001, 105, 733–743. [Google Scholar] [CrossRef]
- Johansson, P.; Nilsson, J.; Ångström, J.; Miller-Podraza, H. Interaction of Helicobacter pylori with sialylated carbohydrates: The dependence on different parts of the binding trisaccharide Neu5Acα3Galβ4GlcNAc. Glycobiology 2005, 15, 625–636. [Google Scholar] [CrossRef] [PubMed]
- Miller-Podraza, H.; Weikkolainen, K.; Larsson, T.; Linde, P.; Helin, J.; Natunen, J.; Karlsson, K.A. Helicobacter pylori binding to new glycans based on N-acetyllactosamine. Glycobiology 2009, 19, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, K.A. Animal glycosphingolipids as membrane attachment sites for bacteria. Annu. Rev. Biochem. 1989, 58, 309–350. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.K.; van de Kar, N.C.; Tarr, P.I. Shiga toxin/verocytotoxin-producing Escherichia coli infections: Practical clinical perspectives. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Fratamico, P.M.; Gunther, N.W., 4th. Shiga toxin-producing Escherichia coli. Adv. Appl. Microbiol. 2014, 86, 145–197. [Google Scholar] [PubMed]
- Bryan, A.; Youngster, I.; McAdam, A.J. Shiga toxin producing Escherichia coli. Clin. Lab. Med. 2015, 35, 247–272. [Google Scholar] [CrossRef] [PubMed]
- Badizadegan, K.; Wolf, A.A.; Rodighiero, C.; Jobling, M.; Hirst, T.R.; Holmes, R.K.; Lencer, W.I. Floating cholera toxin into epithelial cells: Functional association with caveolae-like detergent-insoluble membrane microdomains. Int. J. Med. Microbiol. 2000, 290, 403–408. [Google Scholar] [CrossRef]
- Binnington, B.; Lingwood, D.; Nutikka, A.; Lingwood, C.A. Effect of globotriaosyl ceramide fatty acid α-hydroxylation on the binding by verotoxin 1 and verotoxin 2. Neurochem. Res. 2002, 27, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Mahfoud, R.; Manis, A.; Lingwood, C.A. Fatty acid-dependent globotriaosyl ceramide receptor function in detergent resistant model membranes. J. Lipid Res. 2009, 50, 1744–1755. [Google Scholar] [CrossRef] [PubMed]
- Lingwood, C.A.; Binnington, B.; Manis, A.; Branch, D.R. Globotriaosyl ceramide receptor function—Where membrane structure and pathology intersect. FEBS Lett. 2010, 584, 1879–1886. [Google Scholar] [CrossRef] [PubMed]
- Wernick, N.L.; Chinnapen, D.J.; Cho, J.A.; Lencer, W.I. Cholera toxin: An intracellular journey into the cytosol by way of the endoplasmic reticulum. Toxins (Basel) 2010, 2, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Ewers, H.; Helenius, A. Lipid-mediated endocytosis. Cold Spring Harb. Perspect. Biol. 2011, 3, a004721. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.A.; Chinnapen, D.J.; Aamar, E.; te Welscher, Y.M.; Lencer, W.I.; Massol, R. Insights on the trafficking and retro-translocation of glycosphingolipid-binding bacterial toxins. Front. Cell. Infect. Microbiol. 2012, 2, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, C.; Natori, Y.; Zeng, X.T.; Ohmura, M.; Yamasaki, S.; Takeda, Y.; Natori, Y. Induction of cytokines in a human colon epithelial cell line by Shiga toxin 1 (Stx1) and Stx2 but not by non-toxic mutant Stx1 which lacks N-glycosidase activity. FEBS Lett. 1999, 442, 231–234. [Google Scholar] [CrossRef]
- Jones, N.L.; Islur, A.; Haq, R.; Mascarenhas, M.; Karmali, M.A.; Perdue, M.H.; Zanke, B.W.; Sherman, P.M. Escherichia coli Shiga toxins induce apoptosis in epithelial cells that is regulated by the Bcl-2 family. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 278, G811–G819. [Google Scholar] [PubMed]
- Svensson, M.; Lindstedt, R.; Radin, N.S.; Svanborg, C. Epithelial glycosphingolipid expression as a determinant of bacterial adherence and cytokine production. Infect. Immun. 1994, 62, 4404–4410. [Google Scholar] [PubMed]
- Nakanishi, K.; Morikane, S.; Ichikawa, S.; Kurohane, K.; Niwa, Y.; Akimoto, Y.; Matsubara, S.; Kawakami, H.; Kobayashi, H.; Imai, Y. Protection of human colon cells from Shiga toxin by plant-based recombinant secretory IgA. Sci. Rep. 2017, 7, 45843. [Google Scholar] [CrossRef] [PubMed]
- Kovbasnjuk, O.; Edidin, M.; Donowitz, M. Role of lipid rafts in Shiga toxin 1 interaction with the apical surface of Caco-2 cells. J. Cell Sci. 2001, 114, 4025–4031. [Google Scholar] [PubMed]
- Shimizu, T.; Hamabata, T.; Yoshiki, A.; Hori, T.; Ito, S.; Takeda, Y.; Hayashi, H. An association of 27- and 40-kDa molecules with glycolipids that bind A-B bacterial enterotoxins to cultured cells. Biochim. Biophys. Acta 2003, 1612, 186–194. [Google Scholar] [CrossRef]
- Delmas, O.; Breton, M.; Sapin, C.; Le Bivic, A.; Colard, O.; Trugnan, G. Heterogeneity of raft-type membrane microdomains associated with VP4, the rotavirus spike protein in Caco-2 and MA 104 cells. J. Virol. 2007, 81, 1610–1618. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Tamiya-Koizumi, K.; Yamada, M.; Murate, T.; Kannagi, R.; Kyogashima, M. Individual profiles of free ceramide species and the constituent ceramide species of sphingomyelin and neutral glycosphingolipid and their alteration according to the sequential changes of environmental oxygen content in human colorectal cancer Caco-2 cells. Glycoconj. J. 2014, 31, 209–219. [Google Scholar] [PubMed]
- Müthing, J.; Pörtner, A.; Jäger, V. Ganglioside alterations in YAC-1 cells cultivated in serum-supplemented and serum-free growth medium. Glycoconj. J. 1992, 9, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.E.; Kane, A.V.; Campbell, S.T.; Acheson, D.W.K.; Cochran, B.H.; Thorpe, C.M. Shiga toxin 1 triggers a ribotoxic stress response leading to p38 and JNK activation and induction of apoptosis in intestinal epithelial cells. Infect. Immun. 2003, 71, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Karve, S.S.; Weiss, A.A. Glycolipid binding preferences of Shiga toxin variants. PLoS ONE 2014, 9, e101173. [Google Scholar] [CrossRef] [PubMed]
- Zumbrun, S.D.; Melton-Celsa, A.R.; Smith, M.A.; Gilbreath, J.J.; Merrell, D.S.; O’Brien, A.D. Dietary choice affects Shiga toxin-producing Escherichia coli (STEC) O157:H7 colonization and disease. Proc. Natl. Acad. Sci. USA 2013, 110, E2126–E2133. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.J.; Thorpe, C.M.; Paton, A.W.; Paton, J.C. Role of lipid rafts and flagellin in invasion of colonic epithelial cells by Shiga-toxigenic Escherichia coli O113:H21. Infect. Immun. 2012, 80, 2858–2867. [Google Scholar] [CrossRef] [PubMed]
- Kouzel, I.U.; Soltwisch, J.; Pohlentz, G.; Schmitz, J.S.; Karch, H.; Dreisewerd, K.; Müthing, J. Infrared MALDI mass spectrometry imaging of TLC-separated glycosphingolipids with emphasis on Shiga toxin receptors isolated from human colon epithelial cells. Int. J. Mass Spectrom. 2017, 416, 53–60. [Google Scholar] [CrossRef]
- Philpott, D.J.; Ackerley, C.A.; Kiliaan, A.J.; Karmali, M.A.; Perdue, M.H.; Sherman, P.M. Translocation of verotoxin-1 across T84 monolayers: Mechanism of bacterial toxin penetration of epithelium. Am. J. Physiol. 1997, 273, G1349–G1358. [Google Scholar] [PubMed]
- Windschiegl, B.; Orth, A.; Römer, W.; Berland, L.; Stechmann, B.; Bassereau, P.; Johannes, L.; Steinem, C. Lipid reorganization induced by Shiga toxin clustering on planar membranes. PLoS ONE 2009, 4, e6238. [Google Scholar] [CrossRef] [PubMed]
- Watkins, E.B.; Gao, H.; Dennison, A.C.J.; Chopin, N.; Struth, B.; Arnold, T.; Florent, J.C.; Johannes, L. Carbohydrate conformation and lipid condensation in monolayers containing glycosphingolipid Gb3: Influence of acyl chain structure. Biophys. J. 2014, 107, 1146–1155. [Google Scholar] [CrossRef] [PubMed]
- Schütte, O.M.; Patalag, L.J.; Weber, L.M.; Ries, A.; Römer, W.; Werz, D.B.; Steinem, C. 2-Hydroxy fatty acid enantiomers of Gb3 impact Shiga toxin binding and membrane organization. Biophys. J. 2015, 108, 2775–2778. [Google Scholar] [CrossRef] [PubMed]
- Pezeshkian, W.; Chaban, V.V.; Johannes, L.; Shillcock, J.; Ipsen, J.H.; Khandelia, H. The effects of globotriaosylceramide tails saturation level on bilayer phases. Soft Matter 2015, 11, 1352–1361. [Google Scholar] [CrossRef] [PubMed]
- Solovyeva, V.; Johannes, L.; Simonsen, A.C. Shiga toxin induces membrane reorganization and formation of long range lipid order. Soft Matter 2015, 11, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Chinnapen, D.J.; Hsieh, W.T.; te Welscher, Y.M.; Saslowsky, D.E.; Kaoutzani, L.; Brandsma, E.; D‘Auria, L.; Park, H.; Wagner, J.S.; Drake, K.R.; et al. Lipid sorting by ceramide structure from plasma membrane to ER for the cholera toxin receptor ganglioside GM1. Dev. Cell 2012, 23, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, A.W.; Bielaszewska, M.; Zhang, W.L.; Pulz, M.; Kuczius, T.; Ammon, A.; Karch, H. Escherichia coli harboring Shiga toxin 2 gene variants: Frequency and association with clinical symptoms. J. Infect. Dis. 2002, 185, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Werber, D.; Fruth, A.; Buchholz, U.; Prager, R.; Kramer, M.H.; Ammon, A.; Tschäpe, H. Strong association between Shiga toxin-producing Escherichia coli O157 and virulence genes stx2 and eae as possible explanation for predominance of serogroup O157 in patients with haemolytic uraemic syndrome. Eur. J. Clin. Microbiol. Infect. Dis. 2003, 22, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Ammon, A.; Petersen, L.R.; Karch, H. A large outbreak of hemolytic uremic syndrome caused by an unusual sorbitol-fermenting strain of Escherichia coli. J. Infect. Dis. 1999, 179, 1274–1277. [Google Scholar] [CrossRef] [PubMed]
- Tarr, P.I.; Neill, M.A. Escherichia coli O157:H7. Gastroenterol. Clin. N. Am. 2001, 30, 735–751. [Google Scholar] [CrossRef]
- Leopold, S.R.; Magrini, V.; Holt, N.J.; Shaikh, N.; Mardis, E.R.; Cagno, J.; Ogura, Y.; Iguchi, A.; Hayashi, T.; Mellmann, A.; et al. A precise reconstruction of the emergence and constrained radiations of Escherichia coli O157 portrayed by backbone concatenomic analysis. Proc. Natl. Acad. Sci. USA 2009, 106, 8713–8718. [Google Scholar] [CrossRef] [PubMed]
- Rusconi, B.; Sanjar, F.; Koenig, S.S.; Mammel, M.K.; Tarr, P.I.; Eppinger, M. Whole genome sequencing for genomics-guided investigations of Escherichia coli O157:H7 outbreaks. Front. Microbiol. 2016, 7, 985. [Google Scholar] [CrossRef] [PubMed]
- Bielaszewska, M.; Mellmann, A.; Zhang, W.; Köck, R.; Fruth, A.; Bauwens, A.; Peters, G.; Karch, H. Characterisation of the Escherichia coli strain associated with an outbreak of haemolytic uraemic syndrome in Germany, 2011: A microbiological study. Lancet Infect. Dis. 2011, 11, 671–676. [Google Scholar] [CrossRef]
- Karch, H.; Denamur, E.; Dobrindt, U.; Finlay, B.B.; Hengge, R.; Johannes, L.; Ron, E.Z.; Tønjum, T.; Sansonetti, P.J.; Vicente, M. The enemy within us: Lessons from the 2011 European Escherichia coli O104:H4 outbreak. EMBO Mol. Med. 2012, 4, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Bauwens, A.; Bielaszewska, M.; Kemper, B.; Langehanenberg, P.; von Bally, G.; Reichelt, R.; Mulac, D.; Humpf, H.U.; Friedrich, A.W.; Kim, K.S.; et al. Differential cytotoxic actions of Shiga toxin 1 and Shiga toxin 2 on microvascular and macrovascular endothelial cells. Thromb. Haemost. 2011, 105, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Ledeen, R.W.; Yu, R.K. Gangliosides: Structure, isolation, and analysis. Methods Enzymol. 1982, 83, 139–191. [Google Scholar] [PubMed]
- Müthing, J.; Egge, H.; Kniep, B.; Mühlradt, P.F. Structural characterization of gangliosides from murine T lymphocytes. Eur. J. Biochem. 1987, 163, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A.; Rose, J.K. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell 1992, 68, 533–544. [Google Scholar] [CrossRef]
- Meisen, I.; Friedrich, A.W.; Karch, H.; Witting, U.; Peter-Katalinić, J.; Müthing, J. Application of combined high-performance thin-layer chromatography immunostaining and nanoelectrospray ionisation quadrupole time-of-flight tandem mass spectrometry to the structural characterization of high- and low-affinity binding ligands of Shiga toxin 1. Rapid Commun. Mass Spectrom. 2005, 19, 3659–3665. [Google Scholar] [PubMed]
- Distler, U.; Hülsewig, M.; Souady, J.; Dreisewerd, K.; Haier, J.; Senninger, N.; Friedrich, A.W.; Karch, H.; Hillenkamp, F.; Berkenkamp, S.; et al. Matching IR-MALDI-o-TOF mass spectrometry with the TLC overlay binding assay and its clinical application for tracing tumor-associated glycosphingolipids in hepatocellular and pancreatic cancer. Anal. Chem. 2008, 80, 1835–1846. [Google Scholar] [CrossRef] [PubMed]
- Souady, J.; Soltwisch, J.; Dreisewerd, K.; Haier, J.; Peter-Katalinić, J.; Müthing, J. Structural profiling of individual glycosphingolipids in a single thin-layer chromatogram by multiple sequential immunodetection matched with direct IR-MALDI-o-TOF mass spectrometry. Anal. Chem. 2009, 81, 9481–9492. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, P.; Hülsewig, M.; Duvar, S.; Ziehr, H.; Mormann, M.; Peter-Katalinić, J.; Friedrich, A.W.; Karch, H.; Müthing, J. On the structural diversity of Shiga toxin glycosphingolipid receptors in lymphoid and myeloid cells determined by nanoelectrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2295–2304. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Hakomori, S.I. Quantitative isolation of total glycosphingolipids from animal cells. J. Lipid Res. 1971, 12, 257–259. [Google Scholar] [PubMed]
- Chester, M.A. IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). Nomenclature of glycolipids-recommendations 1997. Eur. J. Biochem. 1998, 257, 293–298. [Google Scholar] [PubMed]
- Meisen, I.; Rosenbrück, R.; Galla, H.J.; Hüwel, S.; Kouzel, I.U.; Mormann, M.; Karch, H.; Müthing, J. Expression of Shiga toxin 2e glycosphingolipid receptors of primary porcine brain endothelial cells and toxin-mediated breakdown of the blood-brain barrier. Glycobiology 2013, 23, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Kouzel, I.U.; Pirkl, A.; Pohlentz, G.; Soltwisch, J.; Dreisewerd, K.; Karch, H.; Müthing, J. Progress in detection and structural characterization of glycosphingolipids in crude lipid extracts by enzymatic phospholipid disintegration combined with thin-layer chromatography immunodetection and IR-MALDI mass spectrometry. Anal. Chem. 2014, 86, 1215–1222. [Google Scholar] [CrossRef] [PubMed]
- Kean, E.L. Separation of gluco- and galactocerebrosides by means of borate thin-layer chromatography. J. Lipid Res. 1966, 7, 449–452. [Google Scholar] [PubMed]
- Dittmer, J.C.; Lester, R.L. A simple, specific spray for the detection of phospholipids on thin-layer chromatograms. J. Lipid Res. 1964, 5, 126–127. [Google Scholar] [PubMed]
- Müthing, J.; Radloff, M. Nanogram detection of phospholipids on thin-layer chromatograms. Anal. Biochem. 1998, 257, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Goswami, S.K.; Frey, C.F. Manganous chloride spray reagent for cholesterol and bile acids on thin-layer chromatograms. J. Chromatogr. 1970, 53, 389–390. [Google Scholar] [CrossRef]
- Betz, J.; Dorn, I.; Kouzel, I.U.; Bauwens, A.; Meisen, I.; Kemper, B.; Bielaszewska, M.; Mormann, M.; Weymann, L.; Sibrowski, W.; et al. Shiga toxin of enterohemorrhagic Escherichia coli directly injures developing human erythrocytes. Cell. Microbiol. 2016, 18, 1339–1348. [Google Scholar] [CrossRef] [PubMed]
- Meisen, I.; Peter-Katalinić, J.; Müthing, J. Direct analysis of silica gel extracts from immunostained glycosphingolipids by nanoelectrospray ionization quadrupole time-of-flight mass spectrometry. Anal. Chem. 2004, 76, 2248–2255. [Google Scholar] [CrossRef] [PubMed]
- Domon, B.; Costello, C.E. A systematic nomenclature for carbohydrate fragmentations in FAB-MS/MS spectra of glycoconjugates. Glycoconj. J. 1988, 5, 397–440. [Google Scholar] [CrossRef]
- Domon, B.; Costello, C.E. Structure elucidation of glycosphingolipids and gangliosides using high-performance tandem mass spectrometry. Biochemistry 1988, 27, 1534–1543. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound b | Ceramide | Formula | m/zexp | m/zcalc |
|---|---|---|---|---|
| SM ([M + H]+) | d18:1, C16:0 | C39H80N2O6P | 703.58 | 703.5754 |
| SM | d18:1, C16:0 | C39H79N2O6PNa | 725.57 | 725.5573 |
| SM | d18:1, C24:1 | C47H93N2O6PNa | 835.67 | 835.6669 |
| SM | d18:1, C24:0 | C47H95N2O6PNa | 837.69 | 837.6825 |
| Lc2Cer | d18:1, C16:0 | C46H87NO13Na | 884.61 | 884.6075 |
| Lc2Cer | d18:1, C22:0 | C52H99NO13Na | 968.71 | 968.7014 |
| Lc2Cer | d18:1, C24:1 | C54H101NO13Na | 994.73 | 994.7171 |
| Lc2Cer | d18:1, C24:0 | C54H103NO13Na | 996.74 | 996.7327 |
| Lc2Cer | d18:1, C24:1-OH | C54H101NO14Na | 1010.72 | 1010.7120 |
| Lc2Cer | d18:1, C24:0-OH | C54H103NO14Na | 1012.73 | 1012.7276 |
| Gb3Cer | d18:1, C16:0 | C52H97NO18Na | 1046.66 | 1046.6603 |
| Gb3Cer | d18:1, C22:0 | C58H109NO18Na | 1130.76 | 1130.7542 |
| Gb3Cer | d18:1, C24:1 | C60H111NO18Na | 1156.78 | 1156.7699 |
| Gb3Cer | d18:1, C24:0 | C60H113NO18Na | 1158.78 | 1158.7855 |
| Gb4Cer | d18:1, C16:0 | C60H110N2O23Na | 1249.75 | 1249.7397 |
| Gb4Cer | d18:1, C22:0 | C66H122N2O23Na | 1333.84 | 1333.8336 |
| Gb4Cer | d18:1, C24:1 | C68H124N2O23Na | 1359.86 | 1359.8493 |
| Gb4Cer | d18:1, C24:0 | C68H126N2O23Na | 1361.87 | 1361.8649 |
| Compound b | Ceramide | Formula | m/zexp | m/zcalc |
|---|---|---|---|---|
| SM ([M + H]+) | d18:1, C16:0 | C39H80N2O6P | 703.57 | 703.5754 |
| SM | d18:1, C16:0 | C39H79N2O6PNa | 725.55 | 725.5573 |
| SM | d18:1, C24:1 | C47H93N2O6PNa | 835.67 | 835.6669 |
| SM | d18:1, C24:0 | C47H95N2O6PNa | 837.68 | 837.6825 |
| Lc2Cer | d18:1, C16:0 | C46H87NO13Na | 884.60 | 884.6075 |
| Lc2Cer | d18:1, C22:0 | C52H99NO13Na | 968.70 | 968.7014 |
| Lc2Cer | d18:1, C22:0-OH | C52H99NO14Na | 984.69 | 984.6963 |
| Lc2Cer | d18:1, C24:1 | C54H101NO13Na | 994.70 | 994.7171 |
| Lc2Cer | d18:1, C24:0 | C54H103NO13Na | 996.73 | 996.7327 |
| Lc2Cer | d18:1, C24:1-OH | C54H101NO14Na | 1010.71 | 1010.7120 |
| Lc2Cer | d18:1, C24:0-OH | C54H103NO14Na | 1012.73 | 1012.7276 |
| Gb3Cer | d18:1, C16:0 | C52H97NO18Na | 1046.65 | 1046.6603 |
| Gb3Cer | d18:1, C22:0 | C58H109NO18Na | 1130.75 | 1130.7542 |
| Gb3Cer | d18:1, C24:1 | C60H111NO18Na | 1156.76 | 1156.7699 |
| Gb3Cer | d18:1, C24:0 | C60H113NO18Na | 1158.78 | 1158.7855 |
| Gb4Cer | d18:1, C16:0 | C60H110N2O23Na | 1249.74 | 1249.7397 |
| Gb4Cer | t18:0, C16:0 | C60H112N2O24Na | 1267.75 | 1267.7502 |
| Gb4Cer | d18:1, C22:0 | C66H122N2O23Na | 1333.83 | 1333.8336 |
| Gb4Cer | t18:0, C22:0 | C66H124N2O24Na | 1351.84 | 1351.8441 |
| Gb4Cer | d18:1, C24:1 | C68H124N2O23Na | 1359.84 | 1359.8493 |
| Gb4Cer | d18:1, C24:0 | C68H126N2O23Na | 1361.86 | 1361.8649 |
| Gb4Cer | t18:0, C24:1 | C68H126N2O24Na | 1377.85 | 1377.8598 |
| Gb4Cer | t18:0, C24:0 | C68H128N2O24Na | 1379.86 | 1379.8598 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kouzel, I.U.; Pohlentz, G.; Schmitz, J.S.; Steil, D.; Humpf, H.-U.; Karch, H.; Müthing, J. Shiga Toxin Glycosphingolipid Receptors in Human Caco-2 and HCT-8 Colon Epithelial Cell Lines. Toxins 2017, 9, 338. https://doi.org/10.3390/toxins9110338
Kouzel IU, Pohlentz G, Schmitz JS, Steil D, Humpf H-U, Karch H, Müthing J. Shiga Toxin Glycosphingolipid Receptors in Human Caco-2 and HCT-8 Colon Epithelial Cell Lines. Toxins. 2017; 9(11):338. https://doi.org/10.3390/toxins9110338
Chicago/Turabian StyleKouzel, Ivan U., Gottfried Pohlentz, Julia S. Schmitz, Daniel Steil, Hans-Ulrich Humpf, Helge Karch, and Johannes Müthing. 2017. "Shiga Toxin Glycosphingolipid Receptors in Human Caco-2 and HCT-8 Colon Epithelial Cell Lines" Toxins 9, no. 11: 338. https://doi.org/10.3390/toxins9110338
APA StyleKouzel, I. U., Pohlentz, G., Schmitz, J. S., Steil, D., Humpf, H. -U., Karch, H., & Müthing, J. (2017). Shiga Toxin Glycosphingolipid Receptors in Human Caco-2 and HCT-8 Colon Epithelial Cell Lines. Toxins, 9(11), 338. https://doi.org/10.3390/toxins9110338