Deuterated Glutamate-Mediated Neuronal Activity on Micro-Electrode Arrays

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primary Cultured Hippocamal Neuronal Network on MEA
2.2. Control of GLU and GLU-D Concentrations and Pharamacological Treatments
2.3. Raman Spectroscopy
2.4. Extracellular Potential Recording
3. Results and Discussion
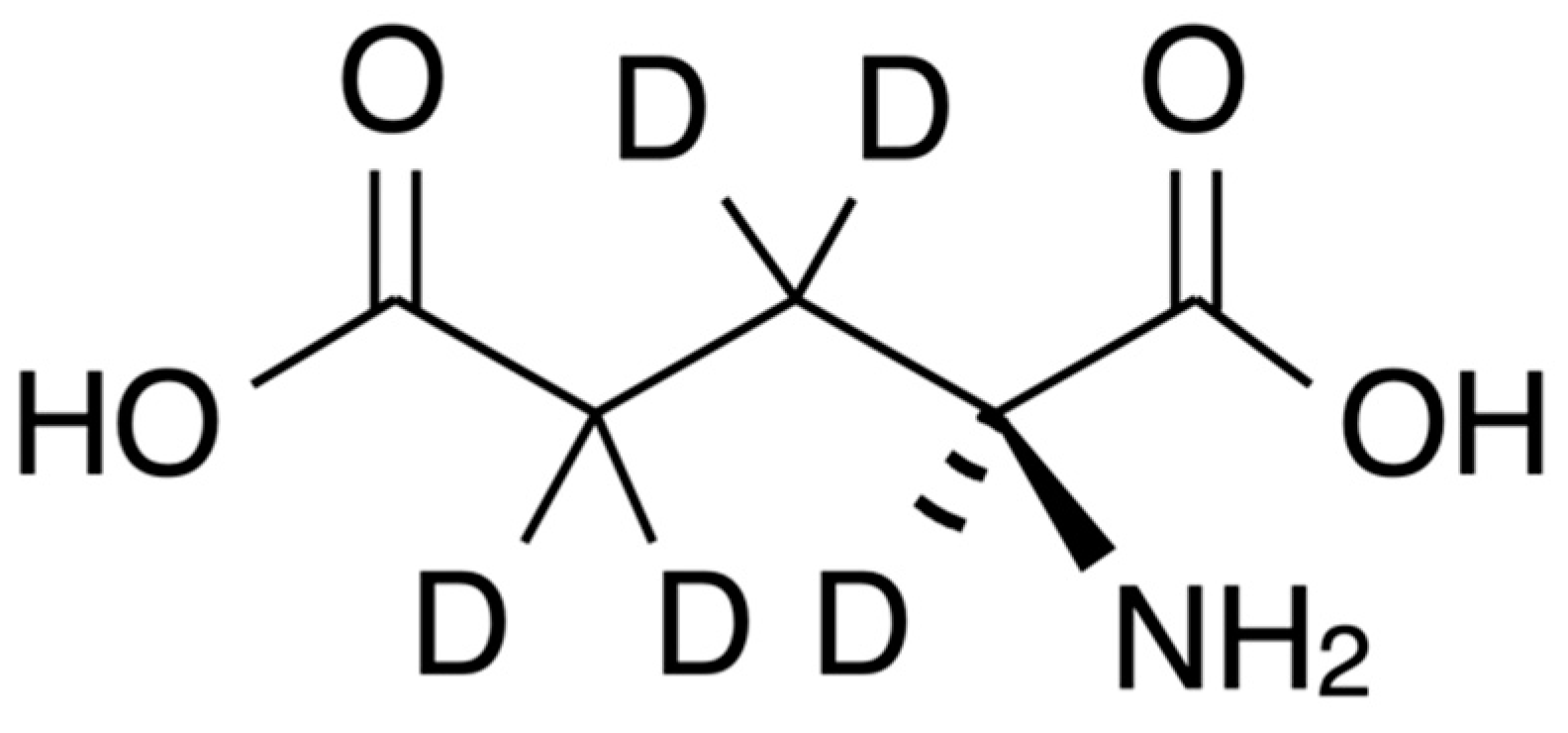
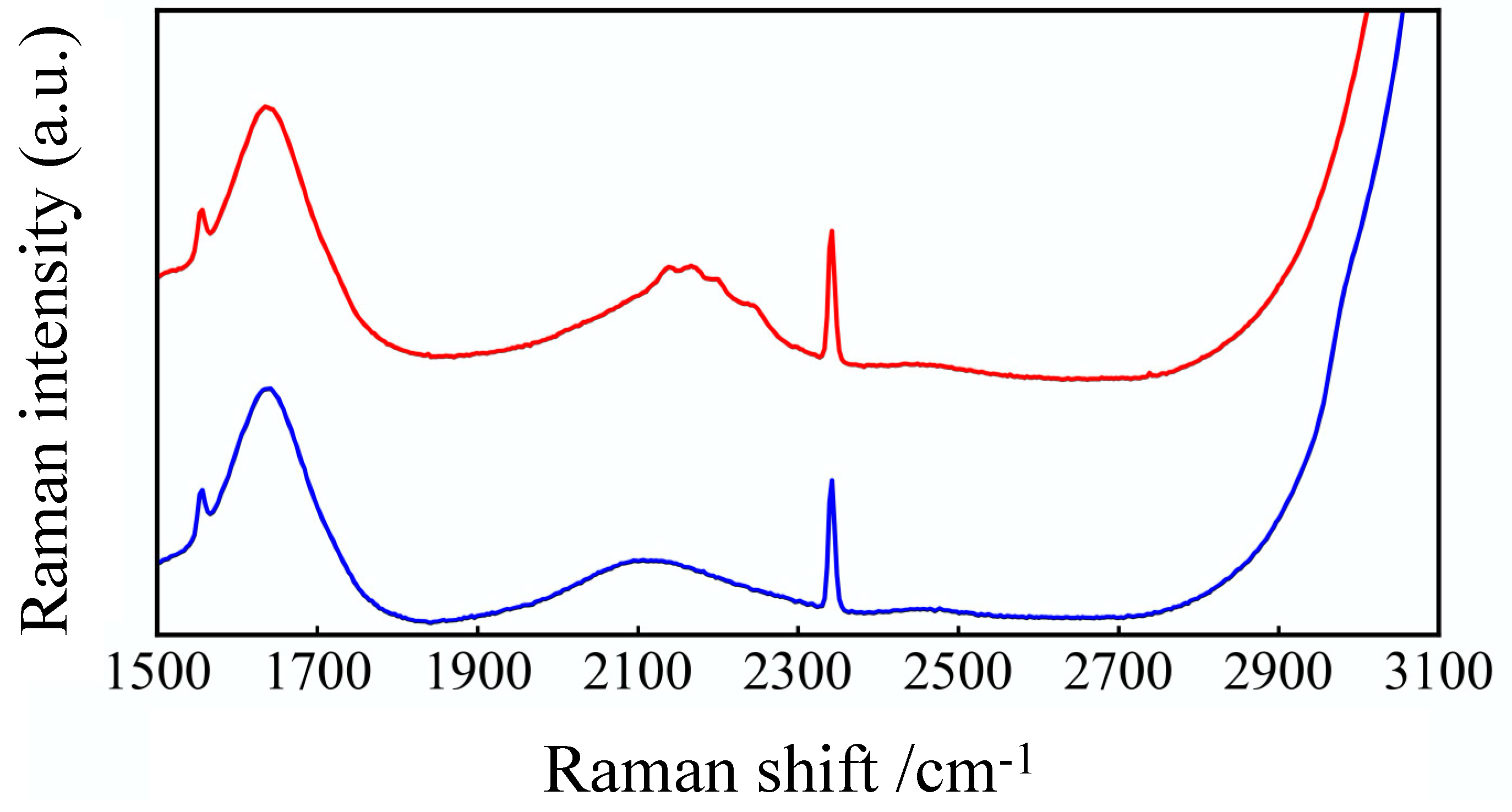
3.1. Raman-Spectra of GLU and GLU-D Solutions
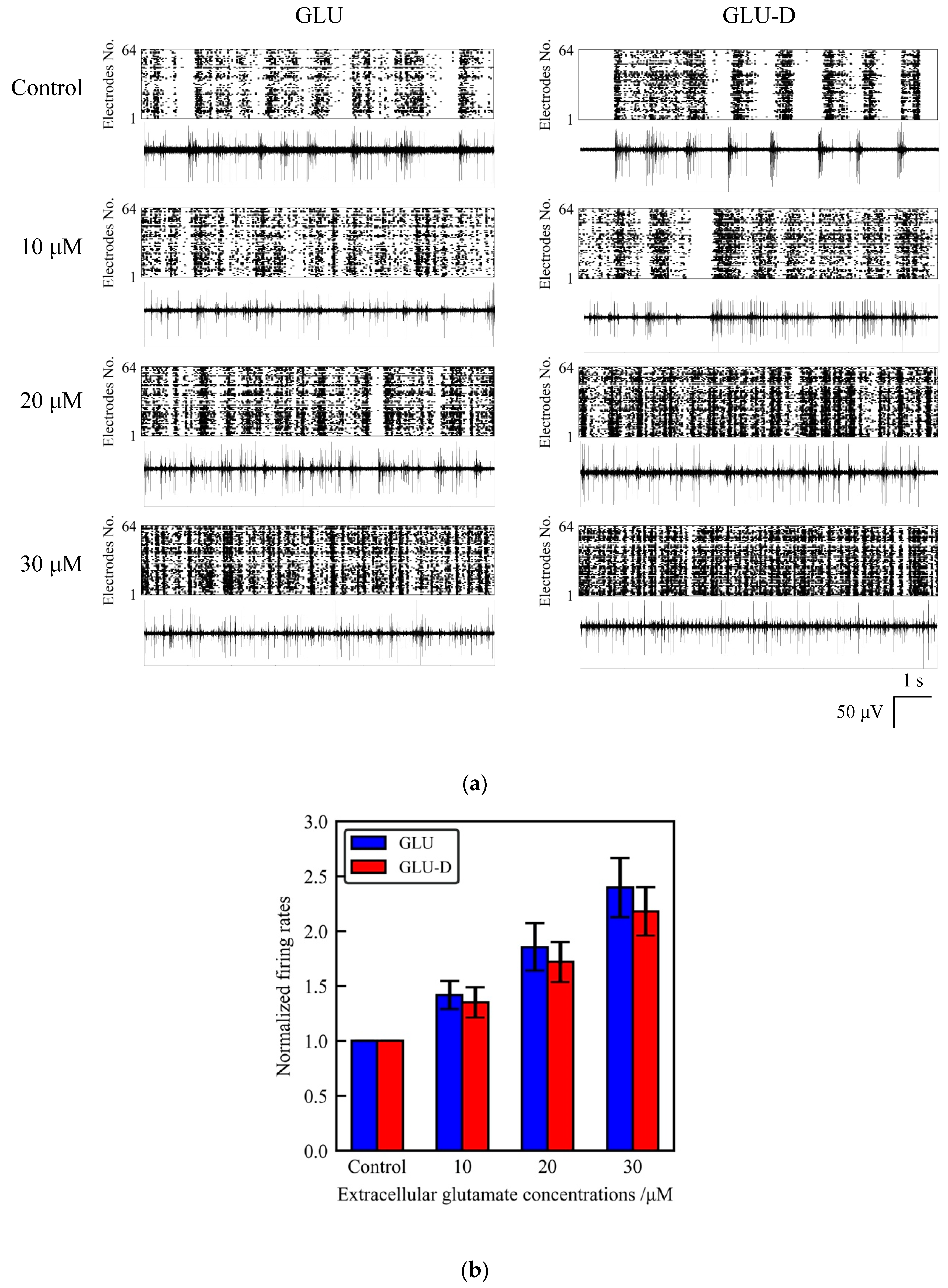
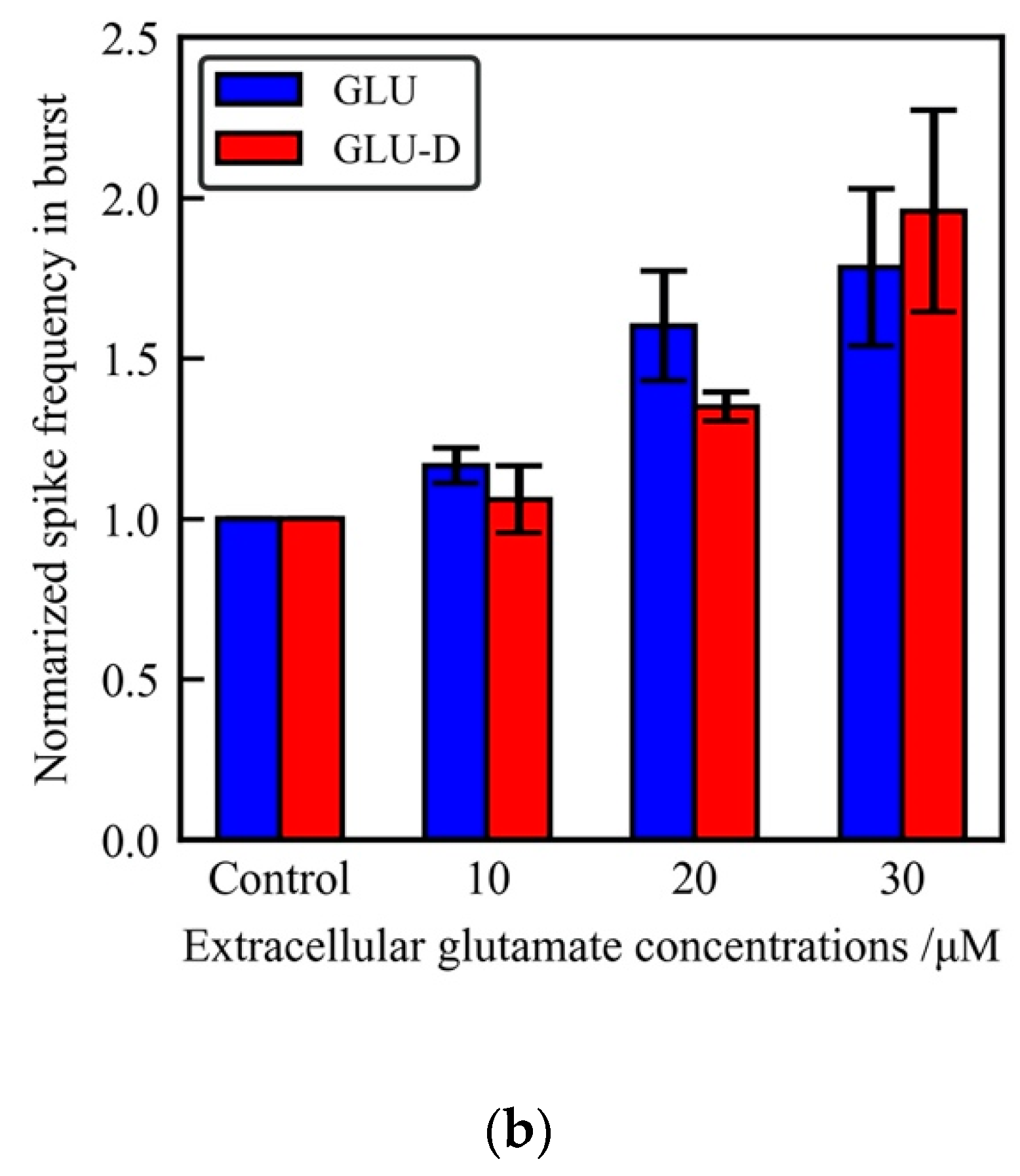
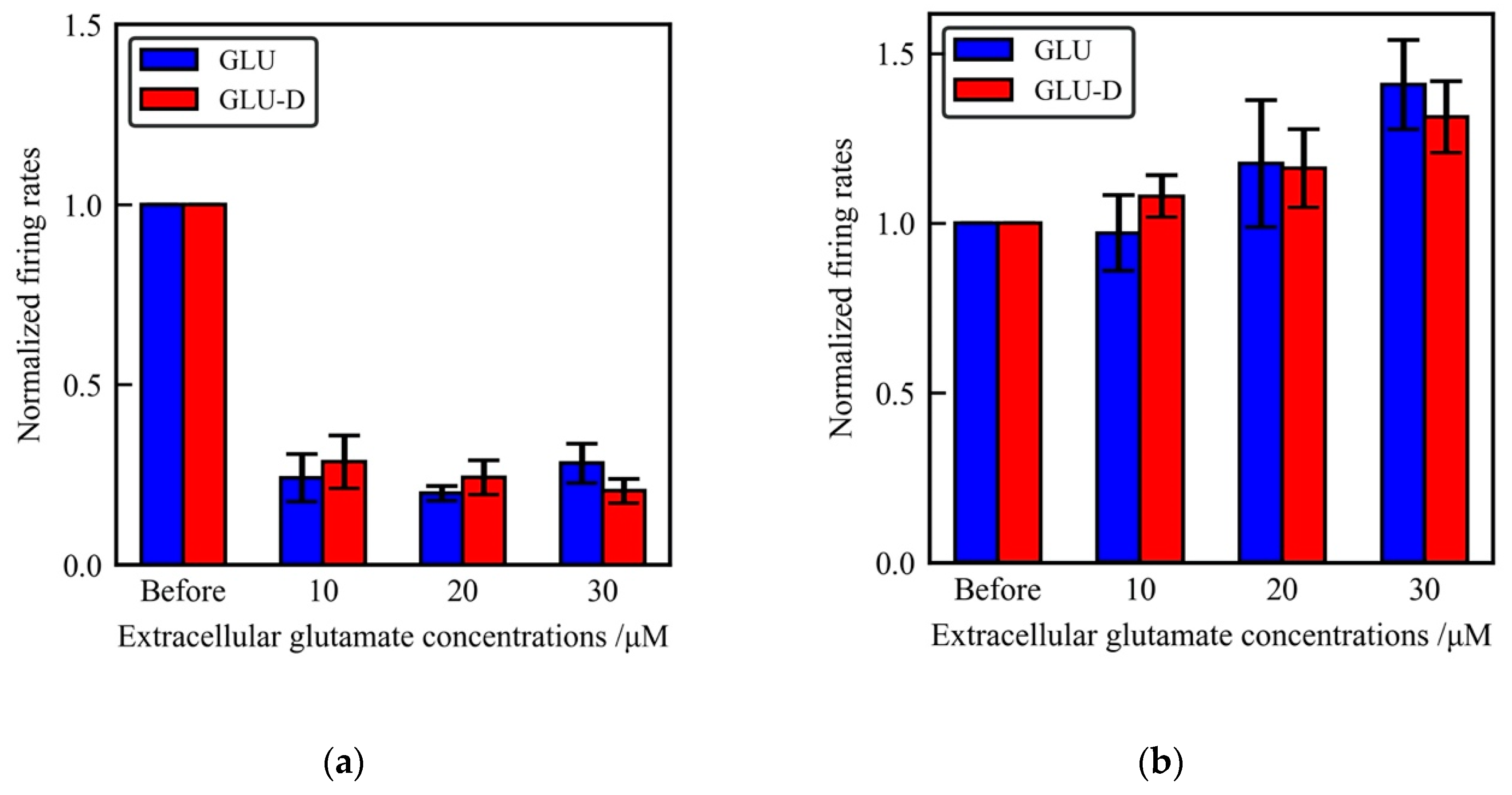
3.2. Effects of Extracellular GLU and GLU-D on the Spontaneous Activity of Cultured Hippocampal Neurons
3.3. Effects of Glutamate Receptors Blockers on Extracellular Glutamate- Induced Neuronal Activity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fischer, M.; Kaech, S.; Wagner, U.; Brinkhaus, H.; Matus, A. Glutamate receptors regulate actin-based plasticity in dendritic spines. Nat. Neurosci. 2000, 3, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Okabe, S.; Miwa, A.; Okado, H. Spine formation and correlated assembly of presynaptic and postsynaptic molecules. J. Neurosci. 2001, 21, 6105–6114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewer, G.J. Isolation and culture of adult rat hippocampal neurons. J. Neurosci. Methods 1997, 71, 143–155. [Google Scholar] [CrossRef]
- Banker, G.; Goslin, K. Culturing Nerve Cells; MIT Press: Cambridge, UK, 1991. [Google Scholar]
- Lesuisse, C.; Martin, L.J. Long-term culture of mouse cortical neurons as a model for neuronal development, aging, and death. J. Neurobiol. 2002, 51, 9–23. [Google Scholar] [CrossRef]
- Optiz, T.; De Lima, A.D.; Voigt, T. Spontaneous development of synchronous oscillatory activity during maturation of cortical networks in vitro. J. Neurophysiol. 2002, 88, 2196–2206. [Google Scholar] [CrossRef]
- Fletcher, T.L.; Cameron, T.L.; De Camilli, P.; Banker, G.A. The distribution of synapsin I and synaptophysin in hippocampal neurons developing in culture. J. Neurosci. 1991, 11, 1617–1626. [Google Scholar] [CrossRef] [Green Version]
- Benson, D.L.; Cohen, P.A. Activity-independent segregation of excitatory and inhibitory synaptic terminals in cultured hippocampal neurons. J. Neurosci. 1996, 16, 6424–6432. [Google Scholar] [CrossRef]
- van Pelt, J.; Wolters, P.S.; Corner, M.A.; Rutten, W.L.; Ramakers, G.J. Long-term characterization of firing dynamics of spontaneous bursts in cultured neural networks. IEEE Trans. Biomed. Eng. 2004, 51, 2051–2062. [Google Scholar] [CrossRef]
- Eytan, D.; Marom, S. Dynamics and effective topology underlying synchronization in networks of cortical neurons. J. Neurosci. 2006, 26, 8465–8476. [Google Scholar] [CrossRef] [Green Version]
- Madhavan, R.; Chao, Z.C.; Potter, S.M. Plasticity of recurring spatiotemporal activity patterns in cortical networks. Phys. Biol. 2007, 4, 181–193. [Google Scholar] [CrossRef]
- Kiyohara, A.; Taguchi, T.; Kudoh, S.N. Effects of electrical stimulation on autonomous electrical activity in a cultured rat hippocampal neuronal network. IEEJ Trans. Electr. Electr. 2011, 6, 163–167. [Google Scholar] [CrossRef]
- Napoli, A.; Obeid, I. Comparative analysis of human and rodent brain primary neuronal Culture spontaneous activity using micro-electrode array technology. J. Cell Biochem. 2016, 117, 559–565. [Google Scholar] [CrossRef]
- Maass, W.; Natschläger, T.; Markram, H. Real-time computing without stable states: A new framework for neural computation based on perturbations. Neural Comput. 2002, 14, 2531–2560. [Google Scholar] [CrossRef] [PubMed]
- Ikegaya, Y.; Aaron, G.; Cossart, R.; Aronov, D.; Lampl, I.; Ferster, D.; Yuste, R. Synfire chains and cortical songs: Temporal modules of cortical activity. Science 2004, 304, 559–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sussillo, D.; Abbott, L.F. Generating coherent patterns of activity from chaotic neural networks. Neuron 2009, 63, 544–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, D.; Tamate, H.; Nagayama, M.; Uchida, T.; Kudoh, S.N.; Gohara, K. Minimum neuron density for synchronized bursts in a rat cortical culture on multi-electrode arrays. Neurosicence 2010, 171, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Massobrio, P.; Tessadori, J.; Chiappalone, M.; Ghirardi, M. In vitro studies of neuronal networks and synaptic plasticity in invertebrates and in mammals using multielectrode arrays. Neural Plast. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napoli, A.; Obeid, I. Investigating brain functional evolution and plasticity using microelectrode array technology. Brain Res. Bull. 2015, 119, 127–135. [Google Scholar] [CrossRef]
- Odawara, A.; Matsuda, N.; Ishibashi, Y.; Yokoi, R.; Suzuki, I. Toxicological evaluation of convulsant and anticonvulsant drugs in human induced pluripotent stem cell-derived cortical neuronal networks using an MEA system. Sci. Rep. 2018, 8, 10416. [Google Scholar] [CrossRef]
- Iida, S.; Shimba, K.; Sakai, K.; Kotani, K.; Jimbo, Y. Synchronous firing patterns of induced pluripotent stem cell-derived cortical neurons depend on the network structure consisting of excitatory and inhibitory neurons. Biochem. Biophys. Res. Commun. 2018, 501, 152–157. [Google Scholar] [CrossRef]
- Matsuda, N.; Odawara, A.; Okamura, A.; Kinoshita, K.; Shirakawa, T.; Suzuki, I. Deep learning on MEA data obtained from human iPS cell-derived neurons for predicting seizure risk of new drugs. J. Pharm. Toxicol. Methods 2019, 99, 106595. [Google Scholar] [CrossRef] [PubMed]
- Minoshima, W.; Ito, H.; Kudoh, S.N. The glucose concentration-dependent spontaneous electrical activity in a cultured neuronal network. Electr. Commun. Jpn. 2014, 97, 35–41. [Google Scholar] [CrossRef]
- Bateup, H.S.; Johnson, C.A.; Denefrio, C.L.; Saulnier, J.L.; Kornacker, K.; Sabatini, B.L. Excitatory/inhibitory synaptic imbalance leads to hippocampal hyperexcitability in mouse models of tuberous sclerosis. Neuron 2013, 78, 510–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.A.; Sombati, S.; DeLorenzo, R.J. Glutamate injury-induced epileptogenesis in hippocampal neurons: An in vitro model of stroke-induced “epilepsy”. Stroke 2001, 32, 2344–2350. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Zhu, H.; Zhao, Y.; Lang, Y.; Sun, H.; Han, J.; Wang, L.; Wang, C.; Zhou, J. The effect of acute glutamate treatment on the functional connectivity and network topology of cortical cultures. Med. Eng. Phys. 2019, 71, 91–97. [Google Scholar] [CrossRef]
- Schammel, A.; Gorren, C.F.A.; Schmidt, K.; Pfeiffer, S.; Mayer, B. S-nitrosation of glutathione by nitric oxide, peroxynitrite, and •NO/O2•−. Free Radic. Biol. Med. 2003, 34, 1078–1088. [Google Scholar] [CrossRef]
- Brown, C.G. Nitric oxide and neuronal death. Nitric Oxide 2010, 23, 153–165. [Google Scholar] [CrossRef]
- Szab, C. Multiple pathways of peroxynitrite cytotoxicity. Txicol. Lett. 2003, 140–141, 105–112. [Google Scholar] [CrossRef]
- Tricoire, L.; Vitalis, T. Neuronal nitric oxide synthase expressing neurons: A journey from birth to neuronal circuits. Front. Neural Circuits 2012, 6. [Google Scholar] [CrossRef] [Green Version]
- Bouvier, M.; Szatkowski, M.; Amato, A.; Attwell, D. The glial cell glutamate uptake carrier countertransports pH-changing anions. Nature 1992, 360, 471–474. [Google Scholar] [CrossRef]
- Danbolt, N.C. Glutamate uptake. Prog. Neurobiol. 2001, 65, 1–105. [Google Scholar] [CrossRef]
- Tamiya, E.; Sugiura, Y.; Takeuchi, T.; Suzuki, M.; Karube, I.; Akiyama, A. Ultra micro glutamate sensor using platinized carbon-fiber electrode and integrated counter electrode. Sens. Actuators B 1993, 10, 179–184. [Google Scholar] [CrossRef]
- Zilkha, E.; Obrenovitch, T.P.; Koshy, A.; Kusakabe, H.; Bennetto, H.P. Extracellular glutamate: On-line monitoring using microdialysis coupled to enzyme-amperometric analysis. J. Neurosci. Methods 1995, 60, 1–9. [Google Scholar] [CrossRef]
- Kasai, N.; Jimbo, Y.; Torimitsu, K. Electrochemical monitoring of glutamate release at multiple positions in a rat hippocampal slice. Anal. Sci. 2002, 18, 1325–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namiki, S.; Sakamoto, H.; Iinuma, S.; Iino, M.; Hirose, K. Optical glutamate sensor for spatiotemporal analysis of synaptic transmission. Eur. J. Neurosci. 2007, 25, 2249–2259. [Google Scholar] [CrossRef] [PubMed]
- Okubo, Y.; Sekiya, H.; Namiki, S.; Sakamoto, H.; Iinuma, S.; Yamasaki, M.; Watanabe, M.; Hirose, K.; Iino, M. Imaging extrasynaptic glutamate dynamics in the brain. Proc. Natl. Acad. Sci. USA 2010, 107, 6526–6531. [Google Scholar] [CrossRef] [Green Version]
- Schoenheimer, R.; Rittenberg, D. The application of isotopes to the study of intermediary metabolism. Science 1938, 87, 221–226. [Google Scholar] [CrossRef]
- Foster, A.B. Deuterium isotope effects in studies of drug metabolism. Trends Pharmacol. Sci. 1984, 5, 524–527. [Google Scholar] [CrossRef]
- Matsumoto, F.; Makino, K.; Maeda, K.; Patzelt, H.; Maéda, Y.; Fujiwara, S. Conformational changes of troponin C within the thin filaments detected by neutron scattering. J. Mol. Biol. 2004, 342, 1209–1221. [Google Scholar] [CrossRef]
- Synpson, T.J. Applications of multinuclear NMR to structural and biosynthetic studies of polyketide microbial metabolites. Chem. Soc. Rev. 1987, 16, 123–160. [Google Scholar] [CrossRef]
- Ohyama, K.; Suzuki, M.; Kikuchi, J.; Saito, K.; Muranaka, T. Dual biosynthetic pathways to phytosterol via cycloartenol and lanosterol in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.A.; Parker, S.J.; Fiske, B.P.; McCloskey, D.; Gui, D.Y.; Green, C.R.; Vokes, N.I.; Feist, A.M.; Vander Heiden, M.G.; Metallo, C.M. Tracing compartmentalized NADPH metabolism in the cytosol and mitochondria of mammalian cells. Mol. Cell 2014, 55, 253–263. [Google Scholar] [CrossRef] [Green Version]
- van Manen, H.J.; Lenferink, A.; Otto, C. Noninvasive imaging of protein metabolic labeling in single human cells using stable isotopes and Raman microscopy. Anal. Chem. 2008, 80, 9576–9582. [Google Scholar] [CrossRef]
- Vanden-Hehir, S.; Cairns, S.A.; Lee, M.; Zoupi, L.; Shaver, M.P.; Brunton, V.G.; Williams, A.; Hulme, A.N. Alkyne-tagged PLGA allows direct visualization of nanoparticles in vitro and ex vivo by stimulated raman scattering microscopy. Biomacromorecules 2019, 20, 4008–4014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xu, J.; Kong, L.; Liu, T.; Yi, L.; Wang, H.; Huang, W.E.; Zheng, C. Raman-deuterium isotope probing to study metabolic activities of single bacterial cells in human intestinal microbiota. Microb. Biotechnol. 2020, 13, 572–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palonpon, A.F.; Ando, J.; Yamakoshi, H.; Dodo, K.; Sodeoka, M.; Kawata, S.; Fujita, K. Raman and SERS microscopy for molecular imaging of live cells. Nat. Protoc. 2013, 8, 677–692. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, Y.; Luo, W.; Wang, Z.; Zeng, S. Detection of bursts in neuronal spike trains by the mean inter-spike interval method. Prog. Nat. Sci. 2009, 19, 229–235. [Google Scholar] [CrossRef]
- Sunder, S.; Mendelsohn, R.; Bernstein, H.J. Raman studies of the C-H and C-D stretching regions in stearic acid and some specifically deuterated derivatives. Chem. Phys. Lipids 1976, 17, 456–465. [Google Scholar] [CrossRef]
- Cheng, J.X.; Xie, X.S. Vibrational spectroscopic imaging of living systems: An emerging platform for biology and medicine. Science 2015, 350. [Google Scholar] [CrossRef]
- Zhao, Z.; Shen, Y.; Hu, F.; Min, W. Applications of vibrational tags in biological imaging by Raman microscopy. Analyst 2017, 142, 4018–4029. [Google Scholar] [CrossRef]
- Miao, K.; Wei, L. Live-Cell Imaging and Quantification of PolyQ Aggregates by Stimulated Raman Scattering of Selective Deuterium Labeling. ACS Cent. Sci. 2020, 6, 478–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manev, H.; Favaron, M.; Guidotti, A.; Costa, E. Delayed increase of Ca2+ influx elicited by glutamate: Role in neuronal death. Mol. Pharm. 1989, 36, 106–112. [Google Scholar]
- Jaiswal, M.K.; Zech, W.D.; Goos, M.; Leutbecher, C.; Ferri, A.; Zippelius, A.; Carrì, M.T.; Nau, R.; Keller, B.U. Impairment of mitochondrial calcium handling in a mtSOD1 cell culture model of motoneuron disease. BMC Neurosci. 2009, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sourial-Bassillious, N.; Rydelius, P.A.; Aperia, A.; Aizman, O. Glutamate-mediated calcium signaling: A potential target for lithium action. Neuroscience 2009, 161, 1126–1134. [Google Scholar] [CrossRef] [Green Version]
- Reinis, S.; Lahue, R.H., Jr.; Weiss, D.S.; Money, K.E.; Landolt, J.P. Deuterium oxide modification of neuronal characteristics in the visual cortex of the cat. Neurosci. Lett. 1983, 38, 269–274. [Google Scholar] [CrossRef]
- Krahe, R.; Gabbiani, F. Burst firing in sensory systems. Nat. Rev. Neurosci. 2004, 5, 13–23. [Google Scholar] [CrossRef]
- Mayer, M.L.; Miller, R.J. Excitatory amino acid receptors, second messengers and regulation of intracellular Ca2+ in mammalian neurons. Trends Pharm. Sci. 1990, 11, 254–260. [Google Scholar] [CrossRef]
- Nakanishi, S. Molecular diversity of glutamate receptors and implications for brain function. Science 1992, 258, 597–603. [Google Scholar] [CrossRef]
- Hascup, K.N.; Hascup, E.R.; Stephens, M.L.; Glaser, P.E.; Yoshitake, T.; Mathé, A.A.; Gerhardt, G.A.; Kehr, J. Resting glutamate levels and rapid glutamate transients in the prefrontal cortex of the Flinders Sensitive Line rat: A genetic rodent model of depression. Neuropsychopharmacology 2011, 36, 1769–1777. [Google Scholar] [CrossRef]
- Choi, Y.M.; Kim, S.H.; Uhm, D.Y.; Park, M.K. Glutamate-mediated [Ca2+]c dynamics in spontaneously firing dopamine neurons of the rat substantia nigra pars compacta. J. Cell Sci. 2003, 116, 2665–2675. [Google Scholar] [CrossRef] [Green Version]
- Obrietan, K.; Van den Pol, A.N. Calcium hyperexcitability in neurons cultured with glutamate receptor blockade. J. Neurophysiol. 1995, 73, 1524–1536. [Google Scholar] [CrossRef]
- Leininger, E.; Belousov, A.B. Recovery of network-driven glutamatergic activity in rat hippocampal neurons during chronic glutamate receptor blockade. Brain Res. 2009, 1251, 87–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brickley, S.G.; Farrant, M.; Swanson, G.T.; Cull-Candy, S.G. CNQX increases GABA-mediated synaptic transmission in the cerebellum by an AMPA/kainate receptor-independent mechanism. Neuropharmacology 2001, 41, 730–736. [Google Scholar] [CrossRef]
- Clements, J.D.; Lester, R.A.; Tong, G.; Jahr, C.E.; Westbrook, G.L. The time course of glutamate in the synaptic cleft. Science 1992, 258, 1498–1501. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.W.; Maulucci-Gedde, M.; Kriegstein, A.R. Glutamate neurotoxicity in cortical cell culture. J. Neurosci. 1987, 7, 357–368. [Google Scholar] [CrossRef]
- Chen, Q.; Harris, C.; Brown, C.S.; Howe, A.; Surmeier, D.J.; Reiner, A. Glutamate-mediated excitotoxic death of cultured striatal neurons is mediated by non-NMDA receptors. Exp. Neurol. 1995, 136, 212–224. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture No. | Before GLU Application (spikes/s) | Before GLU-D Application (spikes/s) |
|---|---|---|
| Culture No.1 | 36.7 ± 24.2 | 16.9 ± 12.8 |
| Culture No.2 | 14.6 ± 9.0 | 2.3 ± 3.0 |
| Culture No.3 | 11.7 ± 12.2 | 8.0 ± 5.8 |
| Culture No.4 | 10.2 ± 6.7 | 16.3 ± 12.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minoshima, W.; Masui, K.; Tani, T.; Nawa, Y.; Fujita, S.; Ishitobi, H.; Hosokawa, C.; Inouye, Y. Deuterated Glutamate-Mediated Neuronal Activity on Micro-Electrode Arrays. Micromachines 2020, 11, 830. https://doi.org/10.3390/mi11090830
Minoshima W, Masui K, Tani T, Nawa Y, Fujita S, Ishitobi H, Hosokawa C, Inouye Y. Deuterated Glutamate-Mediated Neuronal Activity on Micro-Electrode Arrays. Micromachines. 2020; 11(9):830. https://doi.org/10.3390/mi11090830
Chicago/Turabian StyleMinoshima, Wataru, Kyoko Masui, Tomomi Tani, Yasunori Nawa, Satoshi Fujita, Hidekazu Ishitobi, Chie Hosokawa, and Yasushi Inouye. 2020. "Deuterated Glutamate-Mediated Neuronal Activity on Micro-Electrode Arrays" Micromachines 11, no. 9: 830. https://doi.org/10.3390/mi11090830
APA StyleMinoshima, W., Masui, K., Tani, T., Nawa, Y., Fujita, S., Ishitobi, H., Hosokawa, C., & Inouye, Y. (2020). Deuterated Glutamate-Mediated Neuronal Activity on Micro-Electrode Arrays. Micromachines, 11(9), 830. https://doi.org/10.3390/mi11090830