
SiMa Cells for a Serotype Specific and Sensitive Cell-Based Neutralization Test for Botulinum Toxin A and E

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Treatment of SiMa Cells and Sensitivity to BoNT/A and BoNT/E
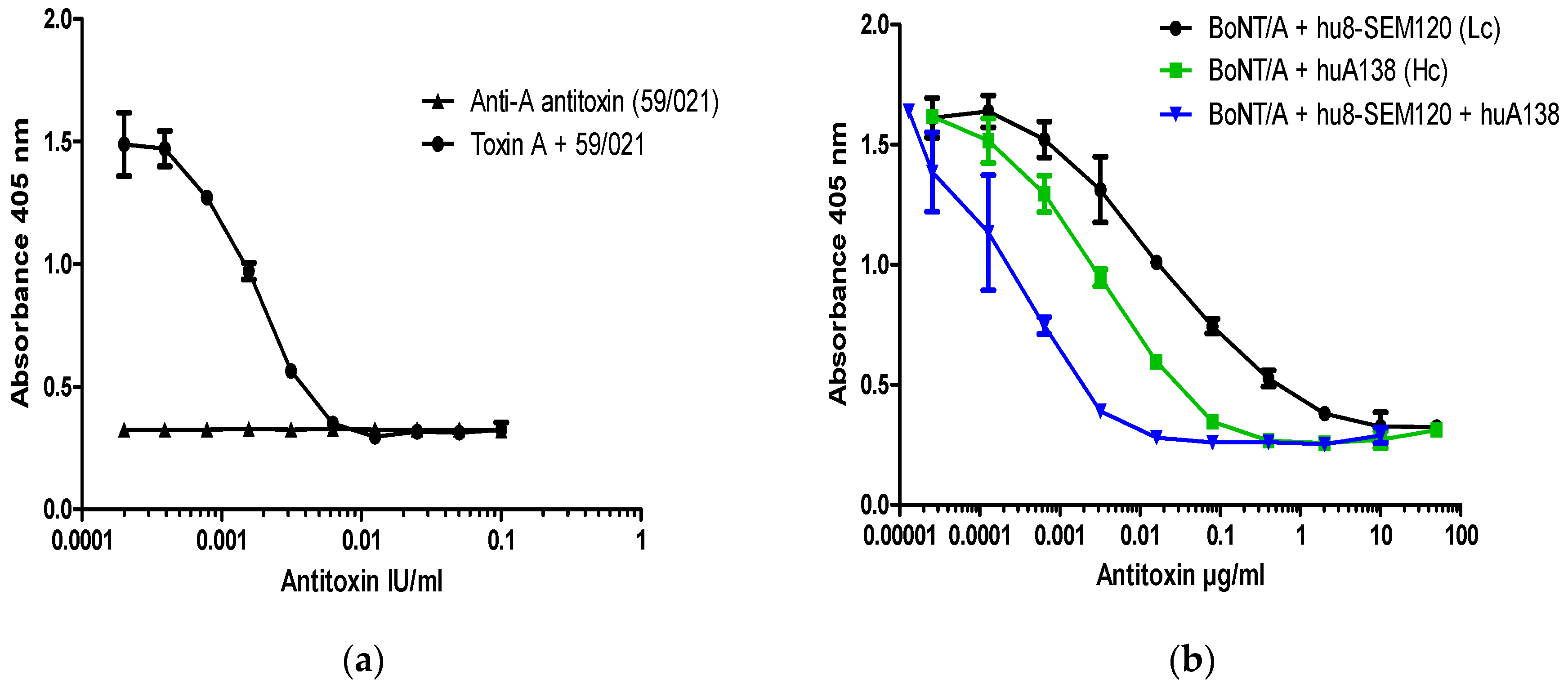
2.2. BoNT/A Neutralization Assay
2.3. BoNT/E Neutralization Assay
3. Discussion
4. Materials and Methods
4.1. Toxins
4.2. Antibodies
4.3. Preparation of SiMa Cells and Neutralization Assays
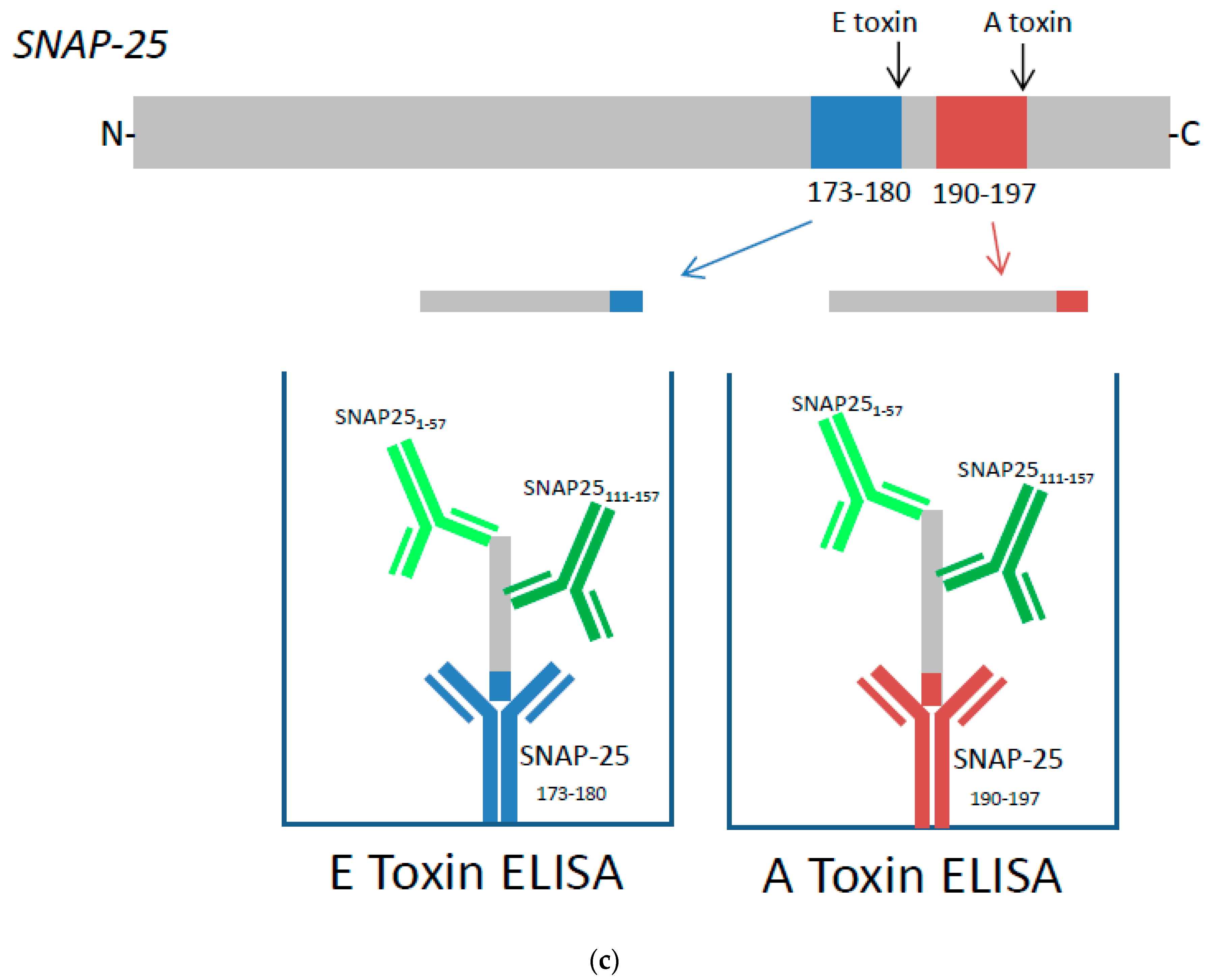
4.4. Preparation of Cell Lysate and Capture ELISA
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Smith, T.J.; Hill, K.K.; Raphael, B.H. Historical and current perspectives on Clostridium botulinum diversity. Res. Microbiol. 2015, 166, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Microbiol. 2014, 12, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Gill, D.M. Bacterial toxins: A table of lethal amounts. Micobiol. Rev. 1982, 46, 86–94. [Google Scholar]
- Pirazzini, M.; Azarnia Tehran, D.; Zanetti, G.; Megighian, A.; Scorzeto, M.; Fillo, S.; Shone, C.C.; Binz, T.; Rossetto, O.; Lista, F.; et al. Thioredoxin and its reductase are present on synaptic vesicles, and their inhibition prevents the paralysis induced by botulinum neurotoxins. Cell Rep. 2014, 8, 1870–1878. [Google Scholar] [CrossRef] [PubMed]
- Azarnia Tehran, D.; Pirazzini, M.; Leka, O.; Mattarei, A.; Lista, F.; Binz, T.; Rossetto, O.; Montecucco, C. Hsp90 is involved in the entry of clostridial neurotoxins into the cytosol of nerve terminals. Cell Microbiol. 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Pirazzini, M.; Azarnia Tehran, D.; Leka, O.; Zanetti, G.; Rossetto, O.; Montecucco, C. On the translocation of botulinum and tetanus neurotoxins across the membrane of acidic intracellular compartments. Biochim. Biophys. Acta 2016, 1858, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Pirazzini, M.; Rossetto, O.; Eleopra, R.; Montecucco, C. Botulinum neurotoxins: Biology, pharmacology, and toxicology. Pharmacol. Rev. 2017, 69, 200–235. [Google Scholar] [CrossRef] [PubMed]
- Mayers, C.N.; Holley, J.L.; Brooks, T. Antitoxin therapy for botulinum intoxication. Rev. Med. Microbiol. 2001, 12, 29–37. [Google Scholar] [CrossRef]
- Arnon, S.S.; Schechter, R.; Maslanka, S.E.; Jewell, N.P.; Hatheway, C.L. Human botulism immune globulin for the treatment of infant botulism. N. Engl. J. Med. 2006, 354, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Adekar, S.P.; Takahashi, T.; Jones, R.M.; Al-Saleem, F.H.; Ancharski, D.M.; Root, M.J.; Kapadnis, B.P.; Simpson, L.L.; Dessain, S.K. Neutralization of botulinum neurotoxin by a human monoclonal antibody specific for the catalytic light chain. PLoS ONE 2008, 3, e3023. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhou, B.; Pellett, S.; Johnson, E.A.; Janda, K.D. Selection and characterisation of a human monoclonal neutralizing antibody for Clostridium botulinum neurotoxin Serotype B. Bioorg. Med. Chem. Lett. 2009, 19, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Miethe, S.; Mazuet, C.; Liu, Y.; Tierney, R.; Rasetti-Escargueil, C.; Avril, A.; Frenzel, A.; Thullier, P.; Pelat, T.; Urbain, R.; et al. Development of GermLine-Humanized Antibodies Neutralizing Botulinum Neurotoxin A and B. PLoS ONE 2016, 11, e0161446. [Google Scholar] [CrossRef] [PubMed]
- Derman, Y.; Selby, K.; Miethe, S.; Frenzel, A.; Liu, Y.; Rasetti-Escargueil, C.; Avril, A.; Pelat, T.; Urbain, R.; Fontayne, A.; et al. Neutralization of Botulinum Neurotoxin Type E by a Humanized Antibody. Toxins 2016, 8, 257. [Google Scholar] [CrossRef] [PubMed]
- Sesardic, D. Bioassays for evaluation of medical products derived from bacterial toxins. Curr. Opin. Microbiol. 2012, 15, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.G.; Alsop, T.A.; Hull, R.; Tierney, R.; Holley, J.; Sesardic, D. Botulinum type A toxin neutralisation by specific IgG and its fragments: A comparison of mouse systemic toxicity and local flaccid paralysis assays. Toxicon 2006, 48, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Broide, R.S.; Rubino, J.; Nicholson, G.S.; Ardila, M.C.; Brown, M.S.; Aoki, K.R.; Francis, J. The rat Digit Abduction Score (DAS) assay: A physiological model for assessing botulinum neurotoxin-induced skeletal muscle paralysis. Toxicon 2013, 71, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Escargueil, C.; Liu, Y.; Rigsby, P.; Jones, R.G.A.; Sesardic, D. Phrenic nerve-hemidiaphragm as a sensitive replacement assay for determination of functional botulinum toxin antibodies. Toxicon 2011, 57, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Ochiai, M.; Liu, Y.; Ekong, T.; Sesardic, D. Development of improved SNAP25 endopeptidase immuno-assays for botulinum type A and E toxins. J. Immunol. Methods 2008, 329, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Sesardic, D.; Martin, H.; Tierney, R.; Bigalke, H. An in vitro assay for testing of neutralising antibodies to botulinum toxins. In Progress in the Reduction, Refinement and Replacement of Animal Experimentation; Balls, M., Zeller, A.M.V., Eds.; Elsevier Science: Amsterdam, NY, USA, 2000; pp. 1001–1008. [Google Scholar]
- Sesardic, D.; Jones, R.G.; Leung, T.; Alsop, T.; Tierney, R. Detection of antibodies against botulinum toxins. Mov. Disord. 2004, 19, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Chahboun, S.; Hust, M.; Liu, Y.; Pelat, T.; Helmsing, S.; Jones, R.G.A.; Sesardic, D.; Thullier, P. Isolation of a neutralizing, nanomolar scFv inhibiting the proteolytic activity of the light chain of botulinum toxin A, by single step panning of an immune phage-displayed library of macaque origin. BMC Biotechnol. 2011, 11, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Rasetti-Escargueil, C.; Avril, A.; Chahboun, S.; Tierney, R.; Bak, N.; Miethe, S.; Mazuet, C.; Popoff, M.R.; Thullier, P.; Hust, M.; et al. Development of human-like scFv-Fc antibodies neutralizing Botulinum toxin serotype B. mAbs 2015, 7, 1161–1177. [Google Scholar] [CrossRef] [PubMed]
- Rosen, O.; Ozeri, E.; Barnea, A.; David, A.B.; Zichel, R. Development of an innovative in vitro potency assay for anti-botulinum antitoxins. Toxins 2016, 8, 276. [Google Scholar] [CrossRef] [PubMed]
- Pellett, S. Progress in cell-based assays for botulinum neurotoxin detection. Curr. Top. Microbiol. Immunol. 2013, 364, 257–285. [Google Scholar] [PubMed]
- Marini, P.; MacLeod, R.A.; Treuner, C.; Bruchelt, G.; Böhm, W.; Wolburg, H.; Schweizer, P.; Girgert, R.A. SiMa, a new neuroblastoma cell line combining poor prognostic cytogenetic markers with high adrenergic differentiation. Cancer Genet. Cytogenet. 1999, 112, 161–164. [Google Scholar] [CrossRef]
- Fernandez-Salas, E.; Wang, J.; Molina, Y.; Nelson, J.B.; Jacky, B.P.S.; Aoki, K.R. Botulinum neurotoxin serotype A specific cell-based potency assay to replace the mouse bioassay. PLoS ONE 2012, 7, e49516. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, J.; Jacky, B.P.S.; Hodges, D.D.; Fernandez-Salas, E. Cells Useful for Immuno-Based Botulinum Toxin Serotype A Activity Assays. US Patent 12/722,801, 16 September 2010. [Google Scholar]
- Pellett, S.; Tepp, W.H.; Toth, S.I.; Johnson, E.A. Comparison of the primary rat spinal cord cell (RSC) assay and the mouse bioassay for botulinum neurotoxin type A potency determination. J. Pharmacol. Toxicol. Methods 2010, 61, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Whitemarsh, R.C.; Strathman, M.J.; Chase, L.G.; Stankewicz, C.; Tepp, W.H.; Johnson, E.A.; Pellett, S. Novel application of human neurons derived from induced pluripotent stem cells for highly sensitive botulinum neurotoxin detection. Toxicol. Sci. 2012, 126, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.B.; Rigsby, P.; Sesardic, D.; Marks, J.D.; Jones, R.G.A. A functional dual-coated (FDC) microtiter plate method to replace the botulinum toxin LD50 test. Anal. Biochem. 2012, 425, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Rasetti-Escargueil, C.; Machado, C.B.; Preneta-Blanc, R.; Fleck, R.A.; Sesardic, D. Enhanced sensitivity to botulinum type A neurotoxin of human neuroblasoma SH-SY5Y cells after differentiation into mature neuronal cells. Botulinum J. 2011, 2, 30–48. [Google Scholar] [CrossRef]
- Adler, S.; Bicker, G.; Bigalke, H.; Bishop, C.; Blümel, J.; Dressler, D.; Fitzgerald, J.; Gessler, F.; Heuschen, H.; Kegel, B.; et al. The current scientific and legal status of alternative methods to the LD50 test for botulinum neurotoxin potency testing. The report and recommendations of a ZEBET Expert Meeting. Altern. Lab. Anim. 2010, 38, 315–330. [Google Scholar] [PubMed]
- Yowler, B.C.; Kensinger, R.D.; Schengrund, C.L. Botulinum neurotoxin A activity is dependent upon, the presence of specific gangliosides in neuroblastoma cells expressing synaptotagmin I. J. Biol. Chem. 2002, 277, 32815–32819. [Google Scholar] [CrossRef] [PubMed]
- Pellett, S.; Tepp, W.H.; Johnson, E.A.; Sesardic, D. Assessment of ELISA as endpoint in neuronal cell-based assay for BoNT detection using hiPSC derived neurons. J. Pharmacol. Toxicol. Methods 2017, 88, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, J.; Cuijpers, S.A.G.; Ferrari, E.; Niranjan, D.; Rust, A.; Leese, C.; O’Brien, J.A.; Binz, T.; Davletov, B. Botulinum protease-cleaved SNARE fragments induce cytotoxicity in neuroblastoma cells. J. Neurochem. 2014, 129, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Liu, H.; Ruan, H.; Tepp, W.H.; Stoothoff, W.H.; Brown, R.H.; Johnson, E.A.; Yao, W.D.; Zhang, S.C.; Dong, M. Cytotoxicity of botulinum neurotoxins reveals a direct role of syntaxin 1 and SNAP-25 in neuron survival. Nat. Commun. 2013, 4, 1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngundi, M.N.; Meade, B.D.; Lin, T.L.; Tang, W.J.; Burns, D.L. Comparison of three Anthrax toxin neutralization assay. Clin. Vaccine Immunol. 2010, 17, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Reller, M.E.; Alcabasa, R.C.; Lema, C.A.; Carroll, K.C. Comparison of two rapid assays for Clostridium difficile Common antigen and a C difficile toxin A/B assay with the cell culture neutralization assay. Am. J. Clin. Pathol. 2010, 133, 107–109. [Google Scholar] [CrossRef] [PubMed]
- McClain, M.S.; Cover, T.L. Functional Analysis of Neutralizing Antibodies against Clostridium perfringens Epsilon-Toxin. Infect. Immun. 2007, 75, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Pellett, S.; Tepp, W.H.; Clancy, C.M.; Borodic, G.E.; Johnson, E.A. A neuronal cell-based neurotoxin assay for highly sensitive and specific detection of neutralizing serum antibodies. FEBS Lett. 2007, 581, 4803–4808. [Google Scholar] [CrossRef] [PubMed]
- Hall, Y.H.; Chaddock, J.A.; Moulsdale, H.J.; Kirby, E.R.; Alexander, F.C.; Marks, J.D.; Foster, K.A. Novel application of an in vitro technique to the detection and quantification of botulinum neurotoxin antibodies. J. Immunol. Methods 2004, 288, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Göschel, H.; Wohlfarth, K.; Frevert, J.; Dengler, R.; Bigalke, H. Botulinum a toxin therapy: Neutralizing and nonneutralizing antibodies-therapeutic consequences. Exp. Neurol. 1997, 147, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Mazuet, C.; Dano, J.; Popoff, M.R.; Créminon, C.; Volland, H. Characterization of botulinum neurotoxin type A neutralizing monoclonal antibodies and influence of their half-lives on therapeutic activity. PLoS ONE 2010, 5, e12416. [Google Scholar] [CrossRef] [PubMed]
- Davletov, B.; (Department of Biomedical Sciences, University of Sheffield, Sheffield, UK). Personal communication, 2016.
- AntibotABE Project. Available online: http://www.antibotabe.com/ (accessed on 15 May 2017).
- Ekong, T.A.; Feavers, I.M.; Sesardic, D. Recombinant SNAP-25 is an effective substrate for Clostridium botulinum type A toxin endopeptidase activity in vitro. Microbiology 1997, 143, 3337–3347. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bak, N.; Rajagopal, S.; Stickings, P.; Sesardic, D. SiMa Cells for a Serotype Specific and Sensitive Cell-Based Neutralization Test for Botulinum Toxin A and E. Toxins 2017, 9, 230. https://doi.org/10.3390/toxins9070230
Bak N, Rajagopal S, Stickings P, Sesardic D. SiMa Cells for a Serotype Specific and Sensitive Cell-Based Neutralization Test for Botulinum Toxin A and E. Toxins. 2017; 9(7):230. https://doi.org/10.3390/toxins9070230
Chicago/Turabian StyleBak, Nicola, Shalini Rajagopal, Paul Stickings, and Dorothea Sesardic. 2017. "SiMa Cells for a Serotype Specific and Sensitive Cell-Based Neutralization Test for Botulinum Toxin A and E" Toxins 9, no. 7: 230. https://doi.org/10.3390/toxins9070230
APA StyleBak, N., Rajagopal, S., Stickings, P., & Sesardic, D. (2017). SiMa Cells for a Serotype Specific and Sensitive Cell-Based Neutralization Test for Botulinum Toxin A and E. Toxins, 9(7), 230. https://doi.org/10.3390/toxins9070230