


Beyond Memristors: Neuromorphic Computing Using Meminductors

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
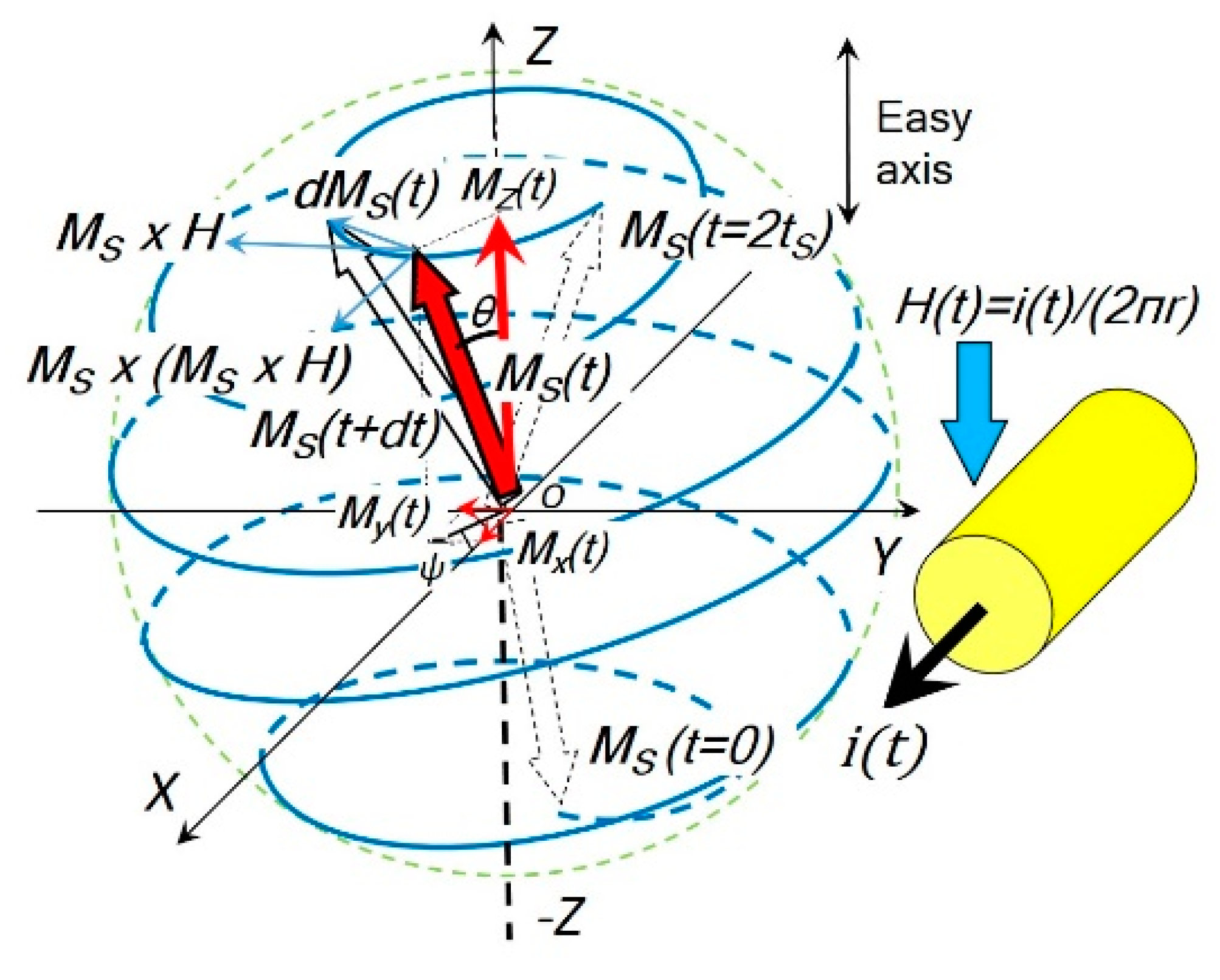
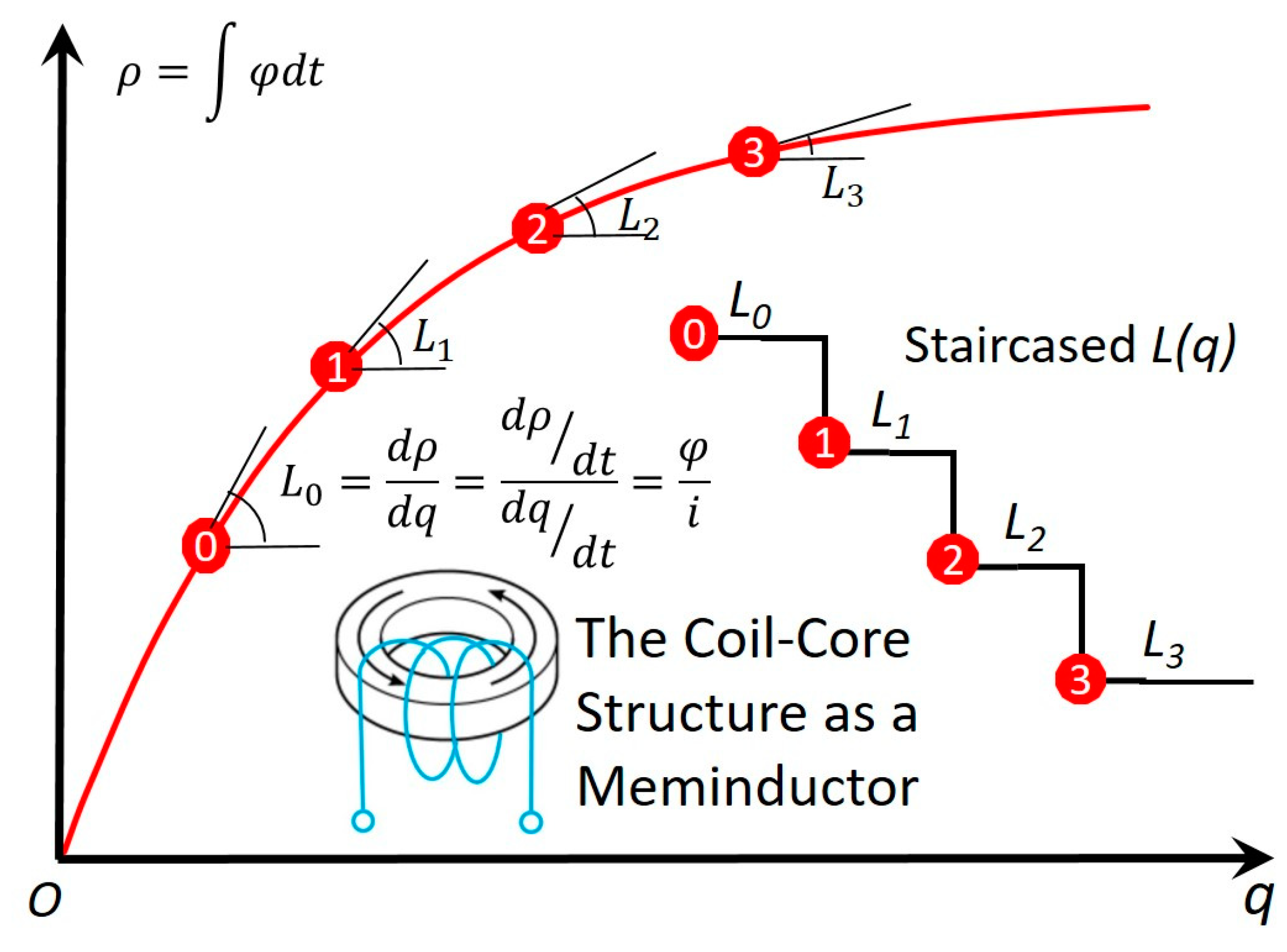
2. LLG Model for the Coil Core Structure
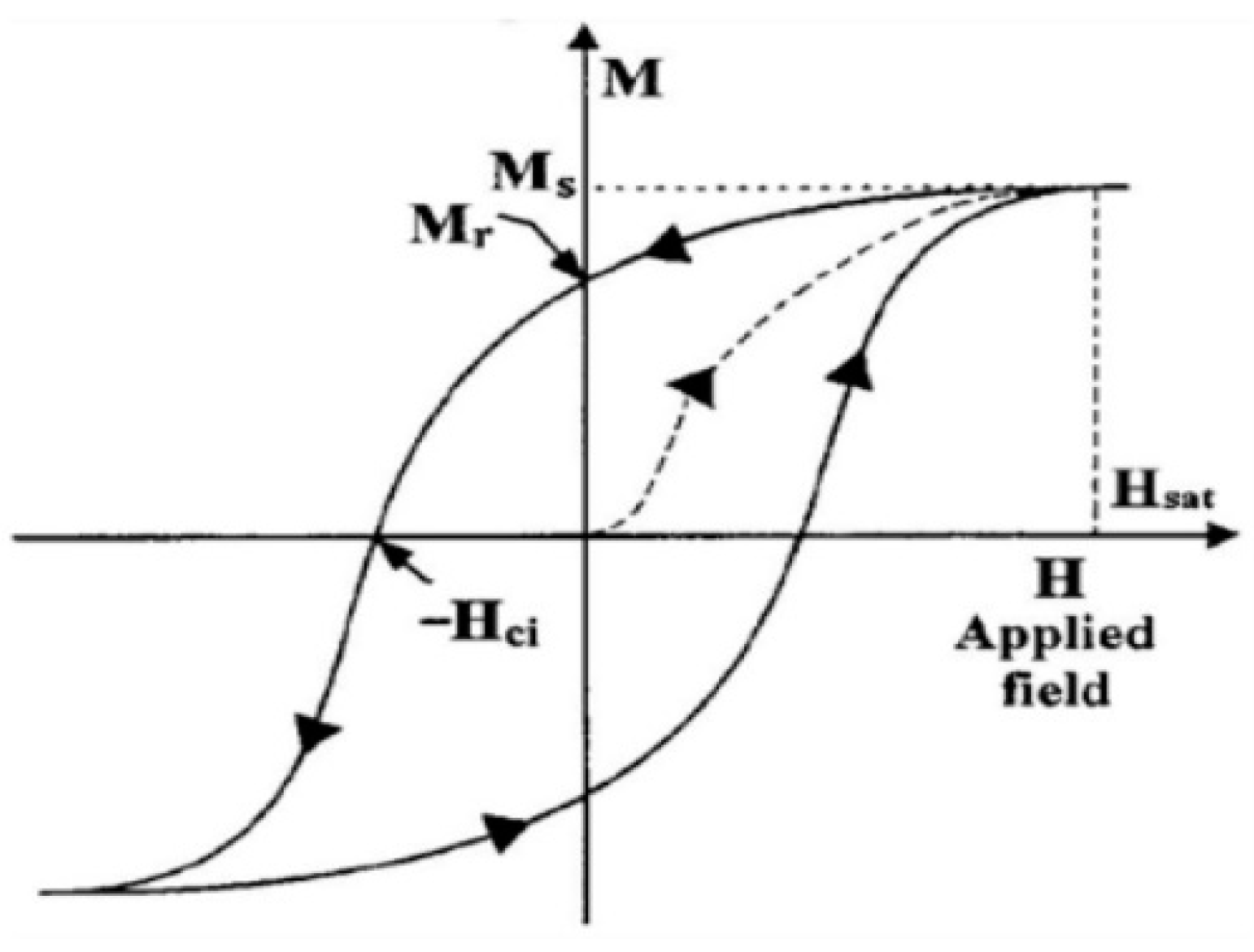
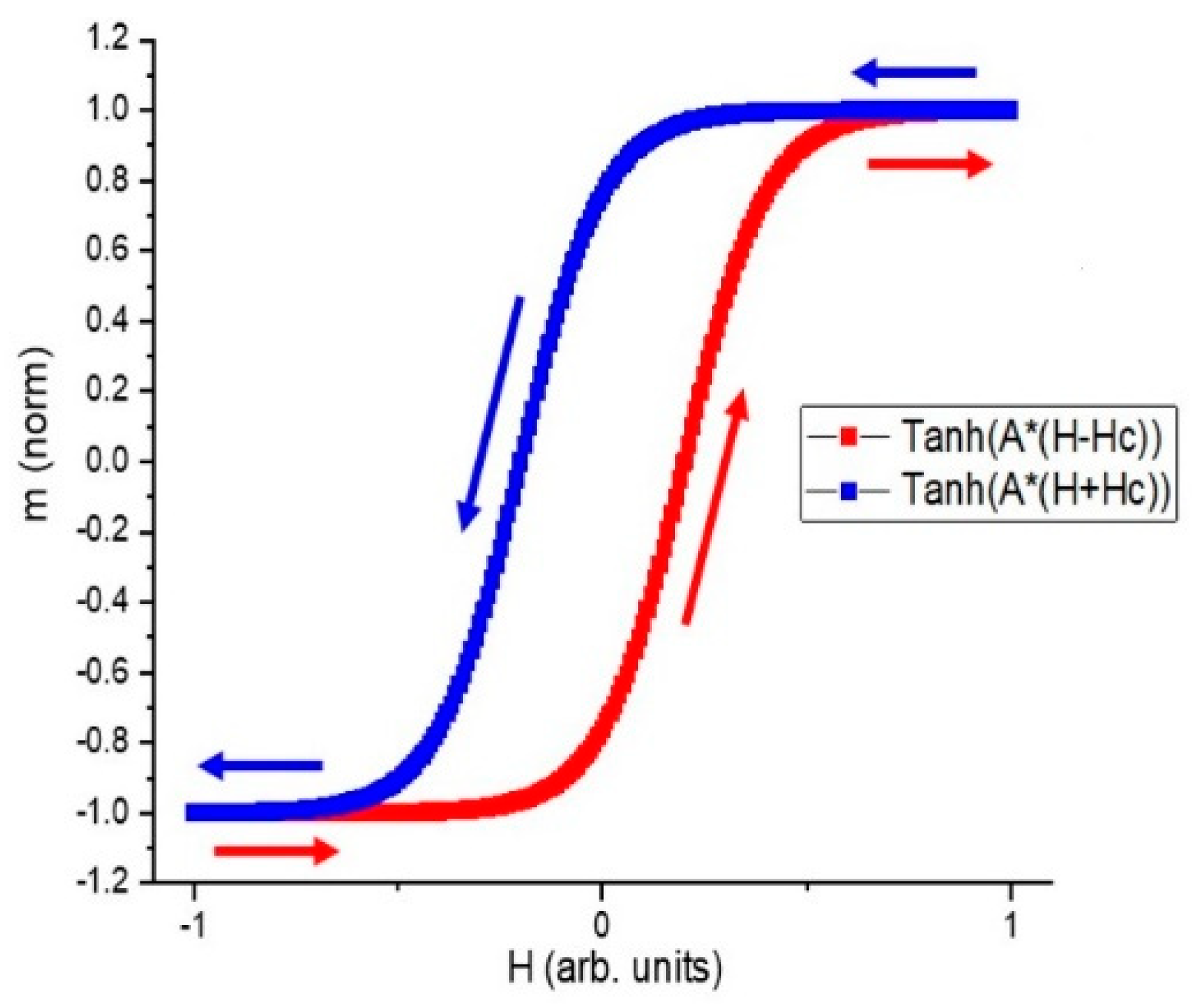
3. Experimental Verification of the Rotational Model
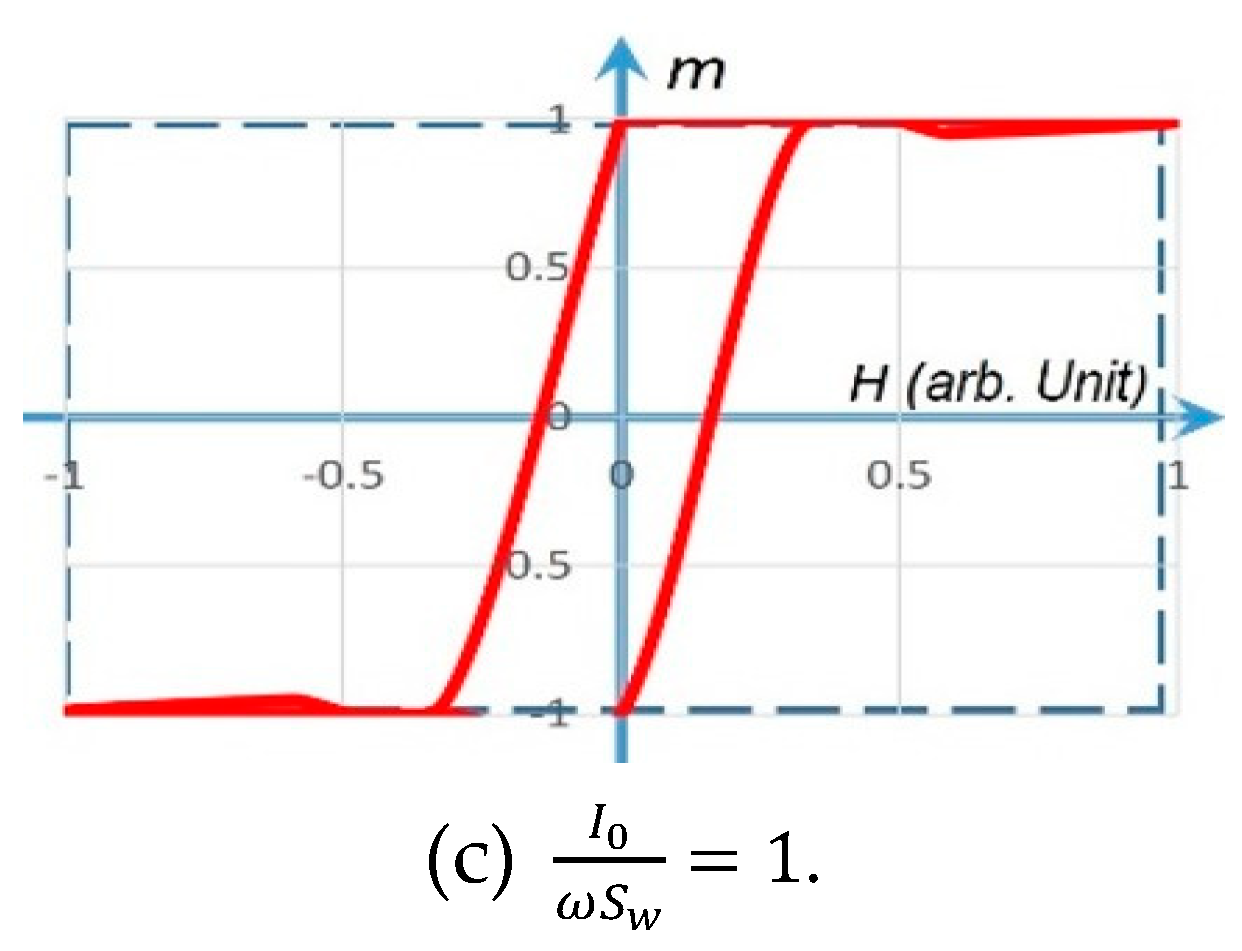
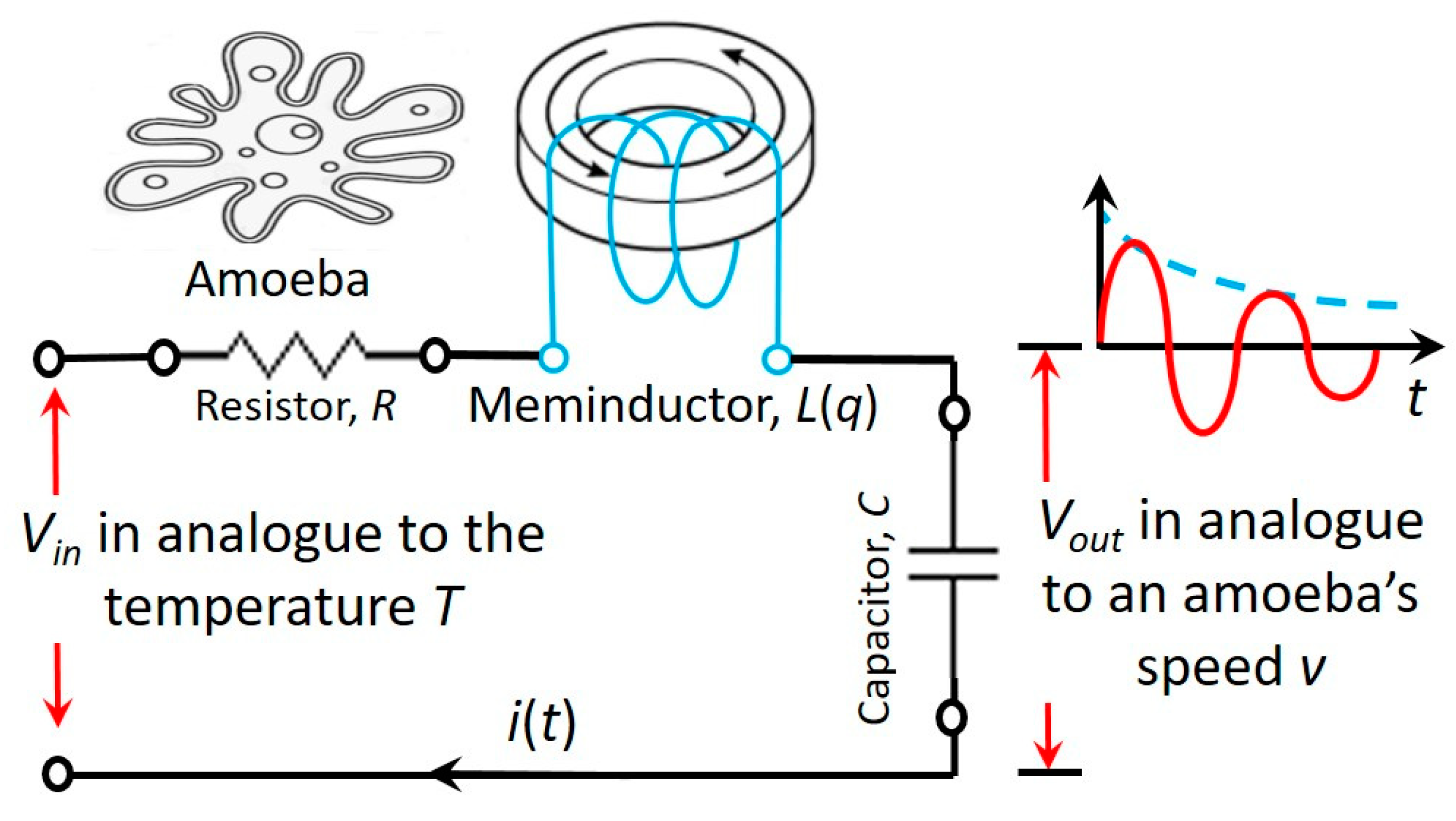
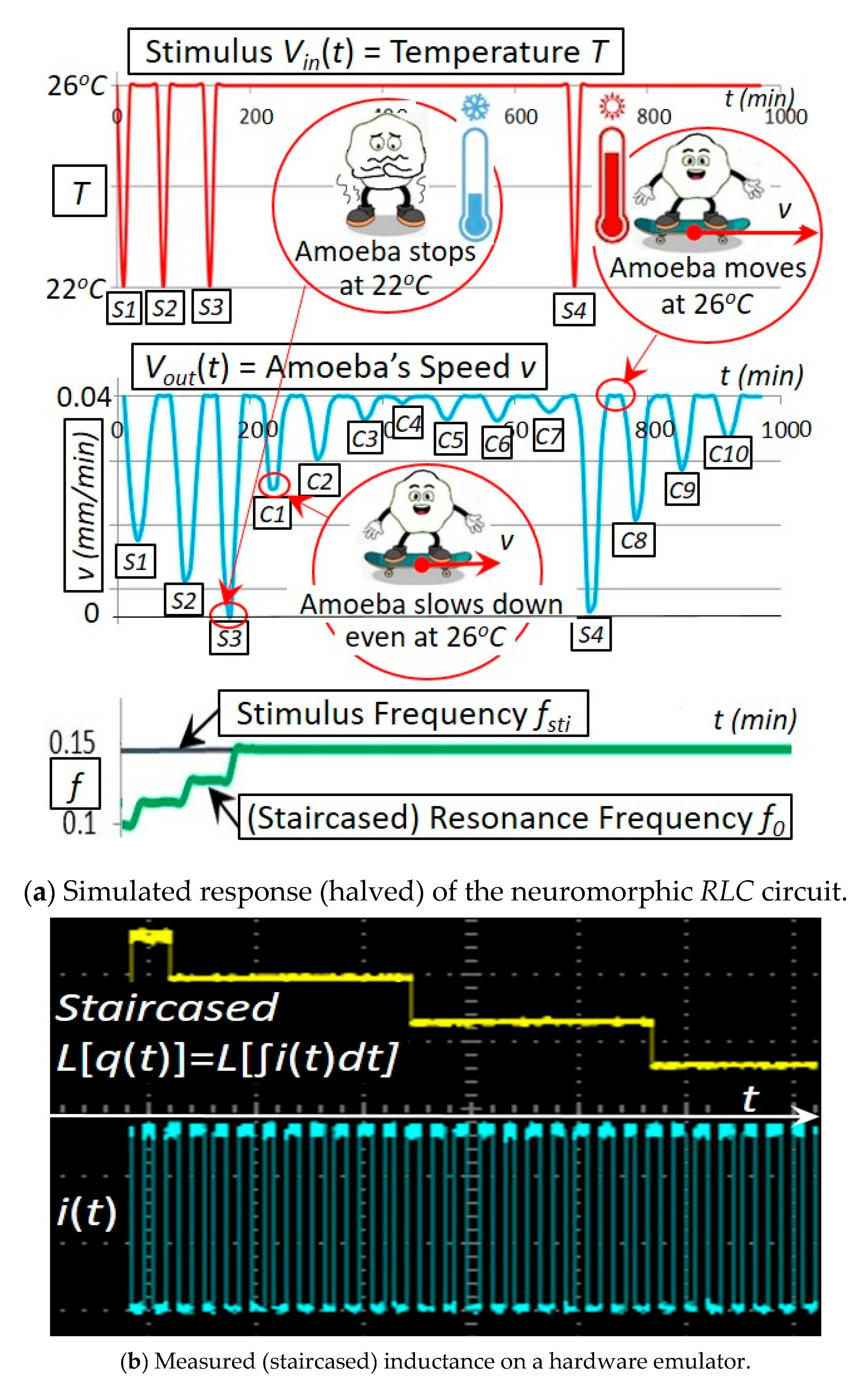
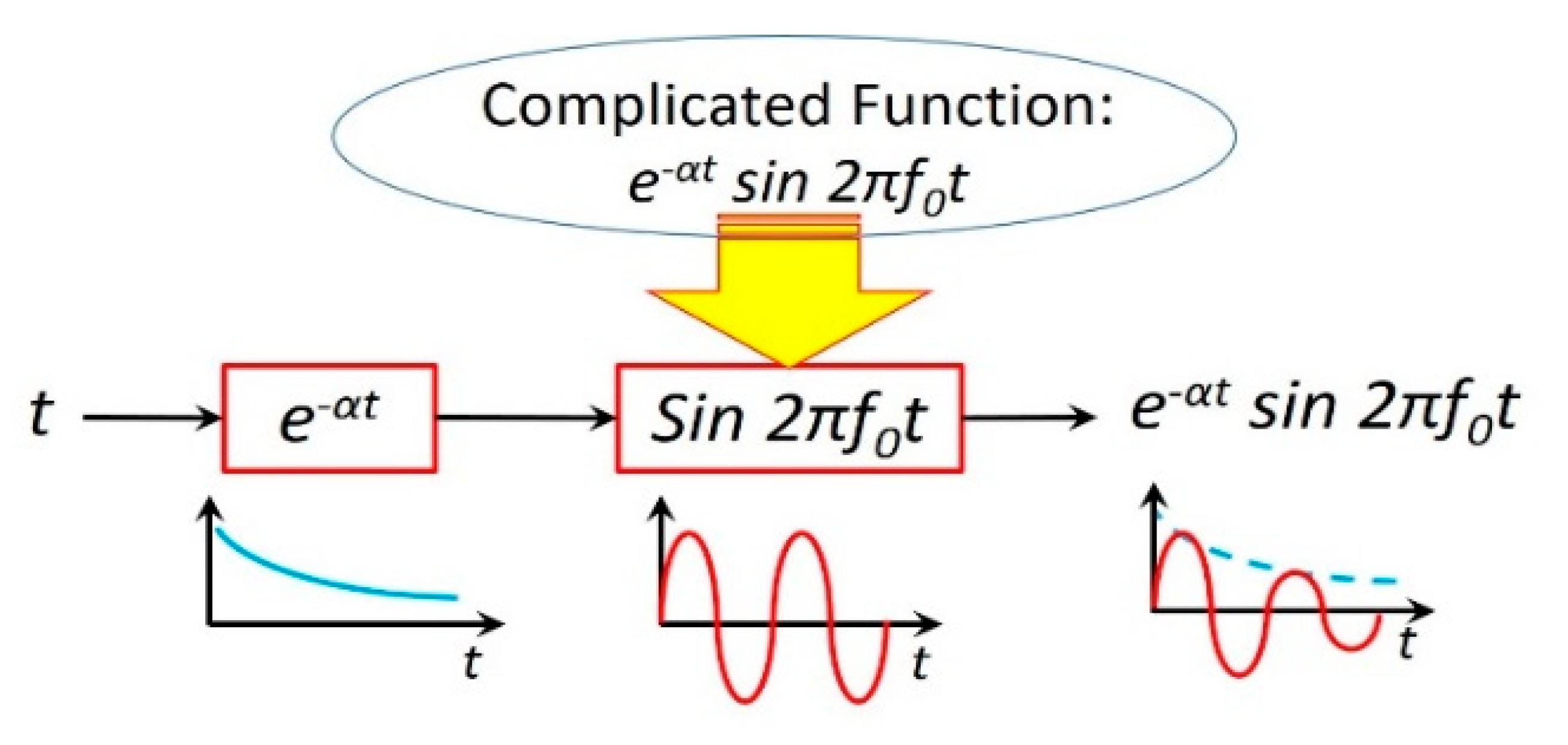
4. Simulations and Experiments of a Coil Core Meminductor for Neuromorphic Computing
5. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Alexander, C.; Sadiku, M. Fundamentals of Electric Circuits, 3rd ed.; McGraw-Hill: New York, NY, USA, 1994; p. 211. [Google Scholar]
- Pershin, Y.V.; La Fontaine, S.; Di Ventra, M. Memristive model of amoeba learning. Phys. Rev. E 2009, 80, 021926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donahue, M.J.; Porter, D.G. Analysis of switching in uniformly magnetized bodies. IEEE Trans. Magn. 2002, 38, 2468–2470. [Google Scholar] [CrossRef]
- Gyorgy, E.M. Rotational model of flux reversal in square loop ferritcs. J. Appl. Phys. 1957, 28, 1011–1015. [Google Scholar] [CrossRef]
- Landau, L.D.; Lifshitz, E.M. Theory of the dispersion of magnetic permeability in ferromagnetic bodies. Phys. Z. Sowjetunion. 1935, 8, 153. [Google Scholar]
- Gilbert, T.L. A Lagrangian formulation of the gyromagnetic equation of the magnetic field. Phys. Rev. 1955, 100, 1243. [Google Scholar]
- Cushman, N. Characterization of Magnetic Switch Cores. IRE Trans. Compon. Parts 1961, 8, 45–50. [Google Scholar] [CrossRef]
- Menyuk, N.; Goodenough, J. Magnetic materials for digital computer components I. J. Appl. Phys. 1955, 26, 8–18. [Google Scholar] [CrossRef]
- Riaza, R. Second order mem-circuits. Int. J. Circuit Theory Appl. 2015, 43, 1719–1742. Available online: https://mc.manuscriptcentral.com/ijcta (accessed on 8 January 2023). [CrossRef] [Green Version]
- Chua, L. Memristor—The Missing Circuit Element. IEEE Trans. Circuit Theory 1971, CT-18, 507–519. [Google Scholar] [CrossRef]
- Georgiou, P.S.; Barahona, M.; Yaliraki, S.N.; Drakakis, E.M. On memristor ideality and reciprocity. Microelectron. J. 2014, 45, 1363–1371. [Google Scholar] [CrossRef] [Green Version]
- Rudowicz, C.; Sung, H.W.F. Textbook treatments of the hysteresis loop for ferromagnets-Survey of misconceptions and misinterpretations. Am. J. Phys. 2003, 71, 1080. [Google Scholar]
- Pershin, Y.; Ventra, M.D. Experimental demonstration of associative memory with memristive neural networks. Nat. Preced. 2009. [Google Scholar] [CrossRef]
- Pershin, Y.; Ventra, M.D.; Chua, L. Circuit Elements with Memory: Memristors, Memcapacitors, and Meminductors. Proc. IEEE 2009, 97, 1717–1724. [Google Scholar]
- Wang, F.Z.; Chua, L.O.; Yang, X.; Helian, N.; Tetzlaff, R.; Schmidt, T.; Li, L.; Carrasco, J.M.; Chen, W.; Chu, D. Adaptive Neuromorphic Architecture (ANA). Neural Netw. 2013, 45, 111–116. [Google Scholar] [CrossRef] [PubMed]
- LeCun, Y.; Bengio, Y.; Hinton, G. Deep Learning. Nature 2015, 521, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Hodgkin, A.; Huxley, A. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar] [CrossRef] [PubMed]
- Chua, L.; Sbitnev, V.; Kim, H. Hodgkin–Huxley axon is made of memristors. Int. J. Bifurc. Chaos 2012, 22, 1230011. [Google Scholar] [CrossRef]
- Chua, L.; Sbitnev, V.; Kim, H. Neurons Are Poised Near The Edge Of Chaos. Int. J. Bifurc. Chaos 2012, 22, 1250098. [Google Scholar] [CrossRef]
- Sah, M.; Kim, H.; Chua, L. Brains Are Made of Memristors. IEEE Circuits Syst. Mag. 2014, 14, 12–36. [Google Scholar] [CrossRef]
- Chaisson, E.J. Cosmic Evolution—Biological, Havard University Course Syllabus, version 7; NIH The National Center for Biotechnology Information: Bethesda, MD, USA, 2012. [Google Scholar]
- Wang, F.Z.; Helian, N.; Wu, S.; Yang, X.; Guo, Y.; Lim, G.; Rashid, M.M. Delayed switching applied to memristor neural networks. J. Appl. Phys. 2012, 111, 07E317. [Google Scholar] [CrossRef]
- Wang, F.Z.; Helian, N.; Wu, S.; Lim, M.G.; Guo, Y.; Parker, M.A. Delayed Switching in Memristors and Memristive Systems. IEEE Electron Device Lett. 2010, 31, 755–757. [Google Scholar] [CrossRef] [Green Version]
- Turing, A.M. On Computable Numbers, with an Application to the Entscheidungs problem. Proc. Lond. Math. Soc. 1936, 42, 230–265. [Google Scholar]
- Post, E. Recursive Unsolvability of a Problem of Thue. J. Symb. Log. 1947, 12, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Turing, A.M. Intelligent Machinery; University Park Press: Baltimore, UK, 1968; p. 31. [Google Scholar]
- Hennie, F.C.; Stearns, F.C. Two-tape simulation of multitape Turing machines. JACM 1966, 13, 533–546. [Google Scholar] [CrossRef]
- von Neumann, J. First Draft of a Report on the EDVAC (PDF), 1945. Available online: http://abelgo.cn/cs101/papers/Neumann.pdf (accessed on 8 January 2023).
- Markgraf, J.D. The Von Neumann Bottleneck. Available online: https://web.archive.org/web/20131212205159/http://aws.linnbenton.edu/cs271c/markgrj/ (accessed on 8 January 2023).
- MFTL (My Favorite Toy Language) Entry Jargon File 4.4.7. Available online: http://catb.org/jargon/html/ (accessed on 8 January 2023).
- Copeland, B. (Ed.) The Essential Turing: Seminal Writings in Computing, Logic, Philosophy, Artificial Intelligence, and Artificial Life plus The Secrets of Enigma; Clarendon Press (Oxford University Press): Oxford, UK, 2004. [Google Scholar]
- Encephalo-Etymology Online Etymology Dictionary. Available online: https://www.etymonline.com/ (accessed on 8 January 2023).
- Parent, A.; Carpenter, M.B. Ch. 1. Carpenter’s Human Neuroanatomy; Williams & Wilkins: Baltimore, MD, USA, 1995; ISBN 978-0-683-06752-1. [Google Scholar]
- Bigos, K.L.; Hariri, A.; Weinberger, D. Neuroimaging Genetics: Principles and Practices; Oxford University Press: Oxford, UK, 2015; p. 157. ISBN 978-0-19-992022-8. [Google Scholar]
- Cosgrove, K.P.; Mazure, C.M.; Staley, J.K. Evolving knowledge of sex differences in brain structure, function, and chemistry. Biol. Psychiatry 2007, 62, 847–855. [Google Scholar] [CrossRef] [Green Version]
- Molina, D. Kimberley; DiMaio, Vincent J.M. Normal Organ Weights in Men. Am. J. Forensic Med. Pathol. 2012, 33, 368–372. [Google Scholar] [CrossRef]
- “Amoeba” Archived at the Wayback Machine. Available online: Oxforddictionaries.com (accessed on 8 January 2023).
- Singleton, P. Dictionary of Microbiology and Molecular Biology, 3rd ed.; John Wiley & Sons: Chichester, UK, 2006; p. 32. ISBN 978-0-470-03545-0. [Google Scholar]
- Patterson, D.J. Amoebae: Protists Which Move and Feed Using Pseudopodia. Tree of Life Web Project. Available online: http://tolweb.org/accessory/Amoebae?acc_id=51 (accessed on 8 January 2023).
- “The Amoebae” The University of Edinburgh. Available online: https://maciverlab.bms.ed.ac.uk/amoebae.htm (accessed on 8 January 2023).
- van Egmond, W. Sun Animalcules and Amoebas. Microscopy-UK. Available online: http://www.microscopy-uk.org.uk/mag/indexmag.html?http://www.microscopy-uk.org.uk/mag/wimsmall/sundr.html (accessed on 8 January 2023).
- Nelson, M.E. Electrophysiological Models. In Databasing the Brain: From Data to Knowledge; Koslow, S., Subramaniam, S., Eds.; Wiley: New York, NY, USA, 2005; pp. 285–301. [Google Scholar]
- Gray, D.J.; Wu, S.M. Foundations of Cellular Neurophysiology, 3rd ed.; MIT Press: Cambridge, MA, USA, 1997; ISBN 978-0-262-10053-3. [Google Scholar]
- Krapivin, V.F.; Varotsos, C.A.; Soldatov, V.Y. New Ecoinformatics Tools in Environmental Science: Applications and Decision-Making; Springer: New York, NY, USA, 2015; pp. 37–38. ISBN 9783319139784. [Google Scholar]
- Rakowski, R.F.; Gadsby, D.C.; De Weer, P. Stoichiometry and voltage dependence of the sodium pump in voltage-clamped, internally dialyzed squid giant axon. J. Gen. Physiol. 1989, 93, 903–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer: Sunderland, MA, USA, 2001; ISBN 978-0-87893-321-1. [Google Scholar]
- Porteus, S.D. Primitive Intelligence and Environment. Nature 1938, 142, 774. [Google Scholar] [CrossRef]
- Sample, I. Primitive intelligence, New Scientist, 27 September 2000. Available online: https://www.newscientist.com/article/dn25-primitive-intelligence/ (accessed on 8 January 2023).
- Näätänen, R.; Tervaniemi, M.; Sussman, E.; Paavilainen, P.; Winkler, I. Primitive intelligence in the auditory cortex. Trends Neurosci. 2001, 24, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Kindratenko, V. Novel Computing Architectures. Comput. Sci. Eng. 2009, 11, 54–57. [Google Scholar] [CrossRef]
- Novel Computing Platforms and Information Processing Approaches. Available online: https://csl.illinois.edu/research/impact-areas/health-it/novel-computing-platforms-and-information-processing-approaches (accessed on 6 February 2023).










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.Z. Beyond Memristors: Neuromorphic Computing Using Meminductors. Micromachines 2023, 14, 486. https://doi.org/10.3390/mi14020486
Wang FZ. Beyond Memristors: Neuromorphic Computing Using Meminductors. Micromachines. 2023; 14(2):486. https://doi.org/10.3390/mi14020486
Chicago/Turabian StyleWang, Frank Zhigang. 2023. "Beyond Memristors: Neuromorphic Computing Using Meminductors" Micromachines 14, no. 2: 486. https://doi.org/10.3390/mi14020486
APA StyleWang, F. Z. (2023). Beyond Memristors: Neuromorphic Computing Using Meminductors. Micromachines, 14(2), 486. https://doi.org/10.3390/mi14020486