
Characterization of Enterotoxigenic Bacillus cereus sensu lato and Staphylococcus aureus Isolates and Associated Enterotoxin Production Dynamics in Milk or Meat-Based Broth

Abstract
:1. Introduction
2. Results
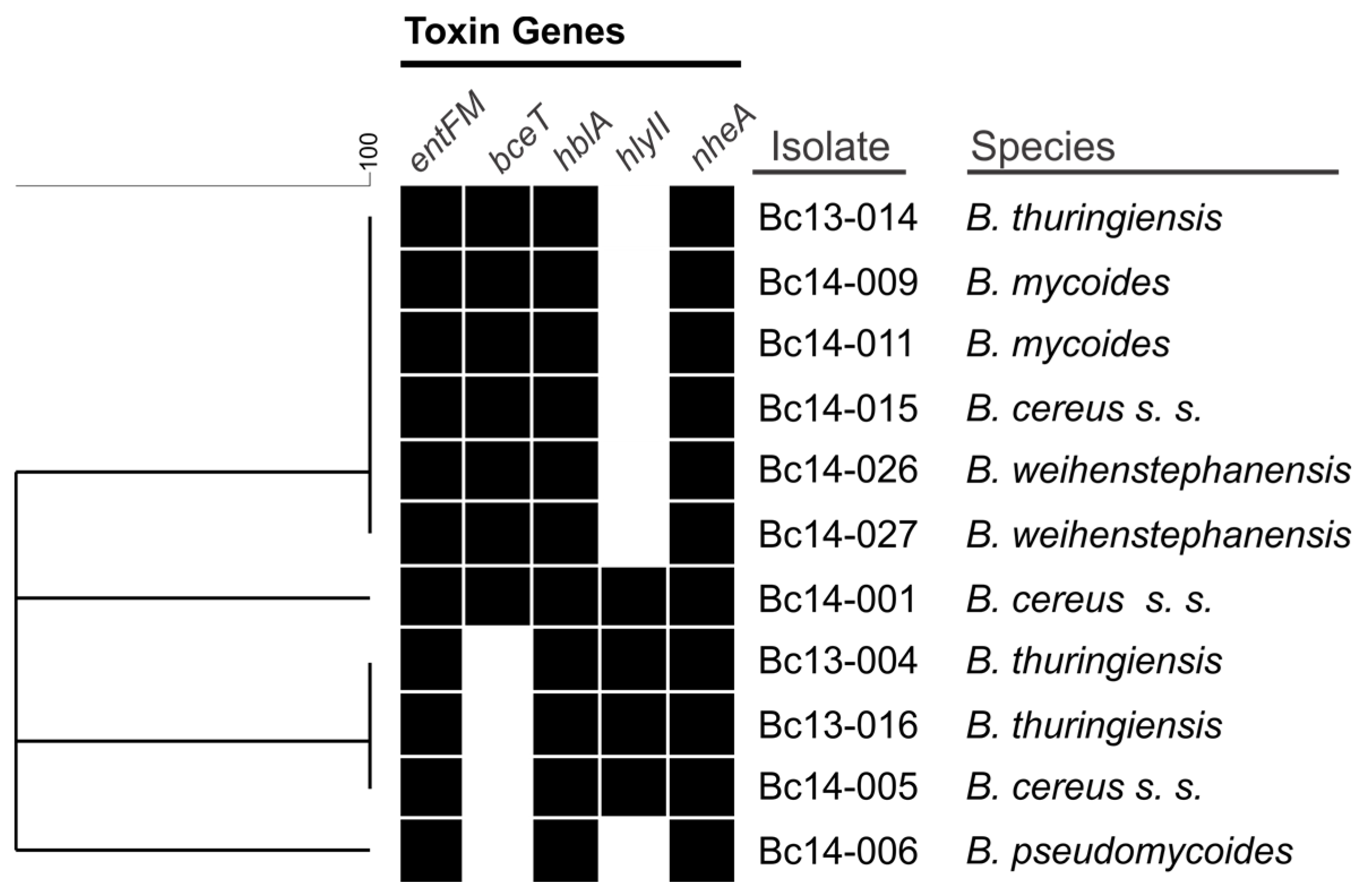
2.1. Toxin Gene Screening of B. Cereus s. l. Isolates
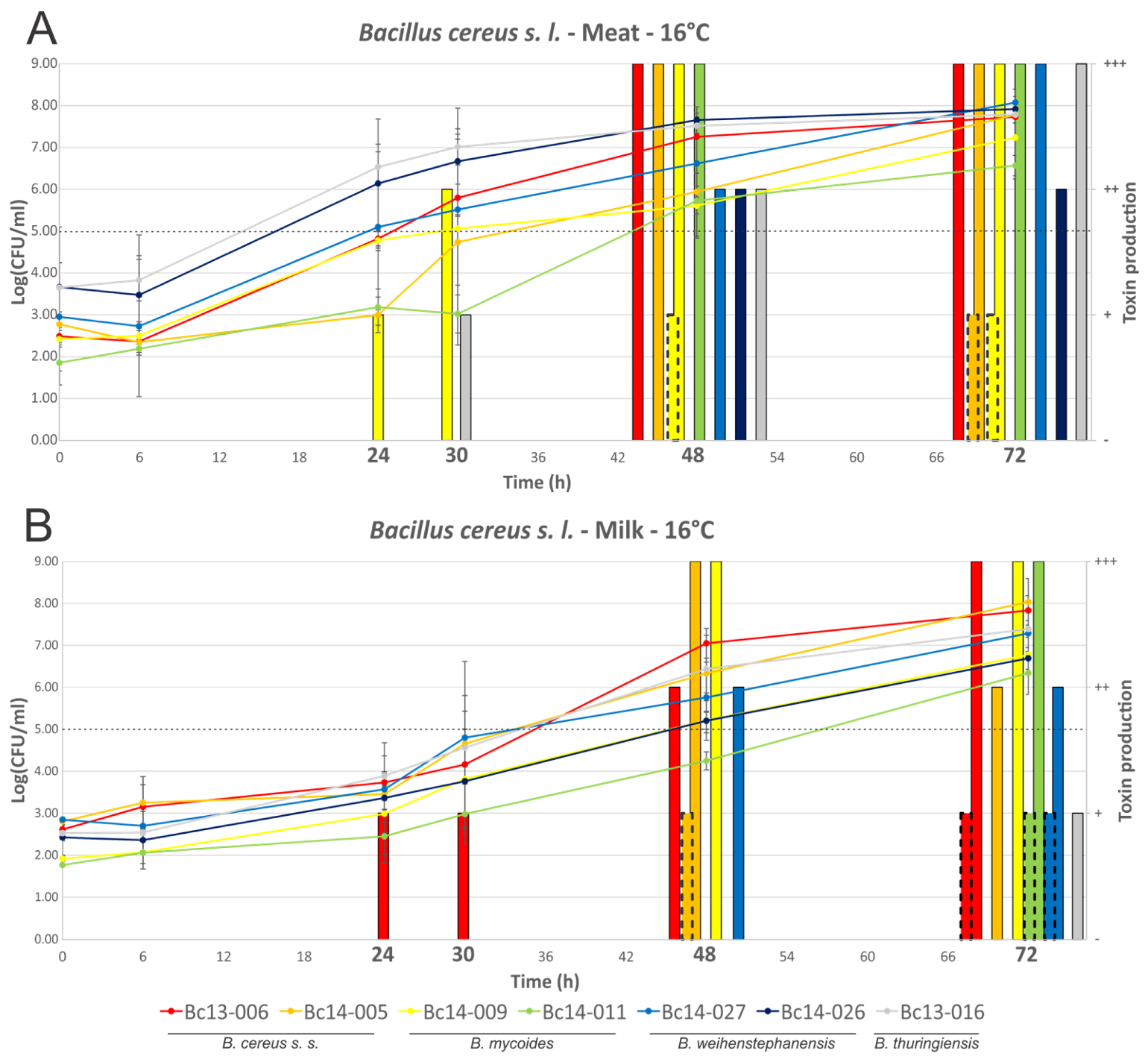
2.2. Growth of B. cereus s. l. and Production of NHE and HBL in MBB or Milk Medium
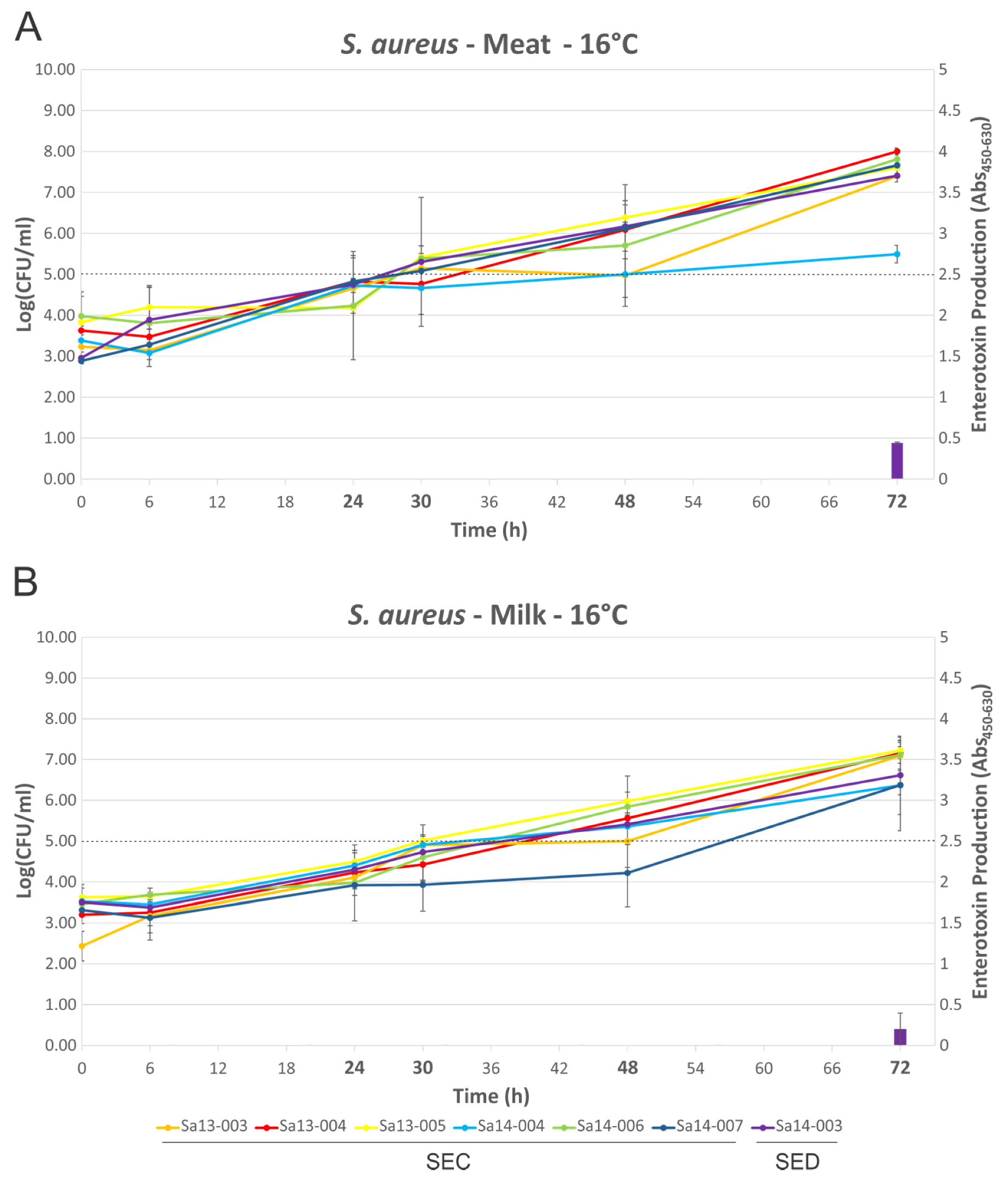
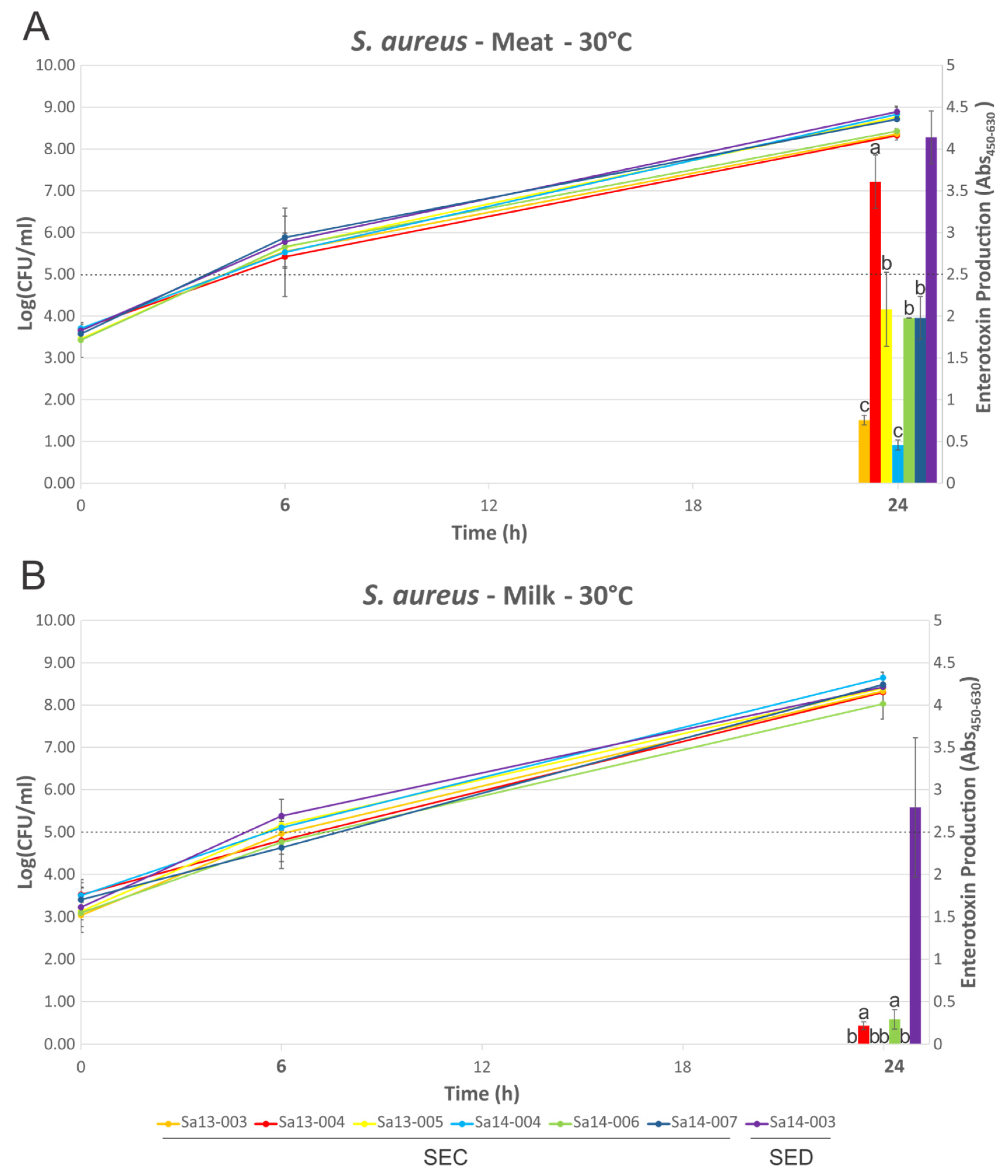
2.3. Growth of S. aureus and Production of SEC or SED Toxin in MBB or Milk Medium
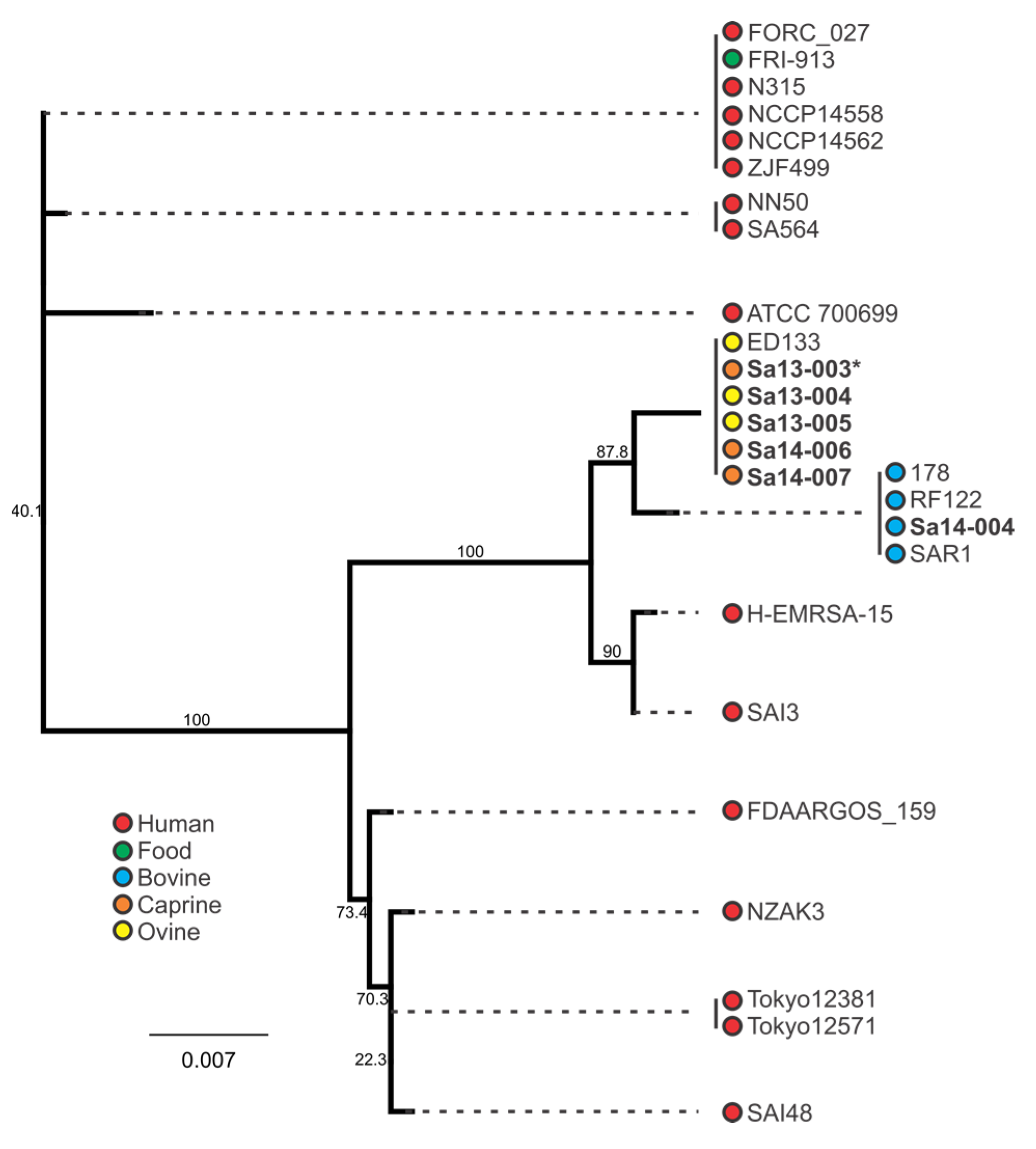
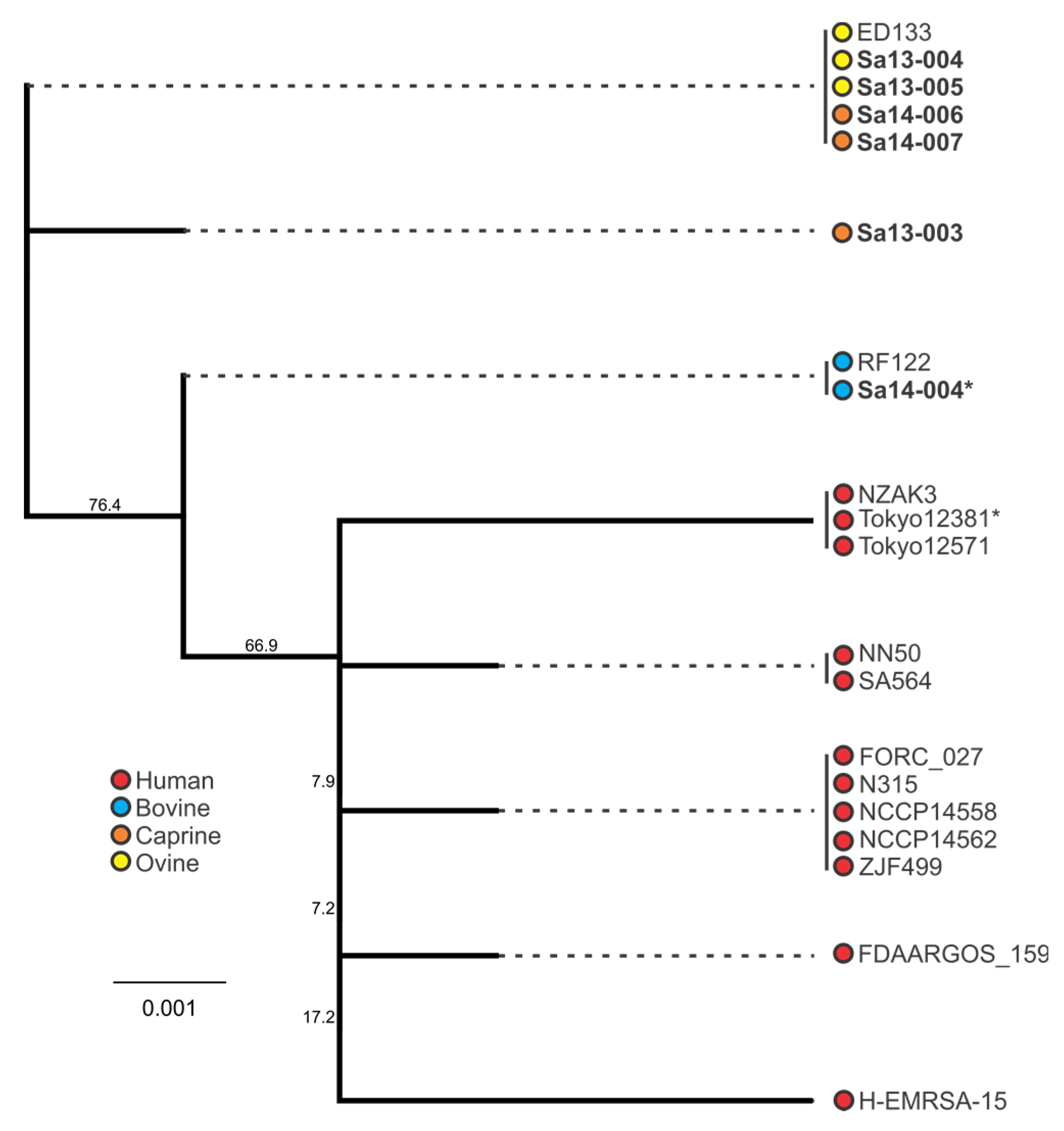
2.4. Phylogenetic Analysis of sec and sel Toxin Genes
3. Discussion
3.1. Toxin Gene Carriage among B. cereus s. l. Isolates in this Study
3.2. Behaviour of B. cereus s. l. in Food Matrices
3.3. Behaviour of S. aureus in Food Matrices
3.4. Phylogenetic Analysis of S. aureus Enterotoxin Genes sec and sel
4. Conclusions
5. Materials and Methods
5.1. Isolates Included in This Study
5.2. Preparation of Milk and MBB
5.3. Enterotoxin Gene Detection
5.4. Growth Curves
5.5. Enterotoxin Detection
5.6. Phylogenetic Analysis of Toxin Genes
5.7. Statistical Analyses
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- WHO. WHO Estimates of the Global Burden of Foodborne Diseases; World Health Organization: Geneva, Switzerland, 2015; p. 268. [Google Scholar]
- Kirk, M.; Ford, L.; Glass, K.; Hall, G. Foodborne illness, Australia, circa 2000 and circa 2010. Emerg. Infect. Dis. 2014, 20, 1857–1864. [Google Scholar] [CrossRef] [PubMed]
- Kirk, M.D.; McKay, I.; Hall, G.V.; Dalton, C.B.; Stafford, R.; Unicomb, L.; Gregory, J. Food safety: Foodborne disease in Australia: The OzFoodNet experience. Clin. Infect. Dis. 2008, 47, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Buzby, J.C.; Roberts, T. The Economics of Enteric Infections: Human Foodborne Disease Costs. Gastroenterology 2009, 136, 1851–1862. [Google Scholar] [CrossRef] [PubMed]
- Flint, J.A.; Van Duynhoven, Y.T.; Angulo, F.J.; DeLong, S.M.; Braun, P.; Kirk, M.; Scallan, E.; Fitzgerald, M.; Adak, G.K.; Sockett, P.; et al. Estimating the Burden of Acute Gastroenteritis, Foodborne Disease, and Pathogens Commonly Transmitted by Food: An International Review. Clin. Infect. Dis. 2005, 41, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Delbrassinne, L.; Botteldoorn, N.; Andjelkovic, M.; Dierick, K.; Denayer, S. An Emetic Bacillus cereus Outbreak in a Kindergarten: Detection and Quantification of Critical Levels of Cereulide Toxin. Foodborne Pathog. Dis. 2014, 12, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Fetsch, A.; Contzen, M.; Hartelt, K.; Kleiser, A.; Maassen, S.; Rau, J.; Kraushaar, B.; Layer, F.; Strommenger, B. Staphylococcus aureus food-poisoning outbreak associated with the consumption of ice-cream. Int. J. Food Microbiol. 2014, 187, 1–6. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, L.; Bernard, K.; Beniac, D.; Isaac-Renton, J.L.; Naseby, D.C. Identification of Bacillus cereus Group Species Associated with Food Poisoning Outbreaks in British Columbia, Canada. Appl. Environ. Microbiol. 2008, 74, 7451–7453. [Google Scholar] [CrossRef] [PubMed]
- Guinebretière, M.-H.; Auger, S.; Galleron, N.; Contzen, M.; De Sarrau, B.; De Buyser, M.-L.; Lamberet, G.; Fagerlund, A.; Granum, P.E.; Lereclus, D.; et al. Bacillus cytotoxicus sp. nov. is a novel thermotolerant species of the Bacillus cereus Group occasionally associated with food poisoning. Int. J. Syst. Evolut. Microbiol. 2013, 63, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Guinebretière, M.-H.; Thompson, F.L.; Sorokin, A.; Normand, P.; Dawyndt, P.; Ehling-Schulz, M.; Svensson, B.; Sanchis, V.; Nguyen-The, C.; Heyndrickx, M.; et al. Ecological diversification in the Bacillus cereus Group. Environ. Microbiol. 2008, 10, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lai, Q.; Göker, M.; Meier-Kolthoff, J.P.; Wang, M.; Sun, Y.; Wang, L.; Shao, Z. Genomic insights into the taxonomic status of the Bacillus cereus group. Sci. Rep. 2015, 5, 14082. [Google Scholar] [CrossRef] [PubMed]
- Beecher, D.J.; MacMillan, J.D. A novel bicomponent hemolysin from Bacillus cereus. Infect. Immun. 1990, 58, 2220–2227. [Google Scholar] [PubMed]
- Lund, T.; De Buyser, M.-L.; Granum, P.E. A new cytotoxin from Bacillus cereus that may cause necrotic enteritis. Mol. Microbiol. 2000, 38, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Lund, T.; Granum, P.E. Characterisation of a non-haemolytic enterotoxin complex from Bacillus cereus isolated after a foodborne outbreak. FEMS Microbiol. Lett. 1996, 141, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Agata, N.; Mori, M.; Ohta, M.; Suwan, S.; Ohtani, I.; Isobe, M. A novel dodecadepsipeptide, cereulide, isolated from Bacillus cereus causes vacuole formation in HEp-2 cells. FEMS Microbiol. Lett. 1994, 121, 31–34. [Google Scholar] [PubMed]
- Agata, N.; Ohta, M.; Arakawa, Y.; Mori, M. The bceT gene of Bacillus cereus encodes an enterotoxic protein. Microbiology 1995, 141, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Miles, G.; Bayley, H.; Cheley, S. Properties of Bacillus cereus hemolysin II: A heptameric transmembrane pore. Protein Sci. 2002, 11, 1813–1824. [Google Scholar] [CrossRef] [PubMed]
- Asano, S.I.; Nukumizu, Y.; Bando, H.; Iizuka, T.; Yamamoto, T. Cloning of novel enterotoxin genes from Bacillus cereus and Bacillus thuringiensis. Appl. Environ. Microbiol. 1997, 63, 1054–1057. [Google Scholar] [PubMed]
- Argudín, M.Á.; Mendoza, M.C.; Rodicio, M.R. Food Poisoning and Staphylococcus aureus Enterotoxins. Toxins 2010, 2, 1751–1773. [Google Scholar] [CrossRef] [PubMed]
- Ono, H.K.; Sato’o, Y.; Narita, K.; Naito, I.; Hirose, S.; Hisatsune, J.; Asano, K.; Hu, D.-L.; Omoe, K.; Sugai, M.; et al. Identification and Characterization of a Novel Staphylococcal Emetic Toxin. Appl. Environ. Microbiol. 2015, 81, 7034–7040. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, H.J.; Mathisen, T.; Løvseth, A.; Omoe, K.; Qvale, K.S.; Loncarevic, S. An outbreak of staphylococcal food poisoning caused by enterotoxin H in mashed potato made with raw milk. FEMS Microbiol. Lett. 2005, 252, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Hall, G.; Kirk, M.D.; Becker, N.; Gregory, J.E.; Unicomb, L.; Millard, G.; Stafford, R.; Lalor, K.; OzFoodNet Working Group. Estimating Foodborne Gastroenteritis, Australia. Emerg. Infect. Dis. 2005, 11, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Valihrach, L.; Alibayov, B.; Zdenkova, K.; Demnerova, K. Expression and production of staphylococcal enterotoxin C is substantially reduced in milk. Food Microbiol. 2014, 44, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Hunt, K.; Butler, F.; Jordan, K. Factors affecting staphylococcal enterotoxin Cbovine production in milk. Int. Dairy J. 2014, 39, 41–46. [Google Scholar] [CrossRef]
- Otero, A.; García, M.C.; García, M.L.; Moreno, B. Production of staphylococcal enterotoxins C1 and C2 and thermonuclease in ewe’s milk. Food Microbiol. 1987, 4, 339–345. [Google Scholar] [CrossRef]
- Schubert, J.; Podkowik, M.; Bystroń, J.; Bania, J. Production of Staphylococcal Enterotoxins D and R in Milk and Meat Juice by Staphylococcus aureus Strains. Foodborne Pathog. Dis. 2017, 14, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Dréan, P.; McAuley, C.M.; Moore, S.C.; Fegan, N.; Fox, E.M. Characterization of the spore-forming Bacillus cereus sensu lato group and Clostridium perfringens bacteria isolated from the Australian dairy farm environment. BMC Microbiol. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, H.; Morozumi, S. Modeling Staphylococcus aureus growth and enterotoxin production in milk. Food Microbiol. 2006, 23, 260–267. [Google Scholar] [CrossRef] [PubMed]
- European Union. Commission Regulation (EC) No. 2073/2005 on Microbiological Criteria for Foodstuffs; European Union: Brussels, Belgium, 2005; p. 26. [Google Scholar]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [PubMed]
- Otero, A.; García, M.L.; García, M.C.; Moreno, B.; Bergdoll, M.S. Production of staphylococcal enterotoxins C1 and C2 and thermonuclease throughout the growth cycle. Appl. Environ. Microbiol. 1990, 56, 555–559. [Google Scholar] [PubMed]
- McMillan, K.; Moore, S.C.; McAuley, C.M.; Fegan, N.; Fox, E.M. Characterization of Staphylococcus aureus isolates from raw milk sources in Victoria, Australia. BMC Microbiol. 2016, 16, 169. [Google Scholar] [CrossRef] [PubMed]
- Guinebretière, M.-H.; Broussolle, V.; Nguyen-The, C. Enterotoxigenic Profiles of Food-Poisoning and Food-Borne Bacillus cereus Strains. J. Clin. Microbiol. 2002, 40, 3053–3056. [Google Scholar] [CrossRef] [PubMed]
- Moravek, M.; Dietrich, R.; Buerk, C.; Broussolle, V.; Guinebretière, M.-H.; Granum, P.E.; Nguyen-the, C.; Märtlbauer, E. Determination of the toxic potential of Bacillus cereus isolates by quantitative enterotoxin analyses. FEMS Microbiol. Lett. 2006, 257, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Fedhila, S.; Gohar, M.; Slamti, L.; Nel, P.; Lereclus, D. The Bacillus thuringiensis PlcR-Regulated Gene inhA2 Is Necessary, but Not Sufficient, for Virulence. J. Bacteriol. 2003, 185, 2820–2825. [Google Scholar] [CrossRef] [PubMed]
- Andreeva, Z.I.; Nesterenko, V.F.; Yurkov, I.S.; Budarina, Z.I.; Sineva, E.V.; Solonin, A.S. Purification and cytotoxic properties of Bacillus cereus hemolysin II. Protein Expr. Purif. 2006, 47, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Harvie, D.R.; Vilchez, S.; Steggles, J.R.; Ellar, D.J. Bacillus cereus Fur regulates iron metabolism and is required for full virulence. Microbiology 2005, 151, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Castiaux, V.; Laloux, L.; Schneider, Y.-J.; Mahillon, J. Screening of Cytotoxic B. cereus on Differentiated Caco-2 Cells and in Co-Culture with Mucus-Secreting (HT29-MTX) Cells. Toxins 2016, 8, 320. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Kwarteng, J.; Wuni, A.; Akabanda, F.; Tano-Debrah, K.; Jespersen, L. Prevalence, virulence factor genes and antibiotic resistance of Bacillus cereus sensu lato isolated from dairy farms and traditional dairy products. BMC Microbiol. 2017, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.A.; Almeida, F.S.; Guimarães, A.G.; Abrahão, W.M.; Arantes, O.M.N.; Vilas-Bôas, G.T. RE-PCR variability and toxigenic profile of food poisoning, foodborne and soil-associated Bacillus cereus isolates from Brazil. Int. J. Food Microbiol. 2011, 151, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Han, J.K.; Park, J.S.; Lee, J.S.; Lee, S.H.; Cho, J.I.; Kim, K.S. Various Enterotoxin and Other Virulence Factor Genes Widespread Among Bacillus cereus and Bacillus thuringiensis Strains. J. Microbiol. Biotechnol. 2015, 25, 872–879. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Opinion of the Scientific Panel on biological hazards (BIOHAZ) on Bacillus cereus and other Bacillus spp. in foodstuffs. EFSA J. 2005, 3, 175. [Google Scholar] [CrossRef]
- Ceuppens, S.; Boon, N.; Uyttendaele, M. Diversity of Bacillus cereus group strains is reflected in their broad range of pathogenicity and diverse ecological lifestyles. FEMS Microbiol. Ecol. 2013, 84, 433–450. [Google Scholar] [CrossRef] [PubMed]
- Wijnands, L.; Dufrenne, J.; van Leusden, F. Bacillus Cereus: Characteristics, Behaviour in the Gastro-Intestinal Tract, and Interaction with Caco-2 Cells; National Institute for Public Health and the Environment “Rijksinstituut voor Volksgezondheid en Milieu”: Bilthoven, The Netherlands, 2005.
- Waterman, S.R.; Small, P.L.C. Acid-Sensitive Enteric Pathogens Are Protected from Killing under Extremely Acidic Conditions of pH 2.5 when They Are Inoculated onto Certain Solid Food Sources. Appl. Environ. Microbiol. 1998, 64, 3882–3886. [Google Scholar] [PubMed]
- Takumi, K.; De Jonge, R.; Havelaar, A. Modelling inactivation of Escherichia coli by low pH: Application to passage through the stomach of young and elderly people. J. Appl. Microbiol. 2000, 89, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Dressman, J.B.; Berardi, R.R.; Dermentzoglou, L.C.; Russell, T.L.; Schmaltz, S.P.; Barnett, J.L.; Jarvenpaa, K.M. Upper Gastrointestinal (GI) pH in Young, Healthy Men and Women. Pharm. Res. 1990, 7, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Ceuppens, S.; Rajkovic, A.; Hamelink, S.; Van de Wiele, T.; Boon, N.; Uyttendaele, M. Enterotoxin production by Bacillus cereus under gastrointestinal conditions and their immunological detection by commercially available kits. Foodborne Pathog. Dis. 2012, 9, 1130–1136. [Google Scholar] [CrossRef] [PubMed]
- Sams, L.; Paume, J.; Giallo, J.; Carriere, F. Relevant pH and lipase for in vitro models of gastric digestion. Food Funct. 2016, 7, 30–45. [Google Scholar] [CrossRef] [PubMed]
- Clavel, T.; Carlin, F.; Lairon, D.; Nguyen-The, C.; Schmitt, P. Survival of Bacillus cereus spores and vegetative cells in acid media simulating human stomach. J. Appl. Microbiol. 2004, 97, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Berthold-Pluta, A.; Pluta, A.; Garbowska, M. The effect of selected factors on the survival of Bacillus cereus in the human gastrointestinal tract. Microb. Pathog. 2015, 82, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Orsi, R.H.; Bowen, B.M.; Wiedmann, M. Homopolymeric tracts represent a general regulatory mechanism in prokaryotes. BMC Genom. 2010, 11, 102. [Google Scholar] [CrossRef] [PubMed]
- Lowder, B.V.; Guinane, C.M.; Ben Zakour, N.L.; Weinert, L.A.; Conway-Morris, A.; Cartwright, R.A.; Simpson, A.J.; Rambaut, A.; Nübel, U.; Fitzgerald, J.R. Recent human-to-poultry host jump, adaptation, and pandemic spread of Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2009, 106, 19545–19550. [Google Scholar] [CrossRef] [PubMed]
- Sakwinska, O.; Giddey, M.; Moreillon, M.; Morisset, D.; Waldvogel, A.; Moreillon, P. Staphylococcus aureus Host Range and Human-Bovine Host Shift. Appl. Environ. Microbiol. 2011, 77, 5908–5915. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.M.; Needs, P.F.; Manley, G.; Green, L.E. Global distribution and diversity of ovine-associated Staphylococcus aureus. Infect. Gen. Evolut. 2014, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- De Souza, E.L.; Meira, Q.G.S.; de Medeiros Barbosa, I.; Athayde, A.J.A.A.; da Conceição, M.L.; de Siqueira Júnior, J.P. Biofilm formation by Staphylococcus aureus from food contact surfaces in a meat-based broth and sensitivity to sanitizers. Braz. J. Microbiol. 2014, 45, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Ween, O.; Lund, T.; Hardy, S.P.; Granum, P.E. Genetic and functional analysis of the cytK family of genes in Bacillus cereus. Microbiology 2004, 150, 2689–2697. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evolut. 1985, 22, 160–174. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | Protein | Fat | Carbohydrate | Moisture | Ash | pH |
|---|---|---|---|---|---|---|
| 10% RSM | 3.4% | 0.15% | ND * | 90% | 0.8% | 6.4 |
| Meat-based broth | 1.1% | 0.1% | <0.1% | 98.5% | 0.4% | 5.94 |
| Primer Name | Gene Target | Annealing Temperature | Product Size | Reference |
|---|---|---|---|---|
| hblA-F | hblA | GTGGTGGATTGGGAGCAG | 390 bp | This study |
| hblA-R | CTTGCATAGARTCGATATTATC | |||
| nheA-F | nheA | TACAGGGTTATTGGTTACAGC | 482 bp | This study |
| nheA-R | CACAATATCTCCACTTGATCCT | |||
| bceT-F | bceT | GCTACGCAAAAACCGAGTGGTG | 679 bp | Kim et al. (2015) |
| bceT-R | AATGCTCCGGACTATGCTGACG | |||
| ENTA | entFM | ATGAAAAAAGTAATTTGCAGG | 1269 bp | Asano et al. (1997) |
| ENTB | TTAGTATGCTTTTGTGTAACC | |||
| Fhly-II | hlyII | GATTCTAAAGGMACTGTAG | 868 bp | Adapted from Fagerlund et al. (2004) |
| Rhly-II | GGTTATCAAGAGTAACTTG |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker-York-Moore, L.; Moore, S.C.; Fox, E.M. Characterization of Enterotoxigenic Bacillus cereus sensu lato and Staphylococcus aureus Isolates and Associated Enterotoxin Production Dynamics in Milk or Meat-Based Broth. Toxins 2017, 9, 225. https://doi.org/10.3390/toxins9070225
Walker-York-Moore L, Moore SC, Fox EM. Characterization of Enterotoxigenic Bacillus cereus sensu lato and Staphylococcus aureus Isolates and Associated Enterotoxin Production Dynamics in Milk or Meat-Based Broth. Toxins. 2017; 9(7):225. https://doi.org/10.3390/toxins9070225
Chicago/Turabian StyleWalker-York-Moore, Laura, Sean C. Moore, and Edward M. Fox. 2017. "Characterization of Enterotoxigenic Bacillus cereus sensu lato and Staphylococcus aureus Isolates and Associated Enterotoxin Production Dynamics in Milk or Meat-Based Broth" Toxins 9, no. 7: 225. https://doi.org/10.3390/toxins9070225
APA StyleWalker-York-Moore, L., Moore, S. C., & Fox, E. M. (2017). Characterization of Enterotoxigenic Bacillus cereus sensu lato and Staphylococcus aureus Isolates and Associated Enterotoxin Production Dynamics in Milk or Meat-Based Broth. Toxins, 9(7), 225. https://doi.org/10.3390/toxins9070225