Highly Localized Enrichment of Trypanosoma brucei Parasites Using Dielectrophoresis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Experimental Methods
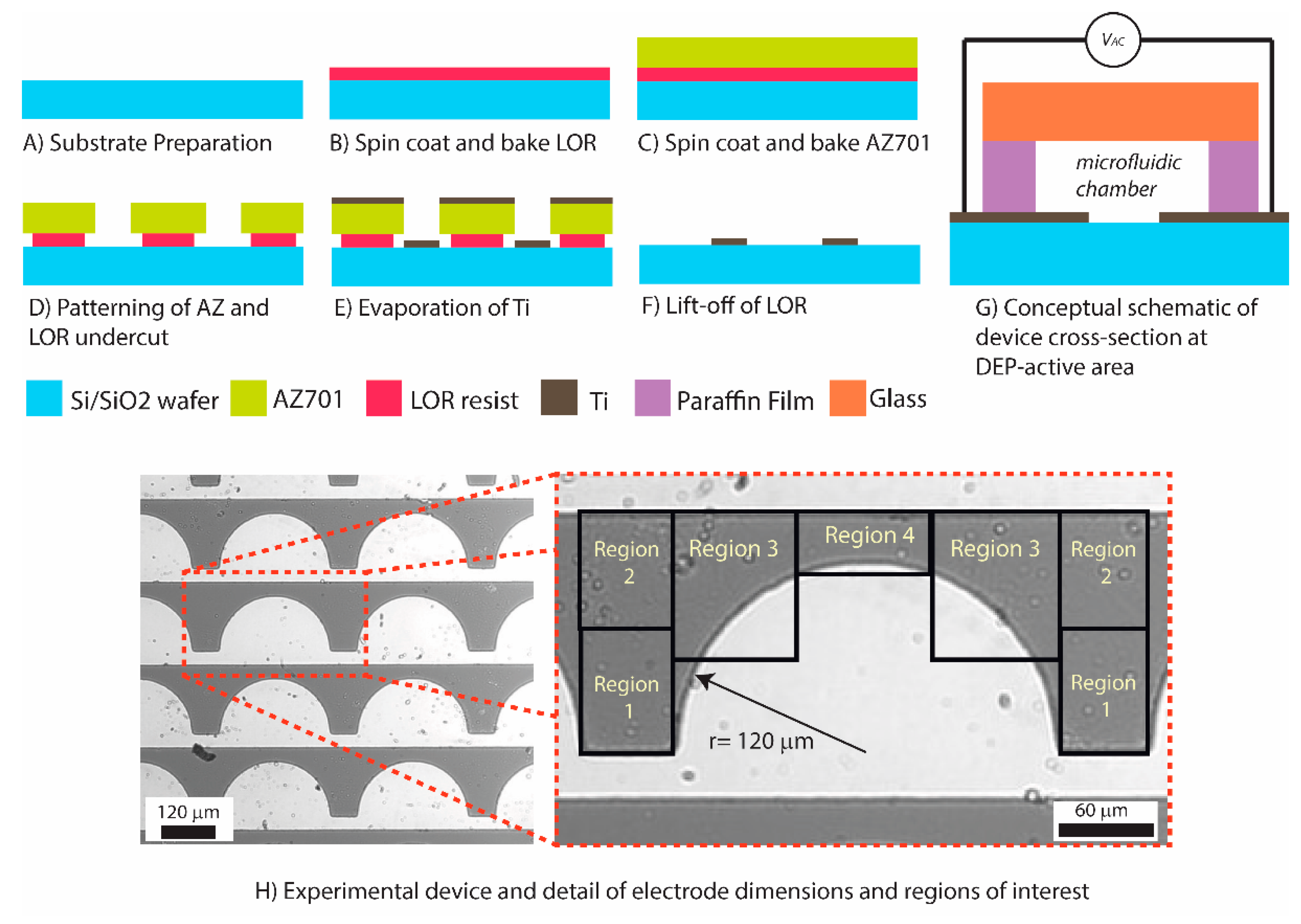
2.1. Fabrication of the Experimental Device
2.2. Sample Preparation and Viability Study
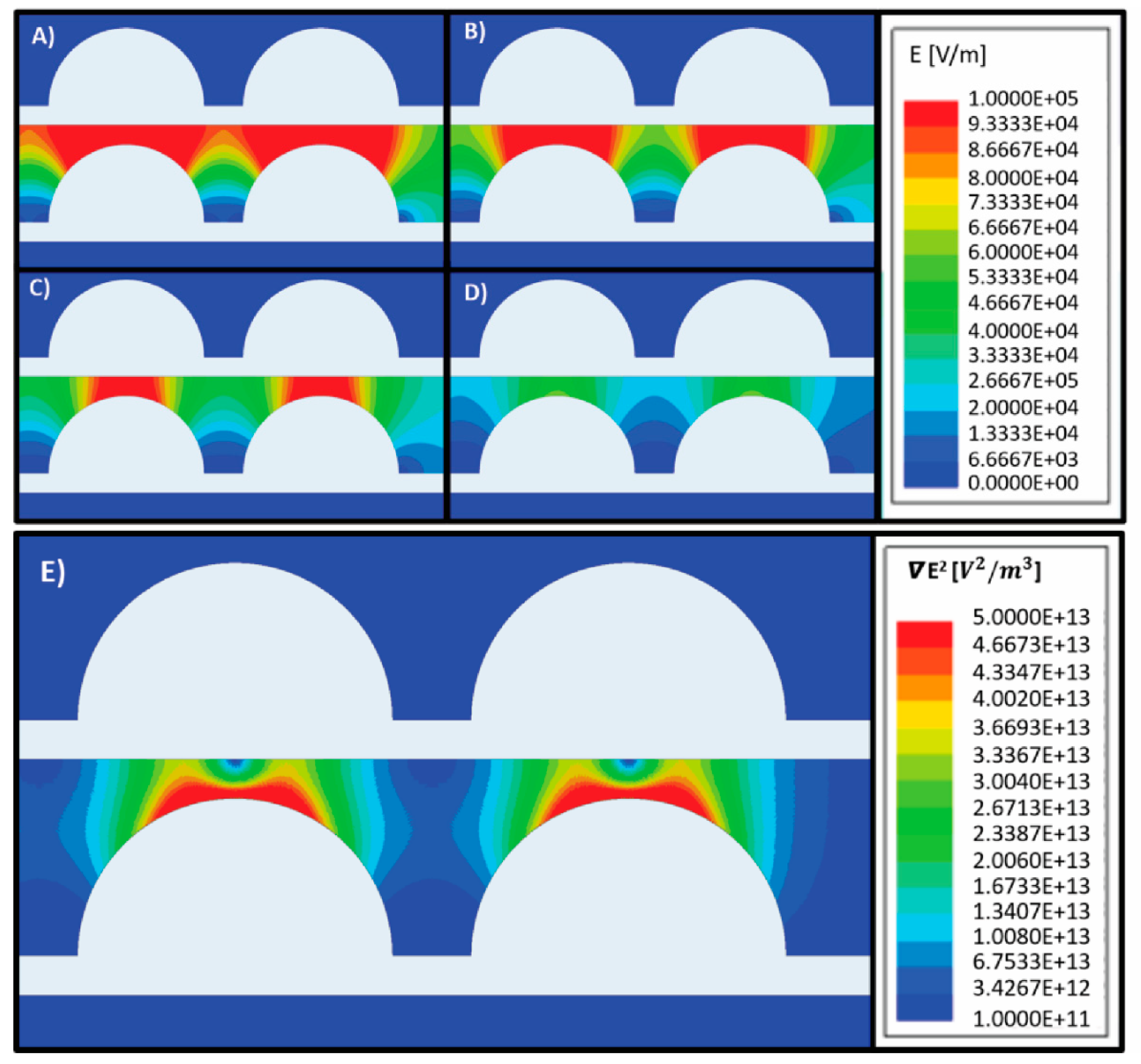
2.3. Computational Modeling
2.4. Protocol for the Dielectrophoretic and Regional Enrichment Characterization of the Parasites
3. Analytical Methods
3.1. Dielectrophoretic Characterization of the Parasites
3.2. Assessment of the Regional Enrichment of the Parasites
4. Results and Discussion
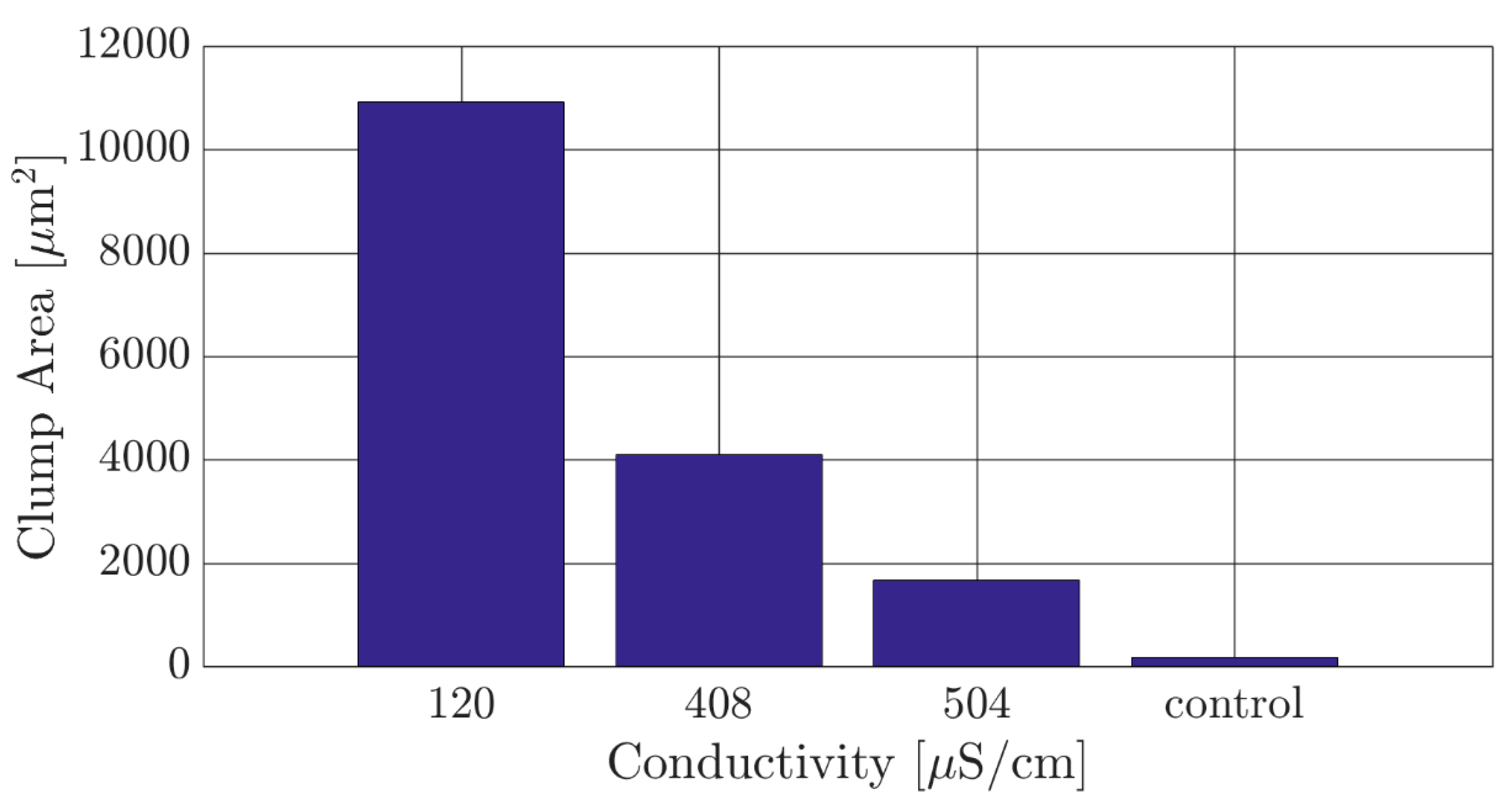
4.1. Optimizing the Experimental DEP Media for T. brucei
4.2. Optimizing Polarization Voltage for DEP Experiments
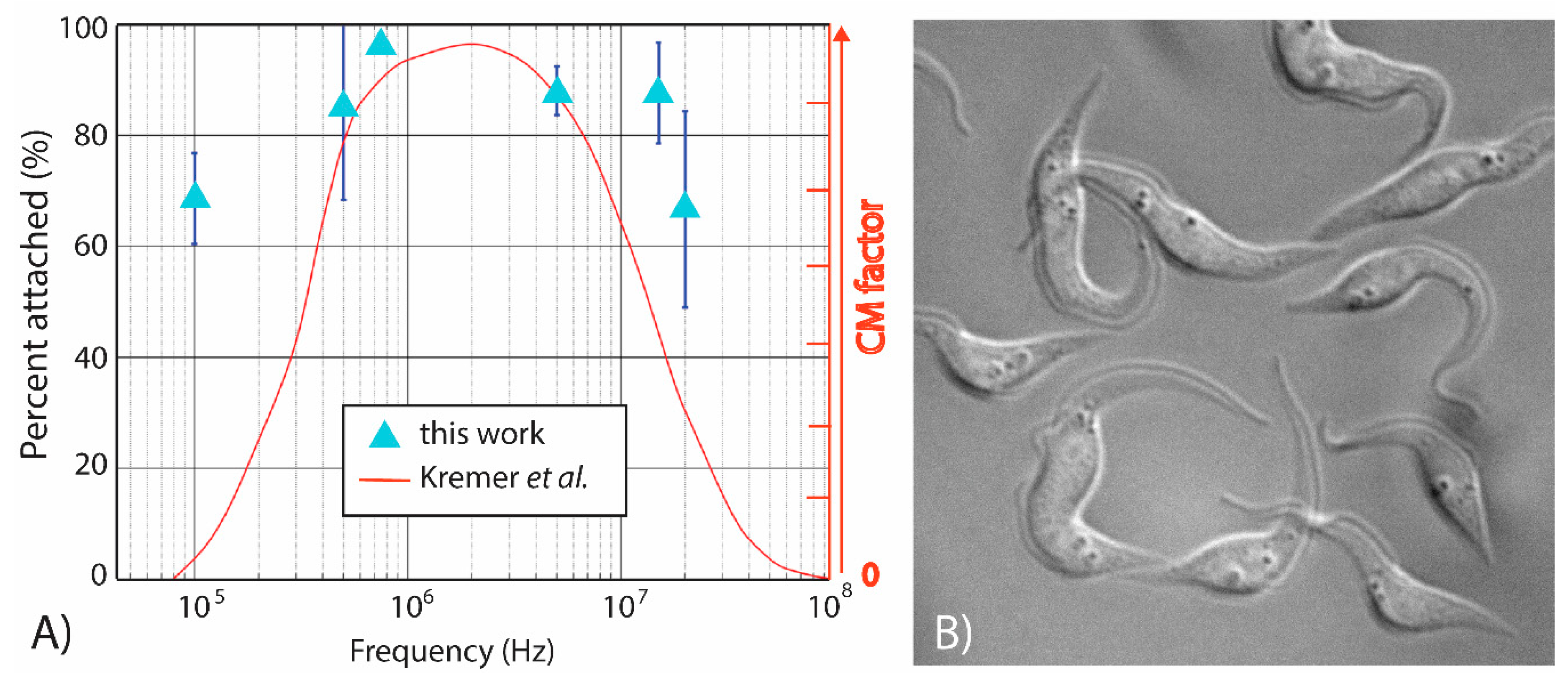
4.3. Characterizing the Dielectrophoretic Response of T. brucei
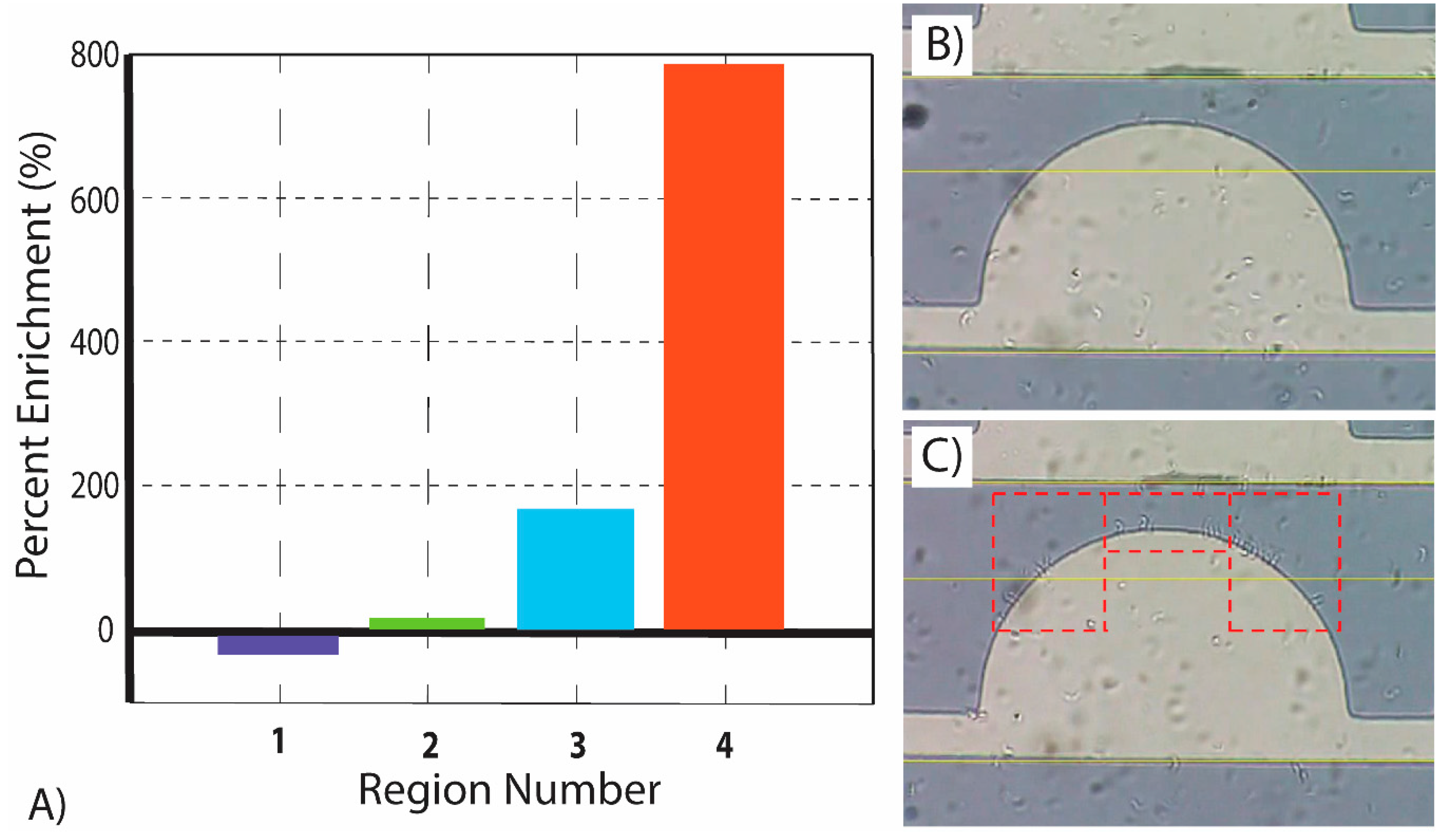
4.4. Determining the Region that Yields the Highest Enrichment of T. brucei
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Babokhov, P.; Sanyaolu, A.; Oyibo, W.; Fagbenro-Beyioku, A.F.; Iriemenam, N.C. A current analysis of chemotherapy strategies for the treatment of human African trypanosomiasis. Pathog. Glob. Health 2013, 107, 242–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chappuis, F.; Loutan, L.; Simarro, P.; Lejon, V.; Büscher, P. Options for Field Diagnosis of Human African Trypanosomiasis. Clin. Microbiol. Rev. 2005, 18, 133–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holm, S.H.; Beech, J.P.; Barrett, M.P.; Tegenfeldt, J.O. Separation of parasites from human blood using deterministic lateral displacement. Lab Chip 2011, 11, 1326. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, J.; Boudot, C.; Courtioux, B. Overview of the Diagnostic Methods Used in the Field for Human African Trypanosomiasis: What Could Change in the Next Years? BioMed. Res. Int. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, N.G.; Loderstaedt, U.; Hahn, A.; Hinz, R.; Zautner, A.E.; Eibach, D.; Fischer, M.; Hagen, R.M.; Frickmann, H. Microbiological laboratory diagnostics of neglected zoonotic diseases (NZDs). Acta Trop. 2017, 165, 40–65. [Google Scholar] [CrossRef]
- Lumbala, C.; Biéler, S.; Kayembe, S.; Makabuza, J.; Ongarello, S.; Ndung’U, J.M. Prospective evaluation of a rapid diagnostic test for Trypanosoma brucei gambiense infection developed using recombinant antigens. PLoS Negl. Trop. Dis. 2018, 12, e0006386. [Google Scholar] [CrossRef] [Green Version]
- Lutumba, P.; Robays, J.; Miaka, C.; Kande, V.; Mumba, D.; Büscher, P.; Dujardin, B.; Boelaert, M. Validité, coût et faisabilité de la mAECT et CTC comme tests de confirmation dans la détection de la Trypanosomiase Humaine Africaine. Trop. Med. Int. Health 2006, 11, 470–478. [Google Scholar] [CrossRef] [Green Version]
- Ngoyi, D.M.; Ekangu, R.A.; Kodi, M.F.M.; Pyana, P.P.; Balharbi, F.; Decq, M.; Betu, V.K.; Van Der Veken, W.; Sese, C.; Menten, J.; et al. Performance of Parasitological and Molecular Techniques for the Diagnosis and Surveillance of Gambiense Sleeping Sickness. PLOS Negl. Trop. Dis. 2014, 8, e2954. [Google Scholar] [CrossRef] [Green Version]
- Gimsa, J. A comprehensive approach to electro-orientation, electrodeformation, dielectrophoresis, and electrorotation of ellipsoidal particles and biological cells. Bioelectrochemistry 2001, 54, 23–31. [Google Scholar] [CrossRef]
- Parkash, O.; Kumar, D.; Prasad, C.D. A unified theory of dielectrophoresis and travelling wave dielectrophoresis. J. Phys. D Appl. Phys. 1994, 27, 1509–1512. [Google Scholar] [CrossRef]
- Jones, T.B. Basic theory of dielectrophoresis and electrorotation. IEEE Eng. Med. Biol. Mag. 2003, 22, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Gascoyne, P.R.C.; Vykoukal, J. Dielectrophoresis-Based Sample Handling in General-Purpose Programmable Diagnostic Instruments. Proc. IEEE Inst. Electr. Electron. Eng. 2004, 92, 22–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.-B.; Hughes, M.P.; Huang, Y.; Becker, F.; Gascoyne, P. Non-uniform spatial distributions of both the magnitude and phase of AC electric fields determine dielectrophoretic forces. Biochim. et Biophys. Acta (BBA) Gen. Subj. 1995, 1243, 185–194. [Google Scholar] [CrossRef]
- Green, N.G.; Ramos, A.; Morgan, H. Numerical solution of the dielectrophoretic and travelling wave forces for interdigitated electrode arrays using the finite element method. J. Electrost. 2002, 56, 235–254. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, Z.R. Cellular dielectrophoresis: Applications to the characterization, manipulation, separation and patterning of cells. Electrophoresis 2011, 32, 2466–2487. [Google Scholar] [CrossRef]
- Fernandez, R.E.; Rohani, A.; Farmehini, V.; Swami, N.S. Review: Microbial analysis in dielectrophoretic microfluidic systems. Anal. Chim. Acta 2017, 966, 11–33. [Google Scholar] [CrossRef] [Green Version]
- Kremer, C.; Neale, S.L.; Menachery, A.; Barrett, M.; Cooper, J.M. Optoelectronic tweezers for medical diagnostics. SPIE BiOS 2012, 8212, 82120. [Google Scholar] [CrossRef]
- Menachery, A.; Kremer, C.; Wong, P.E.; Carlsson, A.; Neale, S.L.; Barrett, M.P.; Cooper, J.M. Counterflow Dielectrophoresis for Trypanosome Enrichment and Detection in Blood. Sci. Rep. 2012, 2, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Duarte, R. Microfabrication technologies in dielectrophoresis applications—A review. Electrophoresis 2012, 33, 3110–3132. [Google Scholar] [CrossRef]
- Sun, T.; Morgan, H.; Green, N.G. Analytical solutions of ac electrokinetics in interdigitated electrode arrays: Electric field, dielectrophoretic and traveling-wave dielectrophoretic forces. Phys. Rev. E 2007, 76, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.T.; Bottausci, F.; Rao, M.P.; Parker, E.R.; Mezić, I.; Macdonald, N.C. Titanium-based dielectrophoresis devices for microfluidic applications. Biomed. Microdevices 2008, 10, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Parker, E.R.; Rao, M.P.; Aimi, M.F.; Mezic, I.; MacDonald, N.C. Titanium bulk micromachining for biomems applications: A DEP device as a demonstration. In Proceedings of the IMECE: 2004 ASME International Mechanical Engineering Congress and Rd & D Expo, Anaheim, CA, USA, 13–19 November 2004. [Google Scholar]
- Tanaka, Y.; Endo, T.; Yanagida, Y.; Hatsuzawa, T. Design and fabrication of a dielectrophoresis-based cell-positioning and cell-culture device for construction of cell networks. Microchem. J. 2009, 91, 232–238. [Google Scholar] [CrossRef]
- Puttaswamy, S.V.; Sivashankar, S.; Chen, R.-J.; Chin, C.-K.; Chang, H.-Y.; Liu, C.-H. Enhanced cell viability and cell adhesion using low conductivity medium for negative dielectrophoretic cell patterning. Biotechnol. J. 2010, 5, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Natu, R.; Islam, M.; Martinez-duarte, R. Carbon cone electrodes for selection, manipulation and lysis of single cells. In Proceedings of the ASME 2015 International Mechanical Engineering Congress and Exposition IMECE 2015, Houston, TX, USA, 13–19 November 2015; pp. 1–5. [Google Scholar]
- Brun, R.; Jenni, L.; Tanner, M.; Schönenberger, M.; Schell, K.F. Cultivation of vertebrate infective forms derived from metacyclic forms of pleomorphic Trypanosoma brucei stocks. Acta Trop. 1979, 36, 387–390. [Google Scholar] [PubMed]
- Sharma, R.; Peacock, L.; Gluenz, E.; Gull, K.; Gibson, W.; Carrington, M. Asymmetric Cell Division as a Route to Reduction in Cell Length and Change in Cell Morphology in Trypanosomes. Protist 2008, 159, 137–151. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Glasser, H.; Fuhr, G. Cultivation of cells under strong ac-electric field—Differentiation between heating and trans-membrane potential effects. BioElectrochem. Bioenerg. 1998, 47, 301–310. [Google Scholar] [CrossRef]
- Pethig, R.; Menachery, A.; Pells, S.; De Sousa, P. Dielectrophoresis: A Review of Applications for Stem Cell Research. J. Biomed. Biotechnol. 2010, 2010, 1–7. [Google Scholar] [CrossRef]
- Pethig, R. Encyclopedia of Surface and Colloidal Science; Taylor & Francis: New York, NY, USA, 2006. [Google Scholar]
- Field, M.C.; Allen, C.L.; Dhir, V.; Goulding, D.; Hall, B.; Morgan, G.W.; Veazey, P.; Engstler, M. New Approaches to the Microscopic Imaging of Trypanosoma brucei. Microsc. Microanal. 2004, 10, 621–636. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keck, D.; Stuart, C.; Duncan, J.; Gullette, E.; Martinez-Duarte, R. Highly Localized Enrichment of Trypanosoma brucei Parasites Using Dielectrophoresis. Micromachines 2020, 11, 625. https://doi.org/10.3390/mi11060625
Keck D, Stuart C, Duncan J, Gullette E, Martinez-Duarte R. Highly Localized Enrichment of Trypanosoma brucei Parasites Using Dielectrophoresis. Micromachines. 2020; 11(6):625. https://doi.org/10.3390/mi11060625
Chicago/Turabian StyleKeck, Devin, Callie Stuart, Josie Duncan, Emily Gullette, and Rodrigo Martinez-Duarte. 2020. "Highly Localized Enrichment of Trypanosoma brucei Parasites Using Dielectrophoresis" Micromachines 11, no. 6: 625. https://doi.org/10.3390/mi11060625
APA StyleKeck, D., Stuart, C., Duncan, J., Gullette, E., & Martinez-Duarte, R. (2020). Highly Localized Enrichment of Trypanosoma brucei Parasites Using Dielectrophoresis. Micromachines, 11(6), 625. https://doi.org/10.3390/mi11060625