
Antivenom Evaluation by Electrophysiological Analysis

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Androctonus mauretanicus and Buthus mardochei Venoms Contain Sodium Channel Toxins
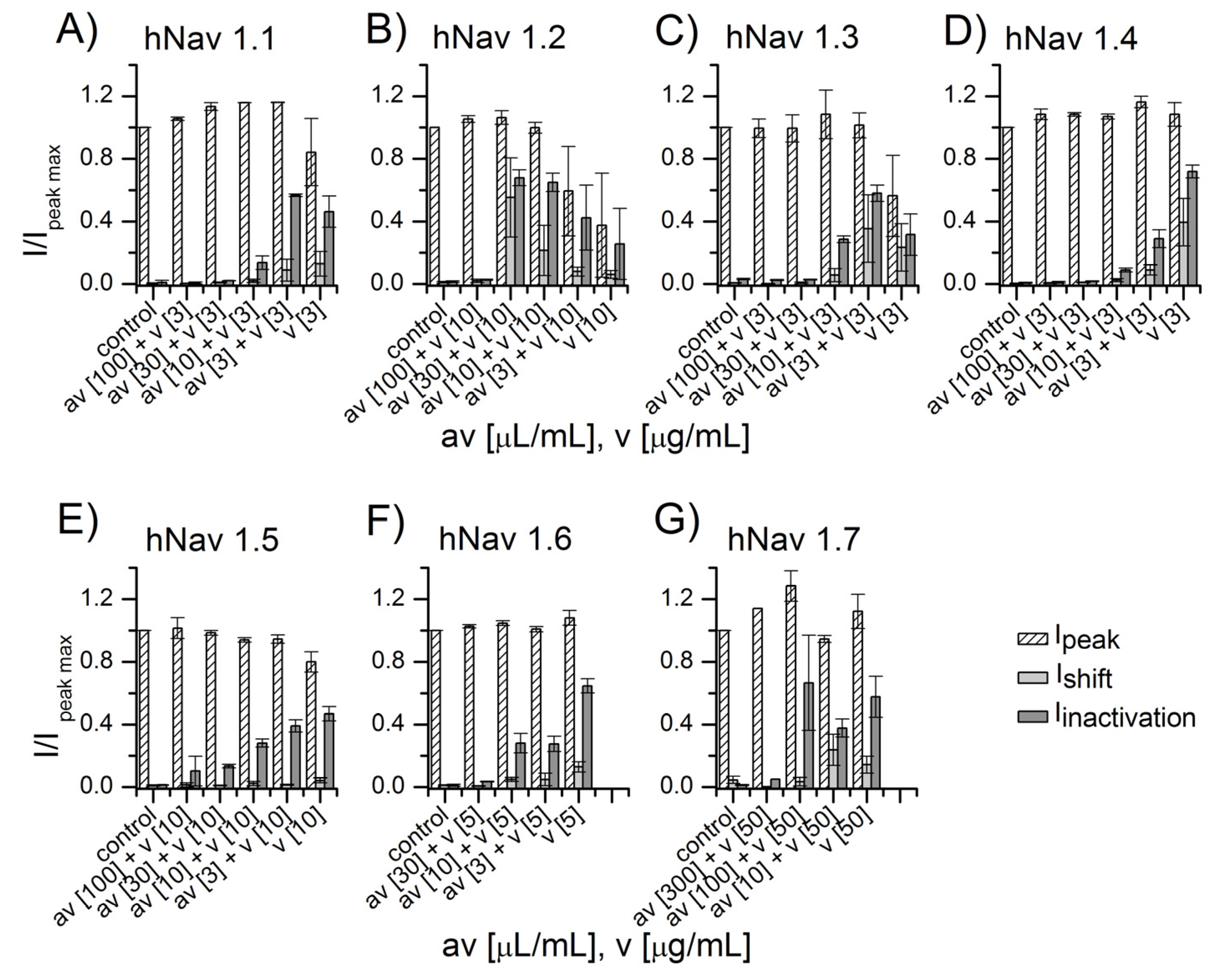
2.2. NA Scorpion Antivenom Neutralizes the Androctonus mauretanicus Venom Effect on Sodium Channels
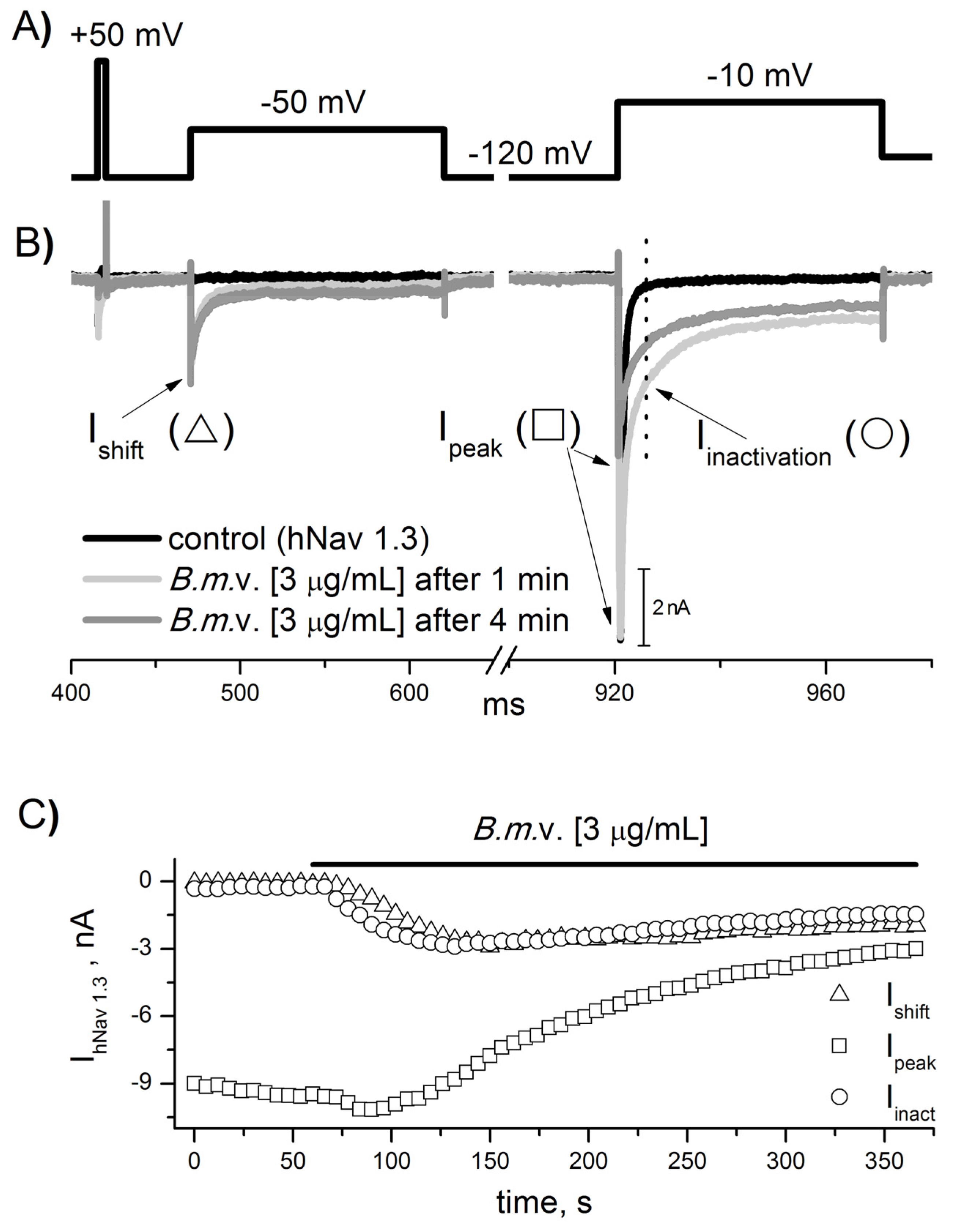
2.3. NA Scorpion Antivenom Neutralizes the Buthus mardochei Venom Effect on Sodium Channels
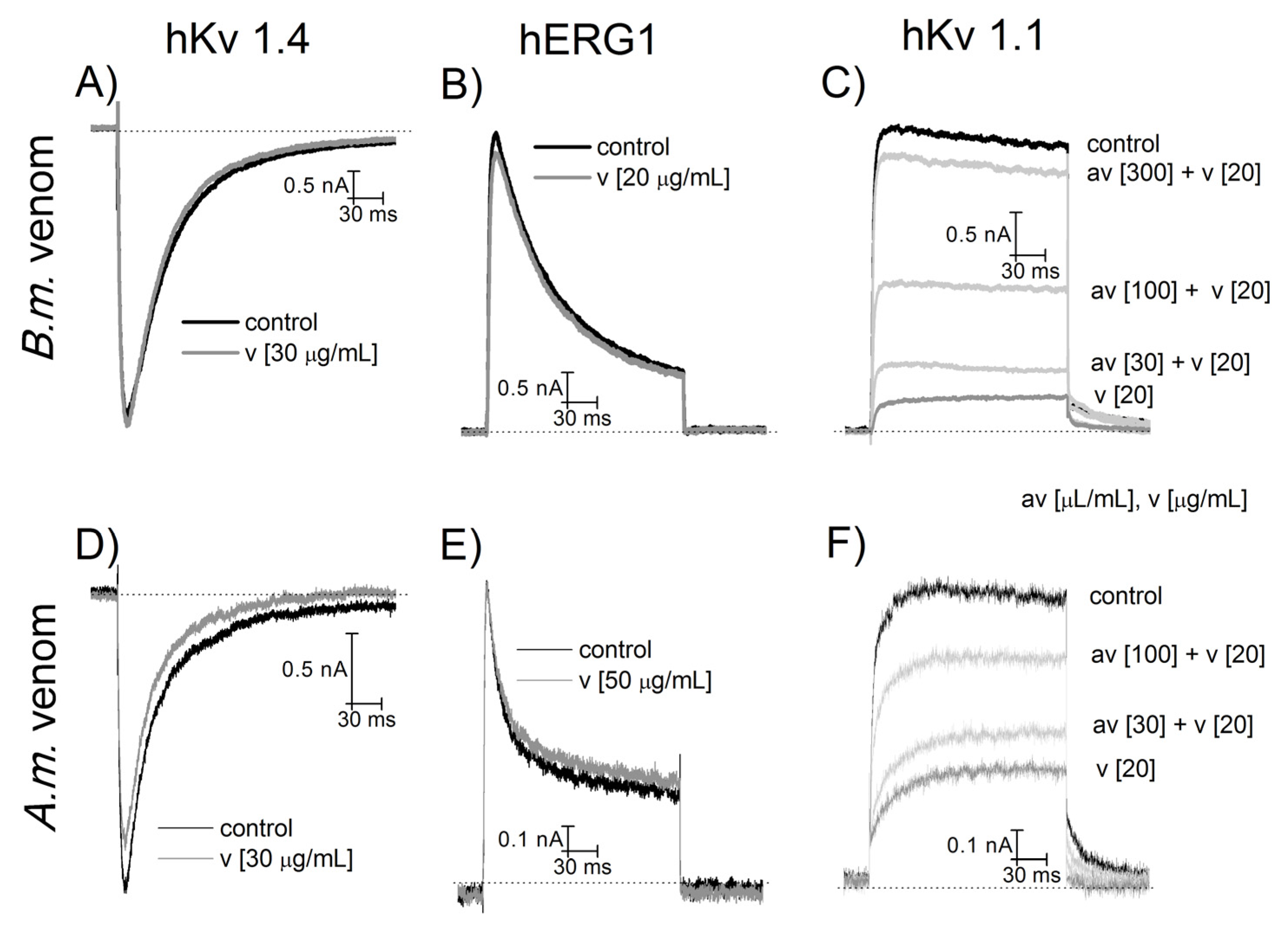
2.4. NA Scorpion Antivenom Neutralizes the Am and Bm Venom Effects on Potassium Channels
2.5. Am and Bm Venoms Do Not Modify Normal Functioning of the Acetylcholine Nicotinic Receptor
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Solutions
4.3. Source of Venom and Antivenom
4.4. Records
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chippaux, J.P.; Goyffon, M. Epidemiology of scorpionism: A global appraisal. Acta Trop. 2008, 107, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Prendini, L.; Order Scorpiones, C.L. Koch, 1850. In Animal Biodiversity: An Outline of Higher-Level Classification and Survey of Taxonomic Richness; Zhang, Z.Q., Ed.; Zootaxa: Auckland, New Zealand, 2011; Volume 3148, pp. 115–177. [Google Scholar]
- Rein-Ove, J. Scorpion Files, Tronheim. Norwegian University of Science and Technology, 2014. Available online: http://www.ntnu.no/ub/scorpion-files (accessed on 1 October 2015).
- Miranda, F.; Kupeyan, C.; Rochat, H.; Rochat, C.; Lissitzky, S. Purification of animal neurotoxins. Isolation and characterization of eleven neurotoxins from the venoms of the scorpions androctonus australis hector, buthus occitanus tunetanus and leiurus quinquestriatus quinquestriatus. Eur. J. Biochem. 1970, 16, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Possani, L.D.; Becerril, B.; Delepierre, M.; Tytgat, J. Scorpion toxins specific for Na+-channels. Eur. J. Biochem. 1999, 264, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Zlotkin, E.; Miranda, F.; Rochat, H. Chemistry and pharmacology of Buthidae scorpion venoms. Arthropod Venom. In Handbook of Experimental Pharmacology; Bettini, S., Ed.; Springer: Verlag/Berlin, Germany, 1978; Volume 48, pp. 317–369. [Google Scholar]
- Dehesa-Davila, M.; Possani, L.D. Scorpionism and serotherapy in mexico. Toxicon 1994, 32, 1015–1018. [Google Scholar] [CrossRef]
- Chippaux, J.P.; Alagon, A. Envenomation and poisoning by venomous or poisonous animals. Vii: Arachnidism in the new world. Med. Trop. (Mars) 2008, 68, 215–221. [Google Scholar] [PubMed]
- Espino-Solis, G.P.; Riano-Umbarila, L.; Becerril, B.; Possani, L.D. Antidotes against venomous animals: State of the art and prospectives. J. Proteom. 2009, 72, 183–199. [Google Scholar] [CrossRef] [PubMed]
- Restano-Cassulini, R.; Korolkova, Y.V.; Diochot, S.; Gurrola, G.; Guasti, L.; Possani, L.D.; Lazdunski, M.; Grishin, E.V.; Arcangeli, A.; Wanke, E. Species diversity and peptide toxins blocking selectivity of Ether-à-go-go-related gene subfamily k+ channels in the central nervous system. Mol. Pharmacol. 2006, 69, 1673–1683. [Google Scholar] [CrossRef] [PubMed]
- Riano-Umbarila, L.; Ledezma-Candanoza, L.M.; Serrano-Posada, H.; Fernandez-Taboada, G.; Olamendi-Portugal, T.; Rojas-Trejo, S.; Gomez-Ramirez, I.V.; Rudino-Pinera, E.; Possani, L.D.; Becerril, B. Optimal neutralization of centruroides noxius venom is understood through a structural complex between two antibody fragments and the cn2 toxin. J. Biol. Chem. 2016, 291, 1619–1630. [Google Scholar] [CrossRef] [PubMed]
- Bosmans, F.; Tytgat, J. Voltage-gated sodium channel modulation by scorpion alpha-toxins. Toxicon 2007, 49, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Cestele, S.; Qu, Y.; Rogers, J.C.; Rochat, H.; Scheuer, T.; Catterall, W.A. Voltage sensor-trapping: Enhanced activation of sodium channels by beta-scorpion toxin bound to the s3-s4 loop in domain ii. Neuron 1998, 21, 919–931. [Google Scholar] [CrossRef]
- Schiavon, E.; Pedraza-Escalona, M.; Gurrola, G.B.; Olamendi-Portugal, T.; Corzo, G.; Wanke, E.; Possani, L.D. Negative-shift activation, current reduction and resurgent currents induced by beta-toxins from centruroides scorpions in sodium channels. Toxicon 2012, 59, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, H.; Chavez-Haro, A.; Garcia-Ubbelohde, W.; Mancilla-Nava, R.; Paniagua-Solis, J.; Alagon, A.; Sevcik, C. Pharmacokinetics of a f(ab’)2 scorpion antivenom in healthy human volunteers. Toxicon 2005, 46, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Boyer, L.V.; Theodorou, A.A.; Chase, P.B.; Osnaya, N.; Berg, M.; Mallie, J.; Carbajal, Y.; de Jesus-Hernandez, T.; Olvera, F.; Alagon, A. Effectiveness of centruroides scorpion antivenom compared to historical controls. Toxicon 2013, 76, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Oukkache, N.; Ahmad Rusmili, M.R.; Othman, I.; Ghalim, N.; Chgoury, F.; Boussadda, L.; Elmdaghri, N.; Sabatier, J.M. Comparison of the neurotoxic and myotoxic effects of two moroccan scorpion venoms and their neutralization by experimental polyclonal antivenom. Life Sci. 2015, 124, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.S.; Silva, C.G.; Neto, B.S.; Grangeiro Junior, C.R.; Lopes, V.H.; Teixeira Junior, A.G.; Bezerra, D.A.; Luna, J.V.; Cordeiro, J.B.; Junior, J.G.; et al. Clinical and epidemiological aspects of scorpionism in the world: A systematic review. Wilderness Environ. Med. 2016, 27, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A.; Cestele, S.; Yarov-Yarovoy, V.; Yu, F.H.; Konoki, K.; Scheuer, T. Voltage-gated ion channels and gating modifier toxins. Toxicon 2007, 49, 124–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintero-Hernandez, V.; Jimenez-Vargas, J.M.; Gurrola, G.B.; Valdivia, H.H.; Possani, L.D. Scorpion venom components that affect ion-channels function. Toxicon 2013, 76, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Escalona, M.P.; Batista, C.V.; Cassulini, R.R.; Rios, M.S.; Coronas, F.I.; Possani, L.D. A proteomic analysis of the early secondary molecular effects caused by cn2 scorpion toxin on neuroblastoma cells. J. Proteom. 2014, 111, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Housley, D.M.; Housley, G.D.; Liddell, M.J.; Jennings, E.A. Scorpion toxin peptide action at the ion channel subunit level. Neuropharmacology 2016. [Google Scholar] [CrossRef] [PubMed]
- Stratton, M.R.; Darling, J.; Pilkington, G.J.; Lantos, P.L.; Reeves, B.R.; Cooper, C.S. Characterization of the human cell line te671. Carcinogenesis 1989, 10, 899–905. [Google Scholar] [CrossRef] [PubMed]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Restano-Cassulini, R.; Garcia, W.; Paniagua-Solís, J.F.; Possani, L.D. Antivenom Evaluation by Electrophysiological Analysis. Toxins 2017, 9, 74. https://doi.org/10.3390/toxins9030074
Restano-Cassulini R, Garcia W, Paniagua-Solís JF, Possani LD. Antivenom Evaluation by Electrophysiological Analysis. Toxins. 2017; 9(3):74. https://doi.org/10.3390/toxins9030074
Chicago/Turabian StyleRestano-Cassulini, Rita, Walter Garcia, Jorge F. Paniagua-Solís, and Lourival D. Possani. 2017. "Antivenom Evaluation by Electrophysiological Analysis" Toxins 9, no. 3: 74. https://doi.org/10.3390/toxins9030074
APA StyleRestano-Cassulini, R., Garcia, W., Paniagua-Solís, J. F., & Possani, L. D. (2017). Antivenom Evaluation by Electrophysiological Analysis. Toxins, 9(3), 74. https://doi.org/10.3390/toxins9030074