
The Myriad Properties of Pasteurella multocida Lipopolysaccharide

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Diseases Caused by Pasteurella multocida
2. P. multocida Virulence Factors
3. General Role of Lipopolysaccharide
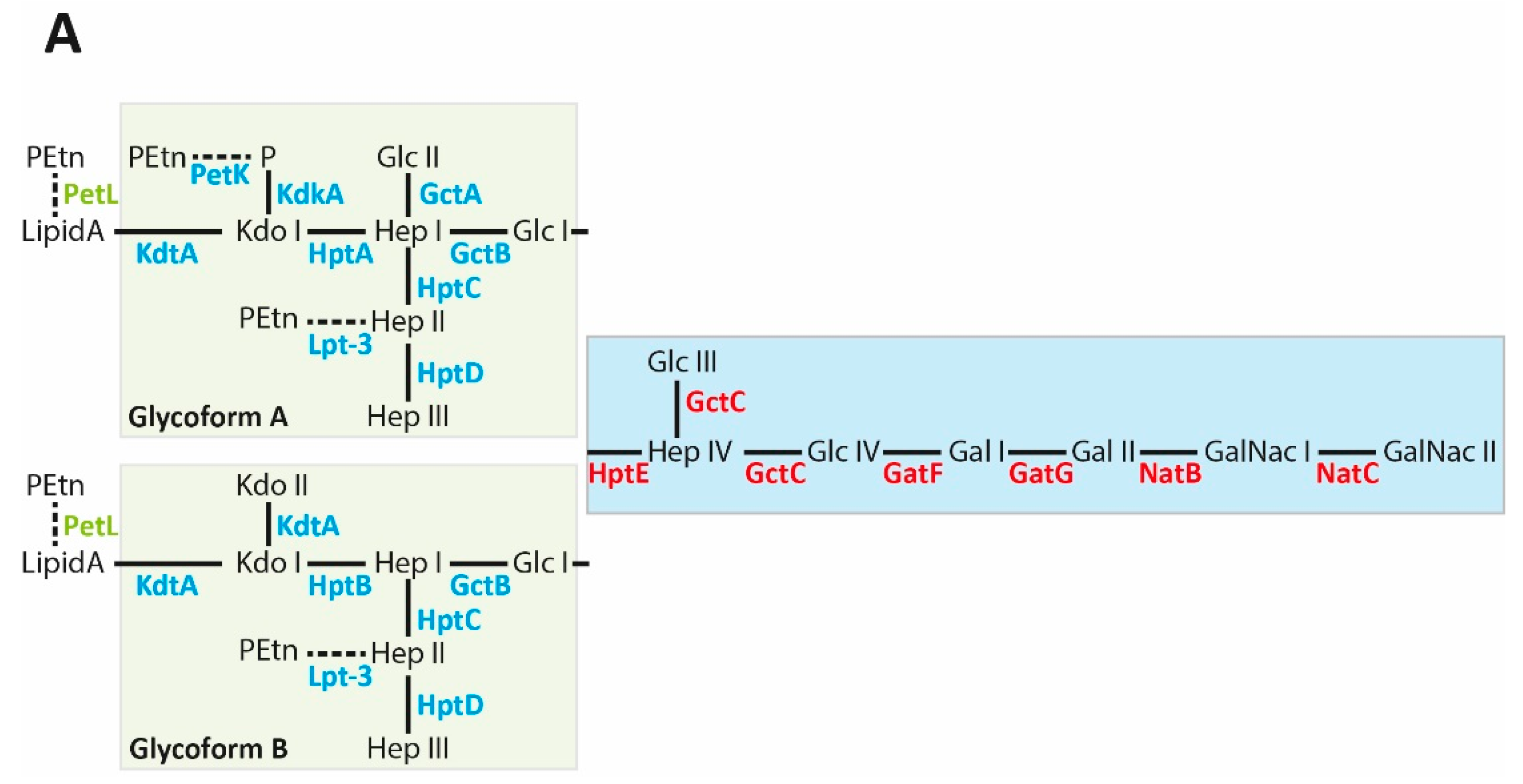
4. P. multocida Strains Produce Two Different LPS Glycoforms
5. The P. multocida LPS Outer Core
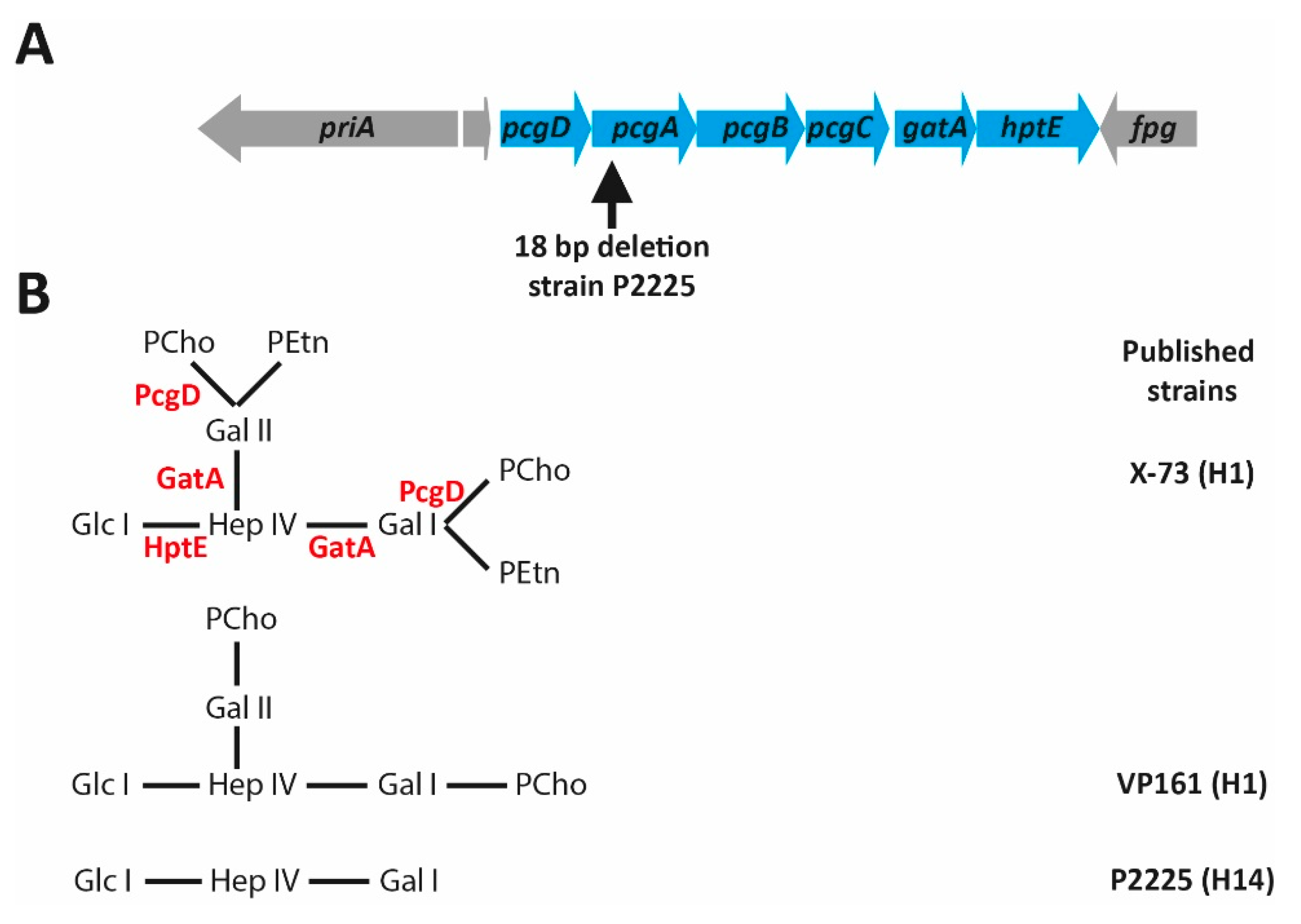
6. The L1 Outer Core Biosynthesis Locus
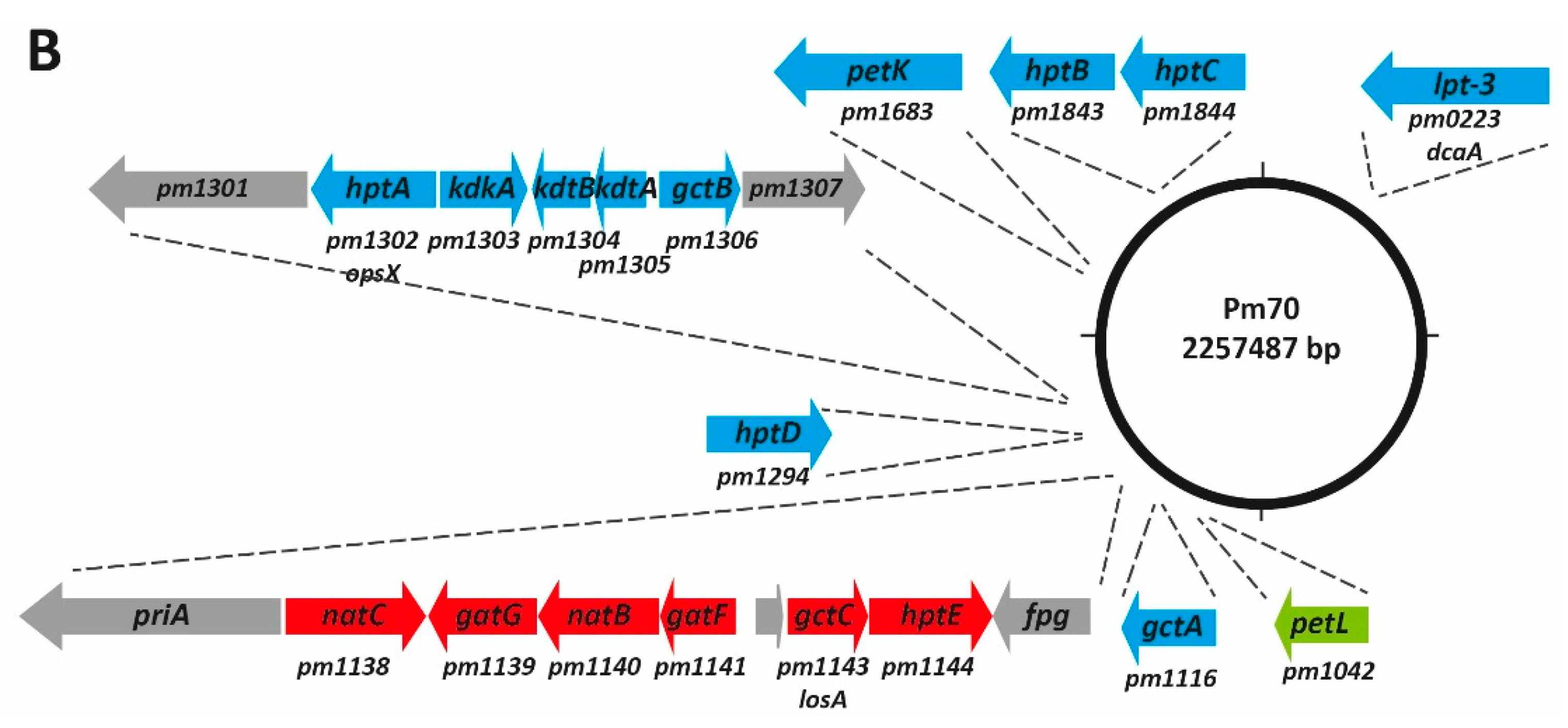
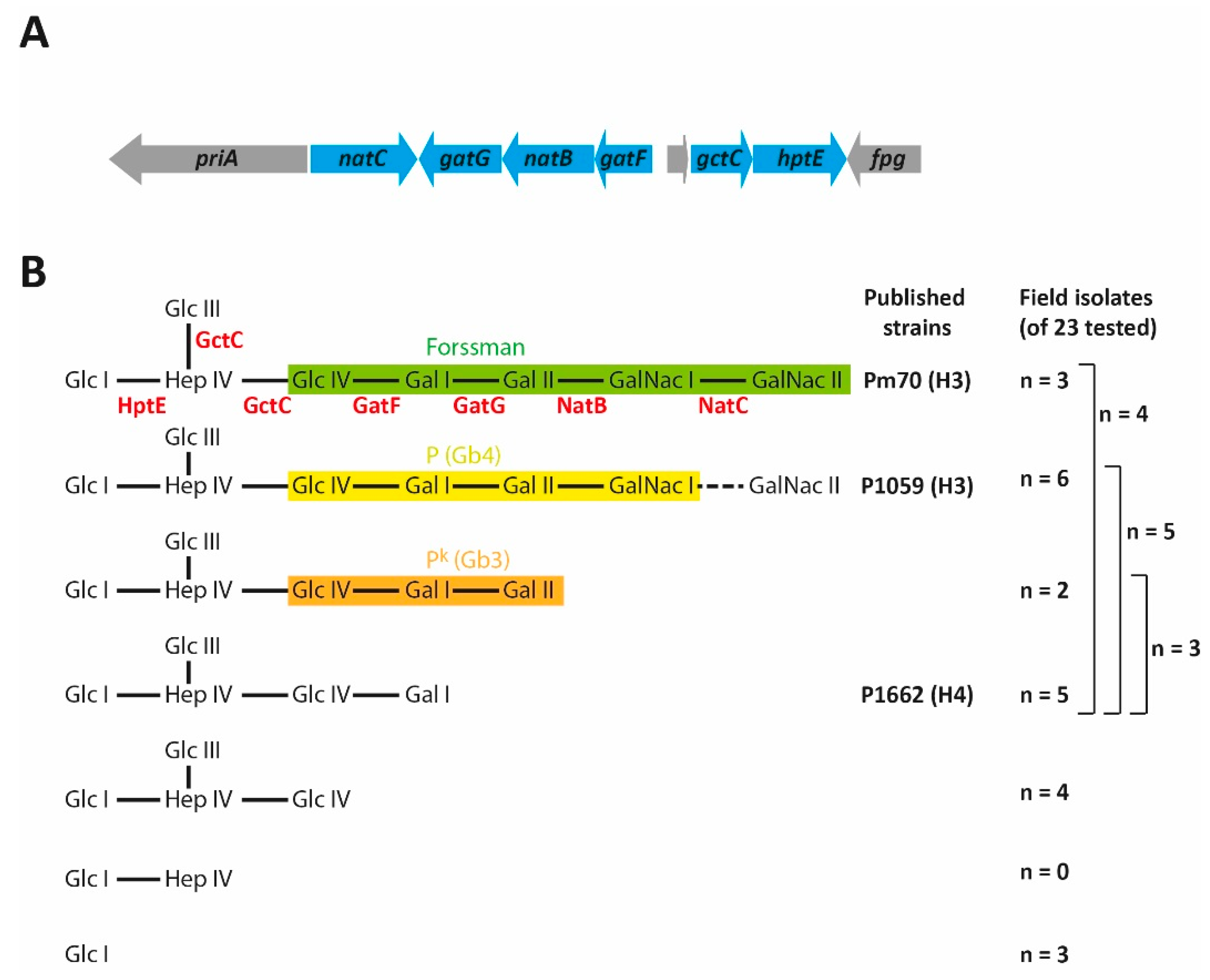
7. The L2 Outer Core Biosynthesis Locus
8. The L3 Outer Core Biosynthesis Locus
9. The L4 and L8 Outer Core Biosynthesis Loci
10. The L5 Outer Core Biosynthesis Locus
11. The L6 Outer Core Biosynthesis Locus
12. The L7 Outer Core Biosynthesis Locus
13. The Role of LPS in P. multocida Strain Typing
14. Role of LPS in P. multocida Pathogenesis
15. Role of LPS in Killed Whole-Cell and Live-Attenuated Vaccines
16. Regulation of LPS Biosynthesis Genes
17. Concluding Remarks
Author Contributions
Conflicts of Interest
References
- Wilkie, I.W.; Harper, M.; Boyce, J.D.; Adler, B. Pasteurella multocida: Diseases and pathogenesis. Curr. Top. Microbiol. Immunol. 2012, 361, 1–22. [Google Scholar] [PubMed]
- Talan, D.A.; Citron, D.M.; Abrahamian, F.M.; Moran, G.J.; Goldstein, E.J. Bacteriologic analysis of infected dog and cat bites. Emergency Medicine Animal Bite Infection Study Group. N. Engl. J. Med. 1999, 340, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Shivachandra, S.B.; Viswas, K.N.; Kumar, A.A. A review of hemorrhagic septicemia in cattle and buffalo. Anim. Health Res. Rev. 2011, 12, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Boyce, J.D.; Harper, M.; Wilkie, I.; Adler, B. Pasteurella. In Pathogenesis of Bacterial Infections in Animals; Gyles, C.L., Prescott, J.F., Songer, J.G., Thoen, C.O., Eds.; Wiley-Blackwell: Ames, IA, USA, 2010; pp. 325–346. [Google Scholar]
- Wilson, B.A.; Ho, M. Pasteurella multocida: From zoonosis to cellular microbiology. Clin. Microbiol. Rev. 2013, 26, 631–655. [Google Scholar] [CrossRef] [PubMed]
- Descamps, S.; Jenouvrier, S.; Gilchrist, H.G.; Forbes, M.R. Avian cholera, a threat to the viability of an Arctic seabird colony? PLoS ONE 2012, 7, e29659. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.A.; Ho, M. Pasteurella multocida toxin interaction with host cells: Entry and cellular effects. Curr. Top. Microbiol. Immunol. 2012, 361, 93–111. [Google Scholar] [PubMed]
- Boyce, J.D.; Adler, B. The capsule is a virulence determinant in the pathogenesis of Pasteurella multocida M1404 (B:2). Infect. Immun. 2000, 68, 3463–3468. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.Y.; Wilkie, I.; Boyce, J.D.; Townsend, K.M.; Frost, A.J.; Ghoddusi, M.; Adler, B. Role of capsule in the pathogenesis of fowl cholera caused by Pasteurella multocida serogroup A. Infect. Immun. 2001, 69, 2487–2492. [Google Scholar] [CrossRef] [PubMed]
- Fuller, T.E.; Kennedy, M.J.; Lowery, D.E. Identification of Pasteurella multocida virulence genes in a septicemic mouse model using signature-tagged mutagenesis. Microb. Pathog. 2000, 29, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Tatum, F.M.; Yersin, A.G.; Briggs, R.E. Construction and virulence of a Pasteurella multocida fhaB2 mutant in turkeys. Microb. Pathog. 2005, 39, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; Cox, A.D.; Michael, F.S.; Wilkie, I.W.; Boyce, J.D.; Adler, B. A heptosyltransferase mutant of Pasteurella multocida produces a truncated lipopolysaccharide structure and is attenuated in virulence. Infect. Immun. 2004, 72, 3436–3443. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; Cox, A.D.; Michael, F.S.; Parnas, H.; Wilkie, I.; Blackall, P.J.; Adler, B.; Boyce, J.D. Decoration of Pasteurella multocida LPS with phosphocholine is important for virulence. J. Bacteriol. 2007, 189, 7384–7391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, M.; John, M.; Edmunds, M.; Wright, A.; Ford, M.; Turni, C.; Blackall, P.; Cox, A.; Adler, B.; Boyce, J.D. Protective efficacy afforded by live Pasteurella multocida vaccines in chickens is independent of lipopolysaccharide structure. Vaccine 2016, in press. [Google Scholar] [CrossRef] [PubMed]
- Heddleston, K.L.; Gallagher, J.E.; Rebers, P.A. Fowl cholera: Gel diffusion precipitin test for serotyping Pasteurella multocida from avian species. Avian Dis. 1972, 16, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; John, M.; Turni, C.; Edmunds, M.; Michael, F.S.; Blackall, P.J.; Cox, A.D.; Boyce, J.D. Development of a rapid multiplex PCR to genotype Pasteurella multocida strains using the lipopolysaccharide outer core biosynthesis locus. J. Clin. Microbiol. 2015, 53, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef] [PubMed]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; Cox, A.D.; Adler, B.; Boyce, J.D. Pasteurella multocida lipopolysaccharide: The long and the short of it. Vet. Microbiol. 2011, 153, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Mandrell, R.E.; Apicella, M.A. Lipo-oligosaccharides (LOS) of mucosal pathogens: Molecular mimicry and host-modification of LOS. Immunobiology 1993, 187, 382–402. [Google Scholar] [CrossRef]
- Weiser, J.N.; Shchepetov, M.; Chong, S.T. Decoration of lipopolysaccharide with phosphorylcholine: A phase-variable characteristic of Haemophilus influenzae. Infect. Immun. 1997, 65, 943–950. [Google Scholar] [PubMed]
- Raetz, C.R.; Reynolds, C.M.; Trent, M.S.; Bishop, R.E. Lipid A modification systems in Gram-negative bacteria. Annu. Rev. Biochem. 2007, 76, 295–329. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Correia, J.; Soldau, K.; Christen, U.; Tobias, P.S.; Ulevitch, R.J. Lipopolysaccharide is in close proximity to each of the proteins in its membrane receptor complex. transfer from CD14 to TLR4 and MD-2. J. Biol. Chem. 2001, 276, 21129–21135. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.; Weaver, C. The Induced Responses of Innate Immunity, in Janeway’s Immunobiology; Garland Science/Taylor and Francis Group, LLC: New York, NY, USA, 2017. [Google Scholar]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [PubMed]
- Seydel, U.; Oikawa, M.; Fukase, K.; Kusumoto, S.; Brandenburg, K. Intrinsic conformation of lipid A is responsible for agonistic and antagonistic activity. Eur. J. Biochem. 2000, 267, 3032–3039. [Google Scholar] [CrossRef] [PubMed]
- Horadagoda, N.U.; Hodgson, J.C.; Moon, G.M.; Wijewardana, T.G.; Eckersall, P.D. Development of a clinical syndrome resembling haemorrhagic septicaemia in the buffalo following intravenous inoculation of Pasteurella multocida serotype B:2 endotoxin and the role of tumour necrosis factor-alpha. Res. Vet. Sci. 2002, 72, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A.; Rhoades, K.R.; Heddleston, K.L. A new serotype of Pasteurella multocida associated with fowl cholera. Avian Dis. 1978, 22, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A.; Rebers, P.A. Serologic examination of the Westphal-type lipopolysaccharides of Pasteurella multocida. Am. J. Vet. Res. 1978, 39, 1680–1682. [Google Scholar] [PubMed]
- Glisson, J.R.; Hofacre, C.L.; Christensen, J.P. Fowl Cholera. In Diseases of Poultry; Swayne, D.E., Ed.; Blackwell Publishing: Ames, IA, USA, 2008; pp. 739–758. [Google Scholar]
- St Michael, F.; Li, J.; Cox, A.D. Structural analysis of the core oligosaccharide from Pasteurella multocida strain X73. Carbohydr. Res. 2005, 340, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- St Michael, F.; Li, J.; Vinogradov, E.; Larocque, S.; Harper, M.; Cox, A.D. Structural analysis of the lipopolysaccharide of Pasteurella multocida strain VP161: Identification of both Kdo-P and Kdo-Kdo species in the lipopolysaccharide. Carbohydr. Res. 2005, 340, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Boyce, J.D.; Harper, M.; Michael, F.S.; John, M.; Aubry, A.; Parnas, H.; Logan, S.M.; Wilkie, I.W.; Ford, M.; Cox, A.D.; et al. Identification of novel glycosyltransferases required for assembly of the Pasteurella multocida A:1 lipopolysaccharide and their involvement in virulence. Infect. Immun. 2009, 77, 1532–1542. [Google Scholar] [CrossRef] [PubMed]
- St Michael, F.; Harper, M.; Parnas, H.; John, M.; Stupak, J.; Vinogradov, E.; Adler, B.; Boyce, J.D.; Cox, A.D. Structural and genetic basis for the serological differentiation of Pasteurella multocida Heddleston serotypes 2 and 5. J. Bacteriol. 2009, 191, 6950–6959. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; St Michael, F.; John, M.; Vinogradov, E.; Adler, B.; Boyce, J.D.; Cox, A.D. Pasteurella multocida Heddleston serovars 1 and 14 express different lipopolysaccharide structures but share the same lipopolysaccharide biosynthesis outer core locus. Vet. Microbiol. 2011, 150, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; St Michael, F.; Vinogradov, E.; John, M.; Boyce, J.D.; Adler, B.; Cox, A.D. Characterization of the lipopolysaccharide from Pasteurella multocida Heddleston serovar 9; identification of a proposed bi-functional dTDP-3-acetamido-3,6-dideoxy-{alpha}-d-glucose biosynthesis enzyme. Glycobiology 2012, 22, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; St Michael, F.; John, M.; Vinogradov, E.; Steen, J.A.; van Dorsten, L.; Steen, J.A.; Turni, C.; Blackall, P.J.; Adler, B.; et al. Pasteurella multocida Heddleston serovar 3 and 4 strains share a common lipopolysaccharide biosynthesis locus but display both inter- and intrastrain lipopolysaccharide heterogeneity. J. Bacteriol. 2013, 195, 4854–4864. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; St Michael, F.; Vinogradov, E.; John, M.; Steen, J.A.; van Dorsten, L.; Boyce, J.D.; Adler, B.; Cox, A.D. Structure and biosynthetic locus of the lipopolysaccharide produced by Pasteurella multocida serovars 8 and 13 and the identification of a novel phospho-glycero moiety. Glycobiology 2013, 23, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; St Michael, F.; Steen, J.A.; John, M.; van Dorsten, L.; Parnas, H.; Vinogradov, E.; Adler, B.; Cox, A.; Boyce, J.D. Structural analysis of lipopolysaccharide produced by Heddleston serovars 10, 11, 12 and 15 and the identification of a new Pasteurella multocida LPS outer core biosynthesis locus, L6. Glycobiology 2014, 24, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; St Michael, F.; Steen, J.A.; John, M.; Wright, C.L.; van Dorsten, L.; Vinogradov, E.; Adler, B.; Cox, A.; Boyce, J.D. Characterisation of the lipopolysaccharide produced by Pasteurella multocida serovars 6, 7 and 16; identification of lipopolysaccharide genotypes L4 and L8. Glycobiology 2015, 25, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; Boyce, J.D.; Cox, A.D.; Michael, F.S.; Wilkie, I.; Blackall, P.; Adler, B. Pasteurella multocida expresses two LPS glycoforms simultaneously both in vitro and in vivo but only a single form is required for virulence: Identification of two acceptor specific heptosyl I transferases. Infect. Immun. 2007, 75, 3885–3893. [Google Scholar] [CrossRef] [PubMed]
- Moxon, R.; Bayliss, C.; Hood, D. Bacterial contingency loci: The role of simple sequence DNA repeats in bacterial adaptation. Annu. Rev. Genet. 2006, 40, 307–333. [Google Scholar] [CrossRef] [PubMed]
- Herget, S.; Toukach, P.V.; Ranzinger, R.; Hull, W.E.; Knirel, Y.A.; von der Lieth, C.W. Statistical analysis of the Bacterial Carbohydrate Structure Data Base (BCSDB): Characteristics and diversity of bacterial carbohydrates in comparison with mammalian glycans. BMC Struct. Biol. 2008, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, R.G.; Sintsova, A.; Buckwalter, C.M.; Leung, N.; Cochrane, A.; Li, J.; Cox, A.D.; Moffat, J.; Gray-Owen, S.D. Cytosolic detection of the bacterial metabolite HBP activates TIFA-dependent innate immunity. Science 2015, 348, 1251–1255. [Google Scholar] [CrossRef] [PubMed]
- Rimler, R.B.; Rebers, P.A.; Phillips, M. Lipopolysaccharides of the Heddleston serotypes of Pasteurella multocida. Am. J. Vet. Res. 1984, 45, 759–763. [Google Scholar] [PubMed]
- Moustafa, A.M.; Seemann, T.; Gladman, S.; Adler, B.; Harper, M.; Boyce, J.D.; Bennett, M.D. Comparative genomic analysis of Asian haemorrhagic septicaemia-associated strains of Pasteurella multocida identifies more than 90 haemorrhagic septicaemia-specific genes. PLoS ONE 2015, 10, e0130296. [Google Scholar] [CrossRef] [PubMed]
- St Michael, F.; Vinogradov, E.; Li, J.; Cox, A.D. Structural analysis of the lipopolysaccharide from Pasteurella multocida genome strain Pm70 and identification of the putative lipopolysaccharide glycosyltransferases. Glycobiology 2005, 15, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Schnaar, R.L.; Suzuki, A.; Stanley, P. Glycosphingolipids. In Essentials of Glycobiology; Varki, A., Chrispeels, M.J., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009. [Google Scholar]
- Shiraishi, T.; Uda, Y. Characterization of neutral sphingolipids from chicken erythrocytes. J. Lipid Res. 1985, 26, 860–866. [Google Scholar] [PubMed]
- Tatum, F.M.; Tabatabai, L.B.; Briggs, R.E. Sialic acid uptake is necessary for virulence of Pasteurella multocida in turkeys. Microb. Pathog. 2009, 46, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Hood, D.W.; Makepeace, K.; Deadman, M.E.; Rest, R.F.; Thibault, P.; Martin, A.; Richards, J.C.; Moxon, E.R. Sialic acid in the lipopolysaccharide of Haemophilus influenzae: Strain distribution, influence on serum resistance and structural characterization. Mol. Microbiol. 1999, 33, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, V.; Hood, D.W.; Li, J.; Brisson, J.R.; Randle, G.A.; Martin, A.; Li, Z.; Goldstein, R.; Schweda, E.K.; Pelton, S.I.; et al. Host-derived sialic acid is incorporated into Haemophilus influenzae lipopolysaccharide and is a major virulence factor in experimental otitis media. Proc. Natl. Acad. Sci. USA 2003, 100, 8898–8903. [Google Scholar] [CrossRef] [PubMed]
- Jurcisek, J.; Greiner, L.; Watanabe, H.; Zaleski, A.; Apicella, M.A.; Bakaletz, L.O. Role of sialic acid and complex carbohydrate biosynthesis in biofilm formation by nontypeable Haemophilus influenzae in the chinchilla middle ear. Infect. Immun. 2005, 73, 3210–3218. [Google Scholar] [CrossRef] [PubMed]
- Blackall, P.J.; Fegan, N.; Chew, G.T.; Hampson, D.J. Population structure and diversity of avian isolates of Pasteurella multocida from Australia. Microbiology 1998, 144 Pt 2, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, B.O.; Heddleston, K.L.; Pfow, C.J. Pasteurella multocida serotyping results (1971–1973). Avian Dis. 1975, 19, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Mandrell, R.E. Further antigenic similarities of Neisseria gonorrhoeae lipooligosaccharides and human glycosphingolipids. Infect. Immun. 1992, 60, 3017–3020. [Google Scholar] [PubMed]
- Mandrell, R.E.; McLaughlin, R.; Kwaik, Y.A.; Lesse, A.; Yamasaki, R.; Gibson, B.; Spinola, S.M.; Apicella, M.A. Lipooligosaccharides (LOS) of some Haemophilus species mimic human glycosphingolipids, and some LOS are sialylated. Infect. Immun. 1992, 60, 1322–1328. [Google Scholar] [PubMed]
- Houliston, R.S.; Vinogradov, E.; Dzieciatkowska, M.; Li, J.; St Michael, F.; Karwaski, M.F.; Brochu, D.; Jarrell, H.C.; Parker, C.T.; Yuki, N.; et al. Lipooligosaccharide of Campylobacter jejuni: Similarity with multiple types of mammalian glycans beyond gangliosides. J. Biol. Chem. 2011, 286, 12361–12370. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.E.; Snow, J.; Li, J.; Zola, T.A.; Weiser, J.N. Phosphorylcholine allows for evasion of bactericidal antibody by Haemophilus influenzae. PLoS Pathog. 2012, 8, e1002521. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.E.; Eichelberger, K.R.; Weiser, J.N. Evasion of killing by human antibody and complement through multiple variations in the surface oligosaccharide of Haemophilus influenzae. Mol. Microbiol. 2013, 88, 603–618. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.L.; Abdullah, F.F.; Ibrahim, H.H.; Marza, A.D.; Zamri-Saad, M.; Haron, A.W.; Lila, M.A.; Norsidin, M.J. Clinico-pathology, hematology and biochemistry responses in buffaloes towards Pasteurella multocida type B:2 immunogen lypopolysaccharide via oral and intravenous routes of infection. Microb. Pathog. 2016, 91, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Marza, A.D.; Abdullah, F.F.J.; Ahmed, I.M.; Chung, E.L.T.; Ibrahim, H.H.; Zamri-Saad, M.; Omar, A.R.; Bakar, M.Z.A.; Saharee, A.A.; Haron, A.W.; et al. The ability of lipopolysaccharide (LPS) of Pasteurella multocida B:2 to induce clinical and pathological lesions in the nervous system of buffalo calves following experimental inoculation. Microb. Pathog. 2017, 104, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Lysenko, E.S.; Gould, J.; Bals, R.; Wilson, J.M.; Weiser, J.N. Bacterial phosphorylcholine decreases susceptibility to the antimicrobial peptide LL-37/hCAP18 expressed in the upper respiratory tract. Infect. Immun. 2000, 68, 1664–1671. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, B.A.; Olson, L.D. Comparison of live avirulent PM-1 and CU fowl cholera vaccines in turkeys. Avian Dis. 1997, 41, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Jaiswal, T.N. Haemorrhagic septicaemia vaccines. Vaccine 1998, 16, 1184–1192. [Google Scholar] [CrossRef]
- Scott, P.C.; Markham, J.F.; Whithear, K.G. Safety and efficacy of two live Pasteurella multocida aro-A mutant vaccines in chickens. Avian Dis. 1999, 43, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, I.W.; Grimes, S.E.; O’Boyle, D.; Frost, A.J. The virulence and protective efficacy for chickens of Pasteurella multocida administered by different routes. Vet. Microbiol. 2000, 72, 57–68. [Google Scholar] [CrossRef]
- Dubin, P.J.; Kolls, J.K. Th17 cytokines and mucosal immunity. Immunol. Rev. 2008, 226, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Noda, K.; Kodama, S.; Umemoto, S.; Nomi, N.; Hirano, T.; Suzuki, M. Th17 cells contribute to nontypeable Haemophilus influenzae-specific protective immunity induced by nasal vaccination with P6 outer membrane protein and alpha-galactosylceramide. Microbiol. Immunol. 2011, 55, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, D.; Heeg, K.; Kubatzky, K.F. Pasteurella multocida toxin manipulates T cell differentiation. Front. Microbiol. 2015, 6, 1273. [Google Scholar] [CrossRef] [PubMed]
- Baba, T. Cell-mediated immune protection in chickens against Pasteurella multocida. Res. Vet. Sci. 1984, 36, 225–230. [Google Scholar] [PubMed]
- Boyce, J.D.; Wilkie, I.; Harper, M.; Paustian, M.L.; Kapur, V.; Adler, B. Genomic-scale analysis of Pasteurella multocida gene expression during growth within liver tissue of chickens with fowl cholera. Microbes Infect. 2004, 6, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Mégroz, M.; Kleifeld, O.; Wright, A.; Powell, D.; Harrison, P.; Adler, B.; Harper, M.; Boyce, J.D. The RNA-binding chaperone Hfq is an important global regulator of gene expression in Pasteurella multocida and plays a crucial role in production of a number of virulence factors including hyaluronic acid capsule. Infect. Immun. 2016, in press. [Google Scholar]
- Steen, J.A.; Steen, J.A.; Harrison, P.; Seemann, T.; Wilkie, I.; Harper, M.; Adler, B.; Boyce, J.D. Fis is essential for capsule production in Pasteurella multocida and regulates expression of other important virulence factors. PLoS Pathog. 2010, 6, e1000750. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harper, M.; Boyce, J.D. The Myriad Properties of Pasteurella multocida Lipopolysaccharide. Toxins 2017, 9, 254. https://doi.org/10.3390/toxins9080254
Harper M, Boyce JD. The Myriad Properties of Pasteurella multocida Lipopolysaccharide. Toxins. 2017; 9(8):254. https://doi.org/10.3390/toxins9080254
Chicago/Turabian StyleHarper, Marina, and John Dallas Boyce. 2017. "The Myriad Properties of Pasteurella multocida Lipopolysaccharide" Toxins 9, no. 8: 254. https://doi.org/10.3390/toxins9080254
APA StyleHarper, M., & Boyce, J. D. (2017). The Myriad Properties of Pasteurella multocida Lipopolysaccharide. Toxins, 9(8), 254. https://doi.org/10.3390/toxins9080254