
Can Inhibitors of Snake Venom Phospholipases A2 Lead to New Insights into Anti-Inflammatory Therapy in Humans? A Theoretical Study


 and
and 
Abstract
:1. Introduction
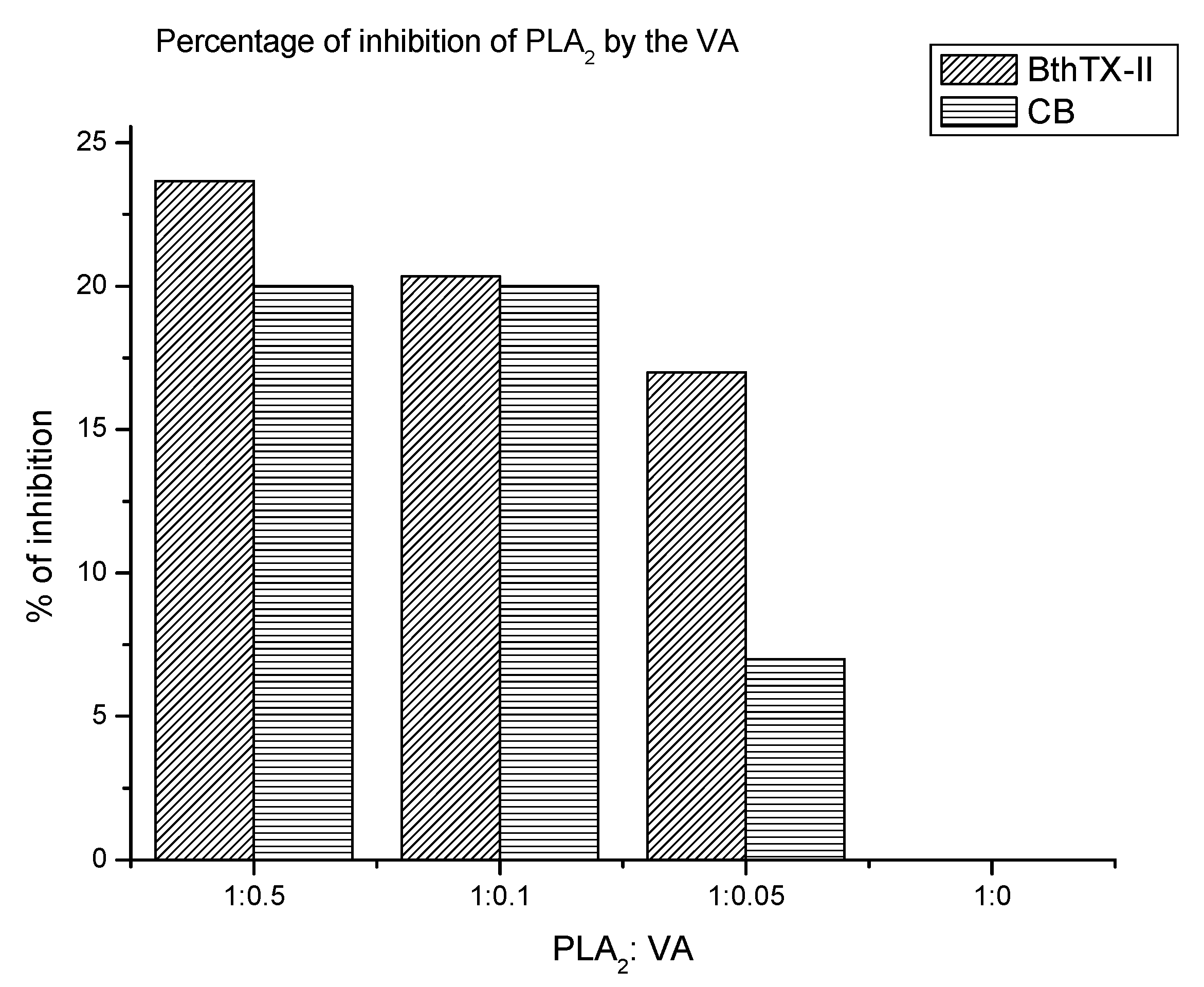
2. Results
2.1. Experimental Assays
2.2. Alignments of Amino Acid Sequences of svPLA2 and Human PLA2
2.3. Theoretical Calculations
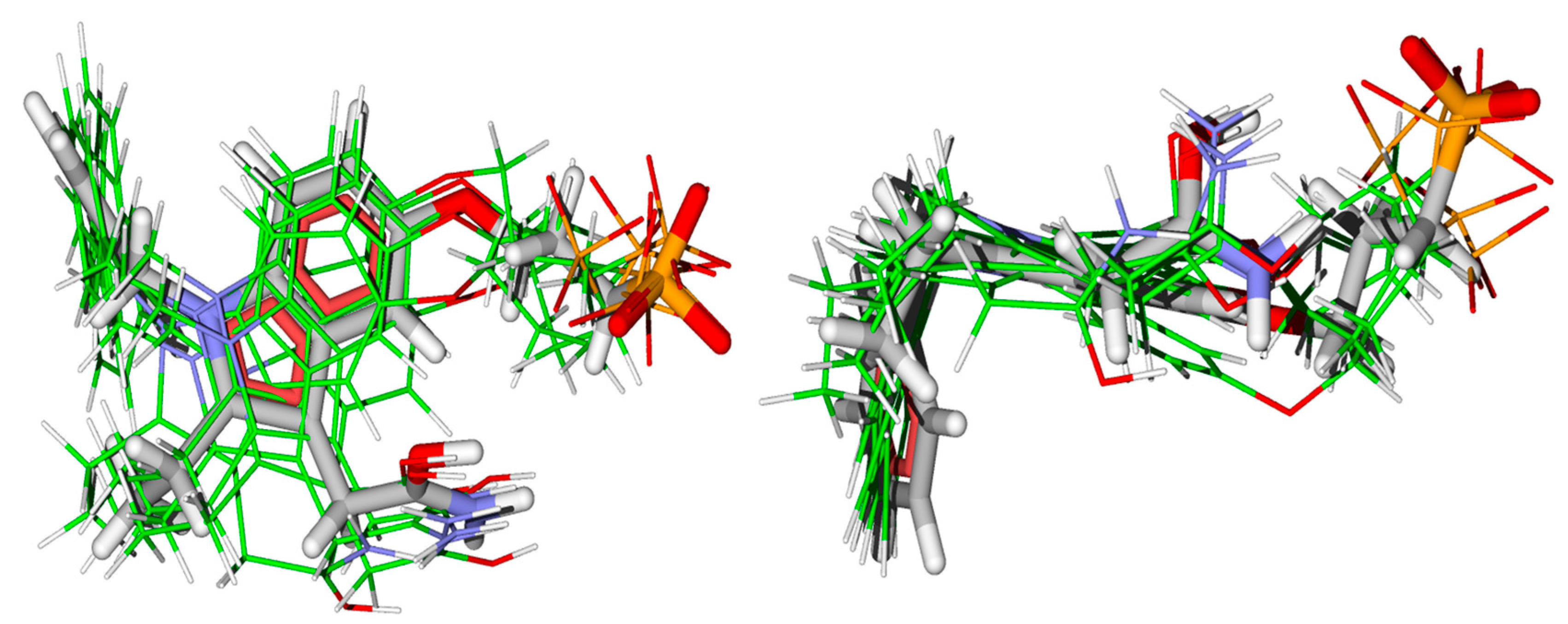
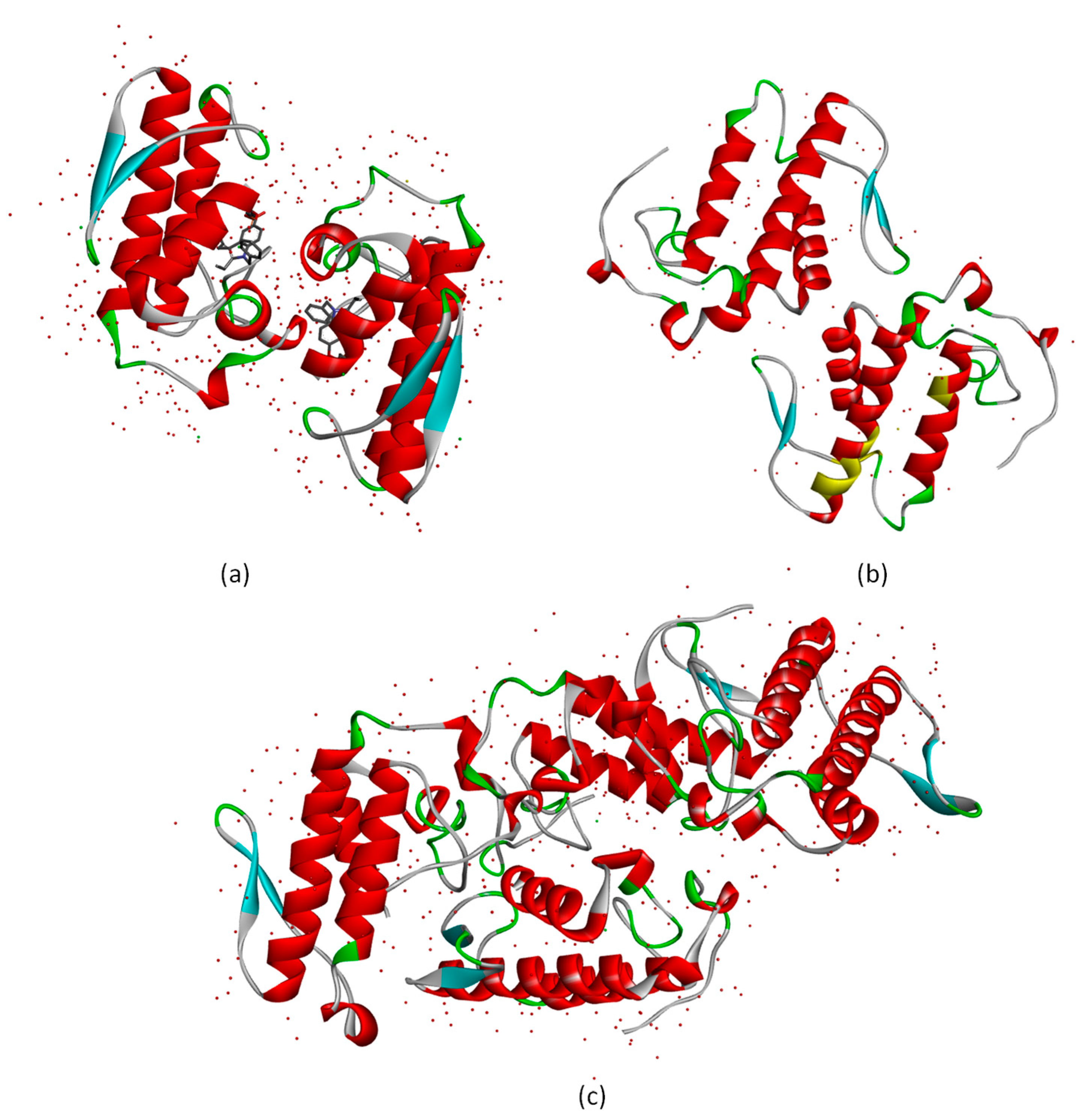
2.3.1. Molecular Docking Calculations
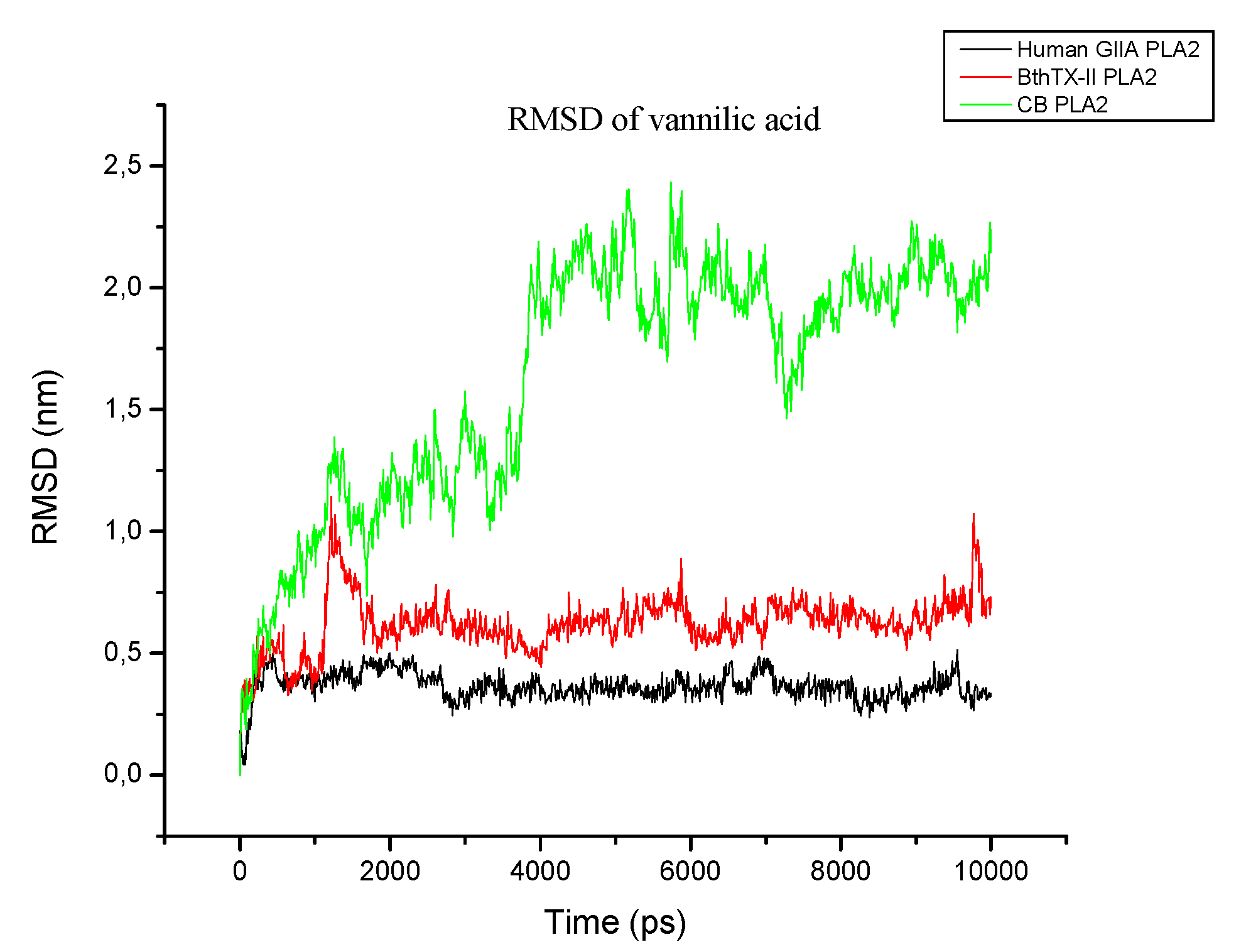
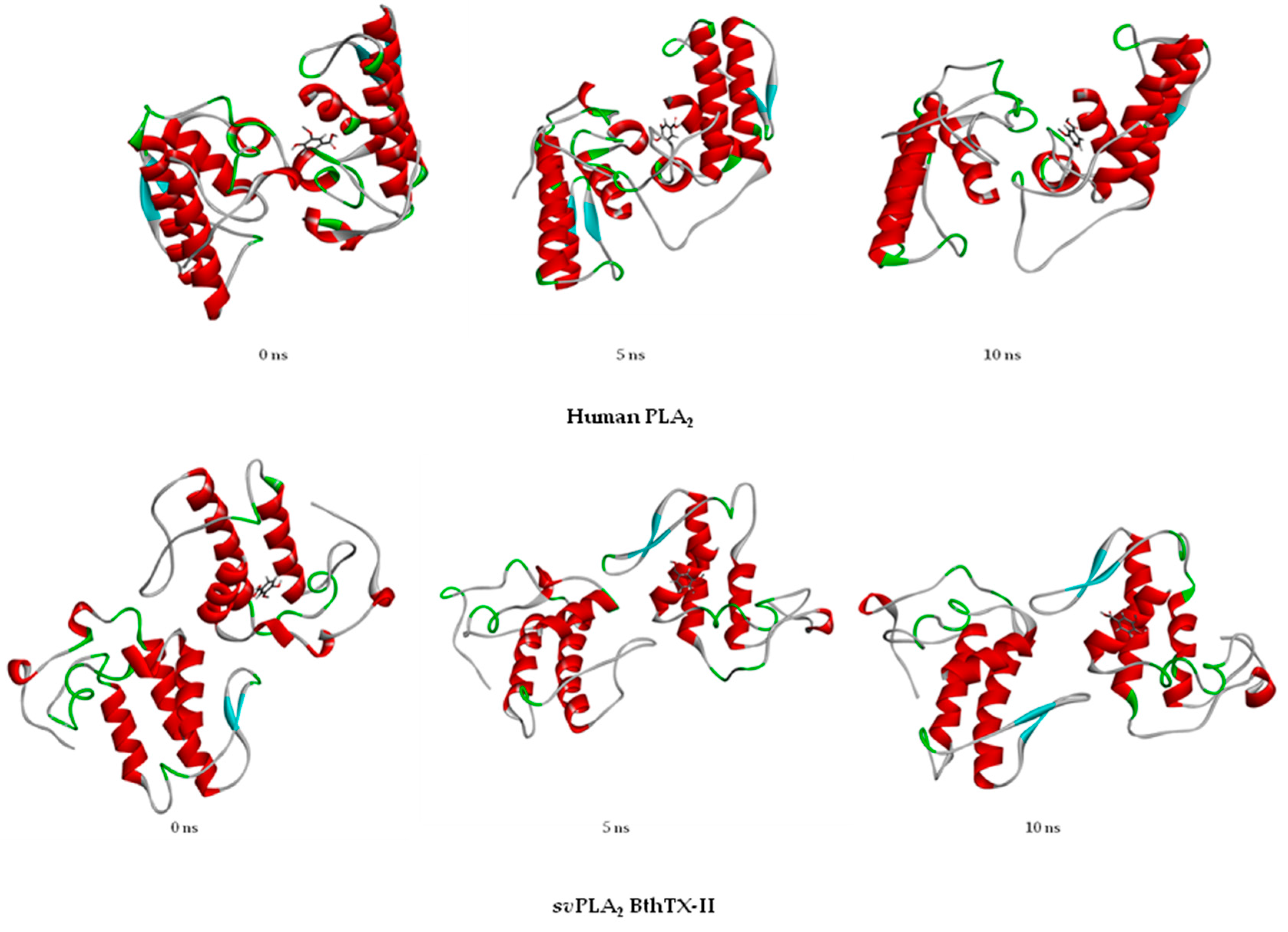
2.3.2. Molecular Dynamics Simulation
3. Discussion
3.1. Can svPLA2/Inhibitors Describe the hPLA2/Inhibitors Interactions?
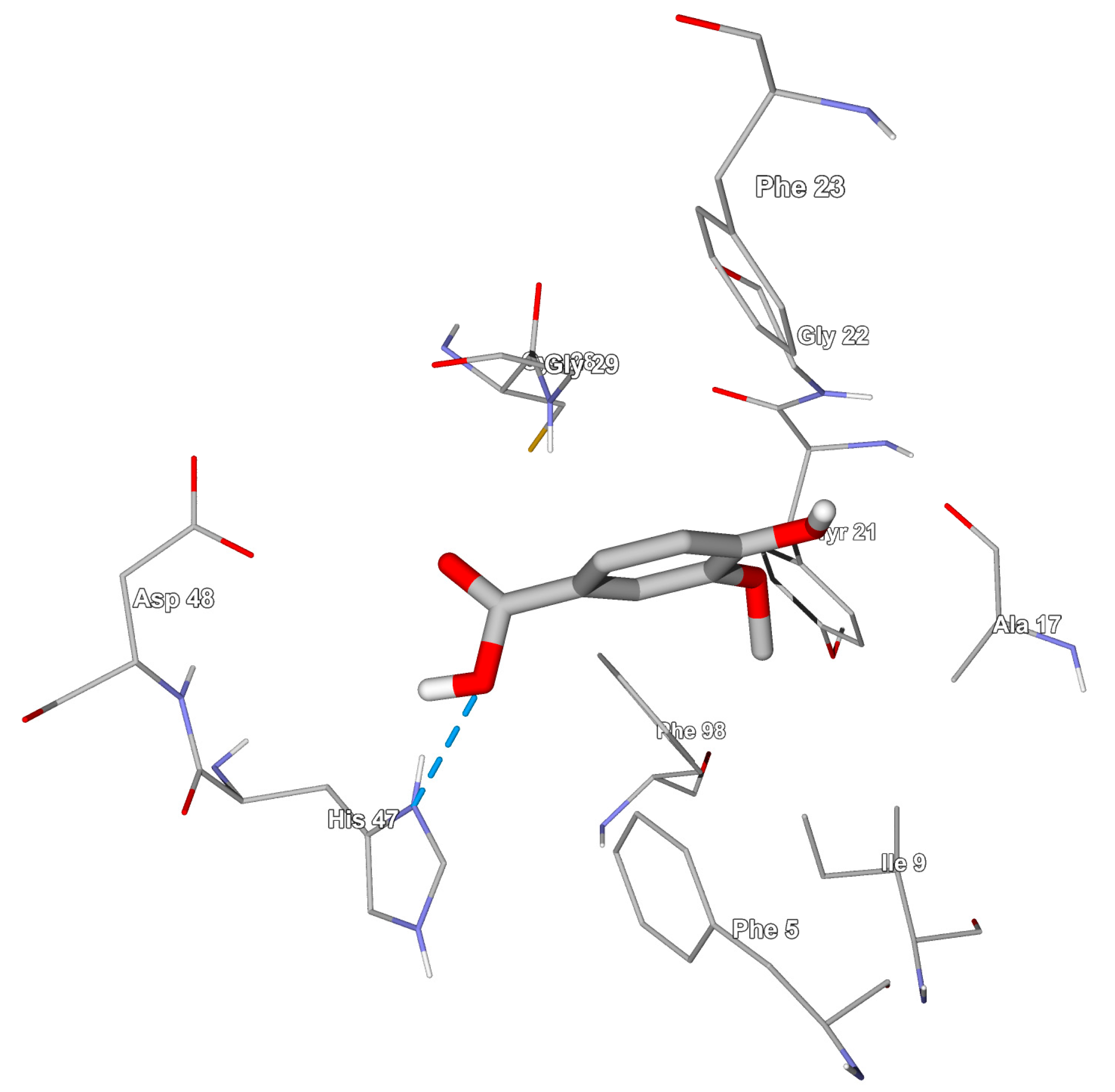
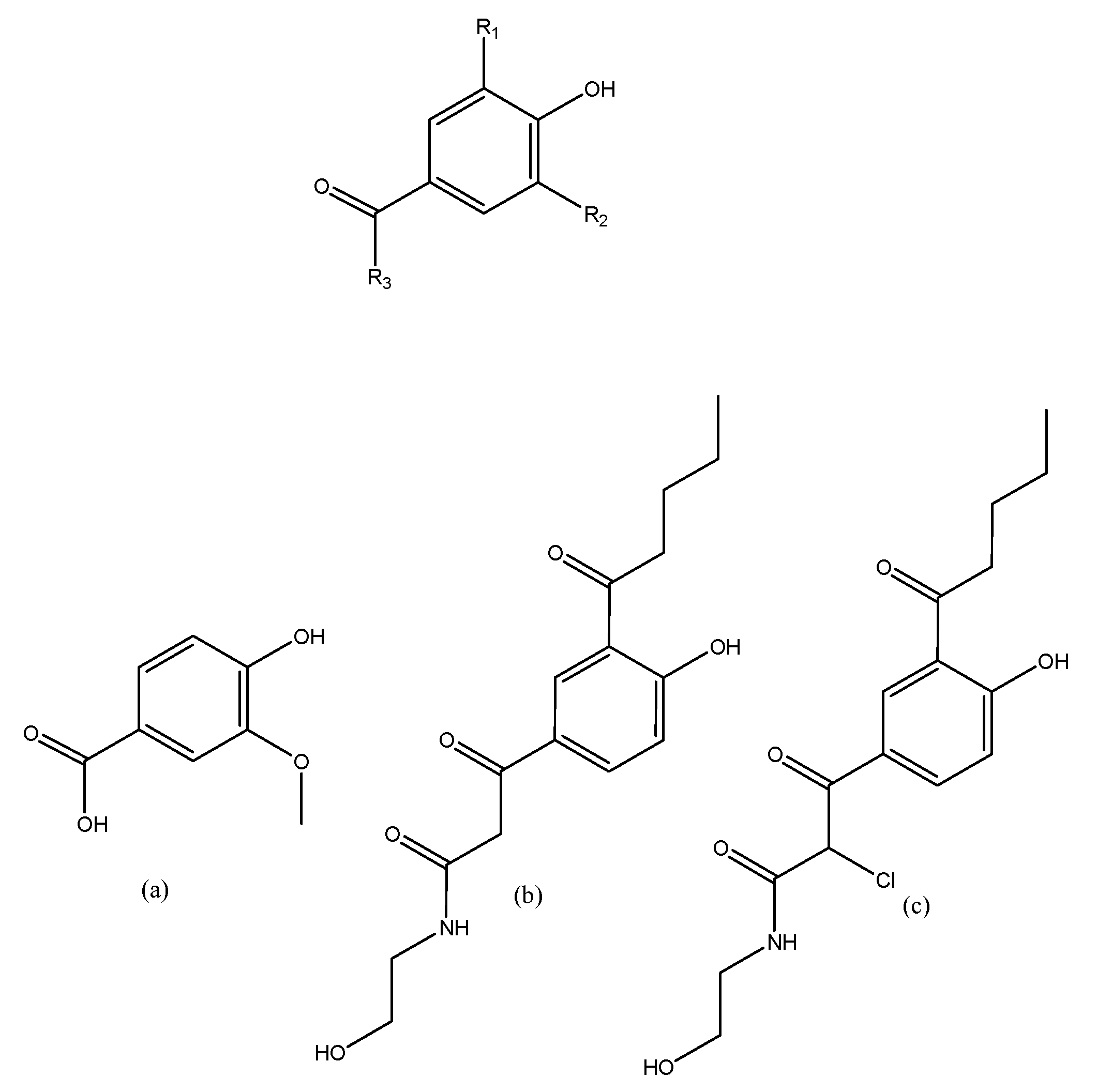
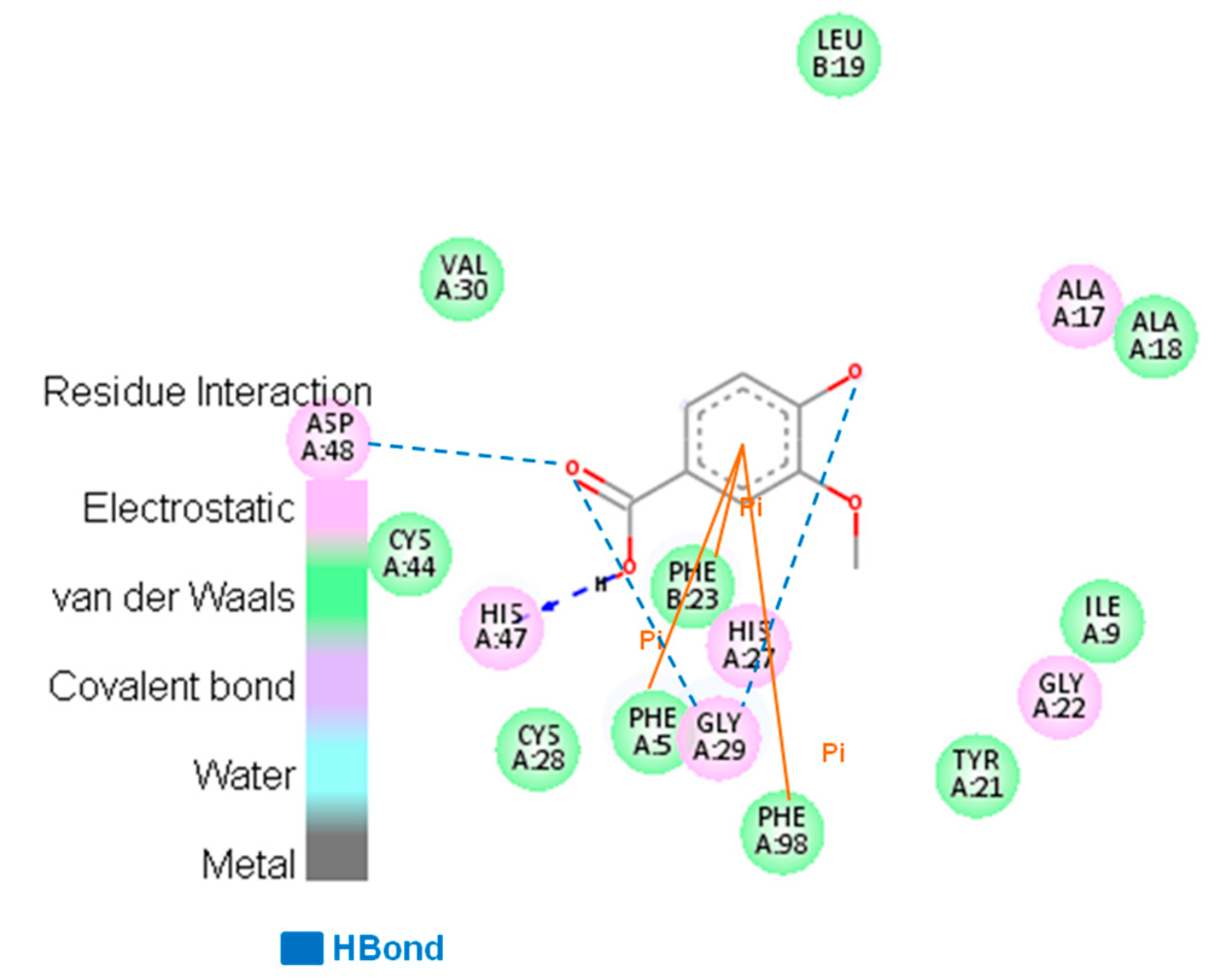
3.2. Searching Molecular Interactions of Vanillic Acid Analogs
4. Conclusions
5. Materials and Methods
5.1. Experimental Assays
5.2. Alignments of Amino Acid Sequences
5.3. Simulation Methods
5.3.1. Docking Energies Calculations
5.3.2. Molecular Dynamics Simulations
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Joshi, V. Dimethyl ester of bilirubin exhibits anti-inflammatory activity through inhibition of secretory phospholipase A2, lipoxygenase and cyclooxygenase. Arch. Biochem. Biophys. 2016, 598, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Silva, P. Farmacologia, 8th ed.; Guanabara Koogan: Rio de Janeiro, Brazil, 2010; 1024p. [Google Scholar]
- Muri, E.M.; Sposito, M.M.M.; Metsavaht, L. Nonsteroidal antiinflammatory drugs and their local pharmacology. Acta Fisiatr. 2009, 16, 186–190. [Google Scholar]
- Anelli, M.G.; Scioscia, C.; Grattagliano, I.; Lapadula, G. Old and new antirheumatic drugs and the risk of hepatotoxicity. Ther. Drug Monit. 2012, 34, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Rafaniello, C.; Ferrajolo, C.; Sullo, M.G.; Sessa, M.; Sportiello, L.; Balzano, A.; Manguso, F.; Aiezza, M.L.; Rossi, F.; Scarpignato, C.; et al. Risk of gastrointestinal complications associated to NSAIDs, low-dose aspirin and their combinations: Results of a pharmacovigilance reporting system. Pharm. Res. 2016, 104, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Yousefpour, A.; Iranagh, S.A.; Nademi, Y.; Modarress, H. Molecular dynamics simulation of nonsteroidal antiinflammatory drugs, naproxen and relafen, in a lipid bilayer membrane. Int. J. Quant. Chem. 2013, 113, 1919–1930. [Google Scholar] [CrossRef]
- Cronstein, B.N.; Weissmann, G. Targets for antiinflammatory drugs. Ann. Rev. Pharmacol. Toxicol. 1995, 35, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Gaddipati, R.S.; Raikundalia, G.K.; Mathai, M.L. Dual and selective lipid inhibitors of cyclooxygenases and lipoxygenase: A molecular docking study. Med. Chem. Res. 2014, 23, 3389–3402. [Google Scholar] [CrossRef]
- Peters-Golden, M.D.M.; Henderson, M.D.W.R., Jr. Leukotrienes. N. Engl. J. Med. 2007, 357, 1841–1854. [Google Scholar] [CrossRef] [PubMed]
- Pyasi, K.; Tufvesson, E.; Moitra, S. Evaluating the role of leukotriene-modifying drugs in asthma management: Are their benefits ‘losing in translation’? Pulm. Pharmacol. Ther. 2016, 41, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Quach, N.D.; Arnold, R.D.; Cummings, B.S. Secretory phospholipase A2 enzymes as pharmacological targets for treatment of disease. Biochem. Pharmacol. 2014, 90, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Tomankova, V.; Anzenbacher, P.; Anzenbacherova, E. Effects of obesity on liver cytochromes P450 in various animal models. Biomed. Pap. Olomouc 2017, 161, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Lerch, M.M.; Adler, G. Experimental animal-models of acute-pancreatitis. Int. J. Pancreatol. 1994, 15, 159–170. [Google Scholar] [PubMed]
- Liu, Y.; Zeng, B.H.; Shang, H.T.; Cen, Y.Y.; Wei, H. Bama Miniature Pigs (Sus scrofa domestica) as a Model for Drug Evaluation for Humans: Comparison of In Vitro Metabolism and In Vivo Pharmacokinetics of Lovastatin. Comp. Med. 2008, 58, 580–587. [Google Scholar] [PubMed]
- Prueksaritanont, T. Use of In Vivo Animal Models to Assess Drug-Drug Interactions. Enzyme- and Transporter-Based Drug-Drug Interactions: Progress and Future Challenges. Pharm. Res. 2010, 27, 283–297. [Google Scholar]
- Siltari, A.; Kivimäki, A.S.; Ehlers, P.I.; Korpela, R.; Vapaatalo, H. Effects of Milk Casein Derived Tripeptides on Endothelial Enzymes In Vitro; a Study with Synthetic Tripeptides. Arzneimittelforschung 2012, 62, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Marchi-Salvador, D.P.; Corrêa, L.C.; Magro, A.J.; Oliveira, C.Z.; Soares, A.M.; Fontes, M.R. Insights into the role of oligomeric state on the biological activities of crotoxin: Crystal structure of a tetrameric phospholipase A2 formed by two isoforms of crotoxin B from Crotalus durissus terrificus venom. Proteins 2008, 72, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, J.I.; Cintra-Francischinelli, M.; Borges, R.J.; Fernandes, C.A.; Pizzo, P.; Cintra, A.C.; Braz, A.S.; Soares, A.M.; Fontes, M.R. Structural, functional, and bioinformatics studies reveal a new snake venom homologue phospholipase A2 class. Proteins 2011, 79, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.F.; Landucci, E.C.; Antunes, E.; Chacur, M.; Cury, Y. Inflammatory effects of snake venom myotoxic phospholipases A2. Toxicon 2003, 42, 947–962. [Google Scholar] [CrossRef] [PubMed]
- Marcussi, S.; Sant’Ana, C.D.; Oliveira, C.Z.; Rueda, A.Q.; Menaldo, D.L.; Beleboni, R.O.; Stabeli, R.G.; Giglio, J.R.; Fontes, M.R.; Soares, A.M. Snake venom phospholipase A2 inhibitors: Medicinal chemistry and therapeutic potential. Curr. Top. Med. Chem. 2007, 7, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.R.; Malde, A.K.; Nematollahi, A.; Scott, K.F.; Church, W.B. Molecular dynamics simulations reveal structural insights into inhibitor binding modes and functionality in human Group IIA phospholipase A2. Proteins 2017, 85, 827–842. [Google Scholar] [CrossRef] [PubMed]
- Batsanov, S.S. Van der Waals Radii of Elements. Inorg. Mater. 2001, 37, 1031–1046. [Google Scholar]
- Da Cunha, E.E.; Barbosa, E.F.; Oliveira, A.A.; Ramalho, T.C. Molecular modeling of Mycobacterium tuberculosis DNA gyrase and its molecular docking study with gatifloxacin inhibitors. J. Biomol. Struct. Dyn. 2010, 27, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Golçalves, M.A.; Santos, L.S.; Prata, D.M.; Fernando, P.C.; Cunha, E.F.F.; Ramalho, T.C. Optimal wavelet signal compression as an efficient alternative to investigate molecular dynamics simulations: Aplication to thermal and solvent effects of MRI probes. Theor. Chem. Acc. 2017, 136, 15. [Google Scholar] [CrossRef]
- Dileep, K.V.; Remya, C.; Cerezo, J.; Fassihi, A.; Pérez-Sánchez, H.; Sadasivan, C. Comparative studies on the inhibitory activities of selected benzoic acid derivatives against secretory phospholipase A2, a key enzyme involved in the inflammatory pathway. Mol. Biosyst. 2015, 11, 1973–1979. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Avila, C.; Rojas, E.; Cerdas, L. An alternative in vitro method for testing the potency of the polyvalent antivenom produced in Costa Rica. Toxicon 1998, 26, 411–413. [Google Scholar] [CrossRef]
- Price, M.F.; Wilkinson, I.D.; Gentry, L.O. Plate method for detection of phospholipase activity in Candida albicans. Sabouraudia 1982, 20, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Miller, W.; Huang, X.; Miller, W. A time-efficient linear-space local similarity algorithm. Adv. Appl. Math. 1991, 12, 337–357. [Google Scholar] [CrossRef]
- Consortium, U. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef] [PubMed]
- Expasy. Available online: http://au.expasy.org/ (accessed on 23 October 2017).
- UniProtKB Database. Available online: http://www.uniprot.org/ (accessed on 23 October 2017).
- Hehre, W.J.; Deppmeier, B.J.; Klunzinger, P.E. Pcspartanpro; Wavefunction, Inc.: Irvine, CA, USA, 1999. [Google Scholar]
- Thomsen, R.; Christensen, M.H. MolDock: A new technique for high-accuracy Molecular docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
- Frisch, M.J. Gaussian 09; Gaussian, Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Canzar, S.; El-Kebir, M.; Pool, R.; Elbassioni, K.; Mark, A.E.; Geerke, D.P.; Stougie, L.; Klau, G.W. Charge group partitioning in biomolecular simulation. J. Comput. Biol. 2013, 20, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Automated Topology Builder (ATB) Server. Available online: http://compbio.biosci.uq.edu.au/atb/ (accessed on 23 October 2017).
- Van Gunsteren, W.F.; Billeter, S.R.; Eising, A.A.; Hunenberger, P.H.; Krüger, P.; Mark, A.E.; Scott, W.R.P.; Tironi, I.G. Biomolecular Simulation: The GROMOS96 Manual and User Guide; VDF Hochschulverlag AG an der ETH Zürich: Zurich, Switzerland, 1996. [Google Scholar]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Pall, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Caddigan, E.J. VMD User’s Guide; University of Illinois; Beckman Institute: Urbana, IL, USA, 2004. [Google Scholar]
- Edwards, P.M. Origin 7.0: Scientific graphing and data analysis software. J. Chem. Inf. Comput. Sci. Wash. 2002, 42, 1270–1271. [Google Scholar] [CrossRef]
- Mancini, D.T.; Matos, K.S.; da Cunha, E.F.; Assis, T.M.; Guimarães, A.P.; França, T.C.; Ramalho, T.C. Molecular modeling studies on nucleoside hydrolase from the biological warfare agent Brucella suis. J. Biomol. Struct. Dyn. 2012, 30, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Martins, T.L.C.; Ramalho, T.C.; Figueroa-Villar, J.D.; Flores, A.F.C.; Pereira, C.M.P. Theoretical and experimental C-13 and N-15 NMR investigation of guanylhydrazones in solution. Magn. Reson. Chem. 2013, 41, 983–988. [Google Scholar] [CrossRef]
- De Castro, A.A.; Prandi, I.G.; Kamil, K.; Ramalho, T.C. Organophosphorus degrading enzymes: Molecular basis and perspectives for enzymatic bioremediation of agrochemicals. Ciencia e Agrotecnologia 2017, 41, 471–482. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | MolDock Score | Rerank Score | Interaction | HBond |
|---|---|---|---|---|
| hPLA2 | ||||
| HGIIA | −69.38 | −58.93 | −75.35 | −2.21 |
| svPLA2 | ||||
| BthTX-II | −71.22 | −57.02 | −79.45 | 0.00 |
| CB | −37.87 | −35.17 | −44.91 | −0.02 |
| Compound | Enzyme | MolDock Score | Rerank Score | Interaction | HBond |
|---|---|---|---|---|---|
| Analogue I | HGIIA | −101.52 | −73.99 | −107.12 | −4.99 |
| Analogue I | BthTX-II | −111.32 | −69.64 | −113.82 | −4.61 |
| Analogue I | CB | −72.26 | 24.57 | −71.94 | −5.14 |
| Analogue II | HGIIA | −117.02 | −26.35 | −115.38 | −4.63 |
| Analogue II | BthTX-II | −123.33 | −101.81 | −126.35 | −7.91 |
| Analogue II | CB | −59.03 | 31.78 | −55.64 | −9.65 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sales, T.A.; Marcussi, S.; Da Cunha, E.F.F.; Kuca, K.; Ramalho, T.C. Can Inhibitors of Snake Venom Phospholipases A2 Lead to New Insights into Anti-Inflammatory Therapy in Humans? A Theoretical Study. Toxins 2017, 9, 341. https://doi.org/10.3390/toxins9110341
Sales TA, Marcussi S, Da Cunha EFF, Kuca K, Ramalho TC. Can Inhibitors of Snake Venom Phospholipases A2 Lead to New Insights into Anti-Inflammatory Therapy in Humans? A Theoretical Study. Toxins. 2017; 9(11):341. https://doi.org/10.3390/toxins9110341
Chicago/Turabian StyleSales, Thaís A., Silvana Marcussi, Elaine F. F. Da Cunha, Kamil Kuca, and Teodorico C. Ramalho. 2017. "Can Inhibitors of Snake Venom Phospholipases A2 Lead to New Insights into Anti-Inflammatory Therapy in Humans? A Theoretical Study" Toxins 9, no. 11: 341. https://doi.org/10.3390/toxins9110341
APA StyleSales, T. A., Marcussi, S., Da Cunha, E. F. F., Kuca, K., & Ramalho, T. C. (2017). Can Inhibitors of Snake Venom Phospholipases A2 Lead to New Insights into Anti-Inflammatory Therapy in Humans? A Theoretical Study. Toxins, 9(11), 341. https://doi.org/10.3390/toxins9110341