Electroceutical Approach for Impairing the Motility of Pathogenic Bacterium Using a Microfluidic Platform


Abstract
:1. Introduction
2. Materials and Methods
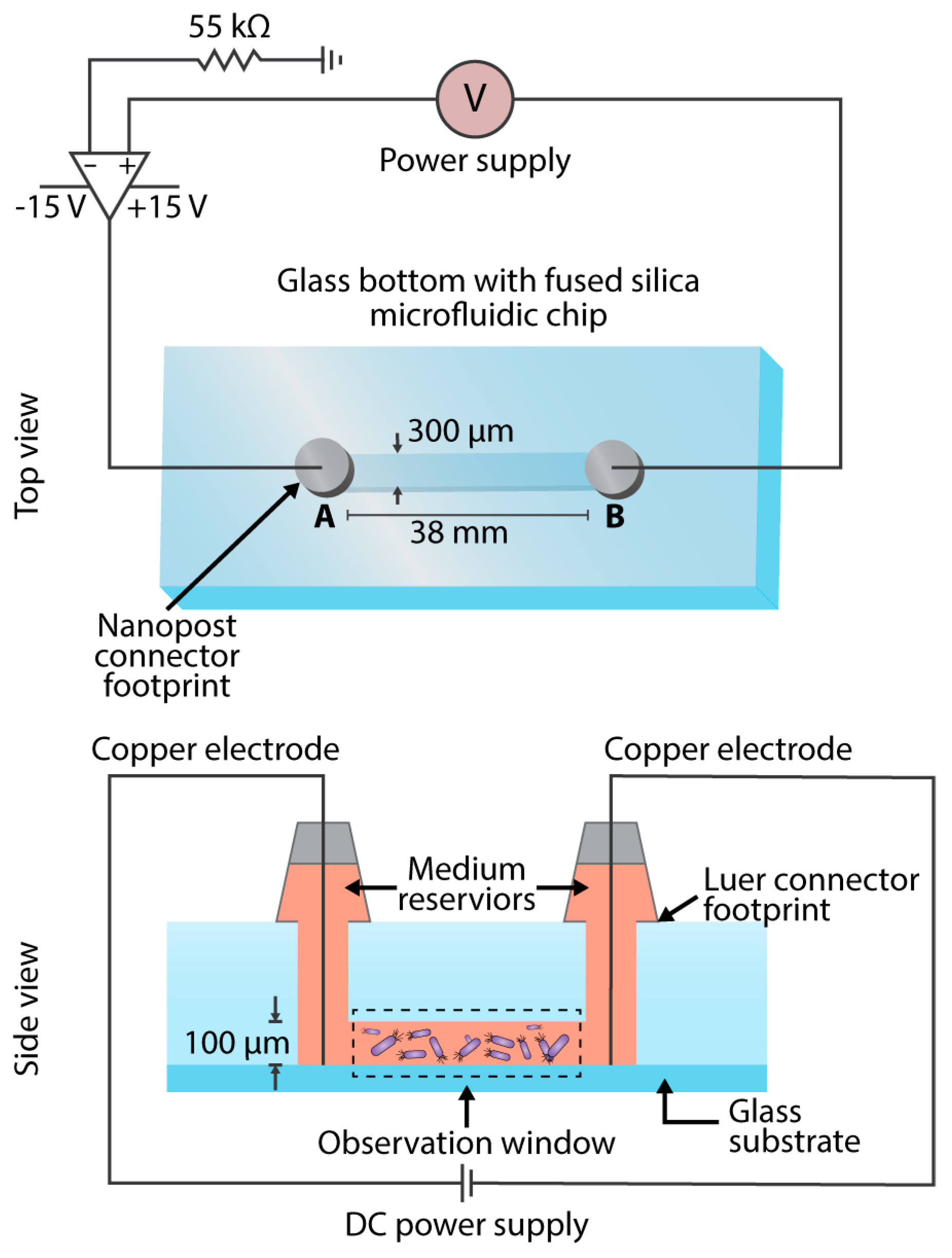
2.1. Experimental Parameter Design
2.2. Cell Culture
2.3. Metrics Acquisition
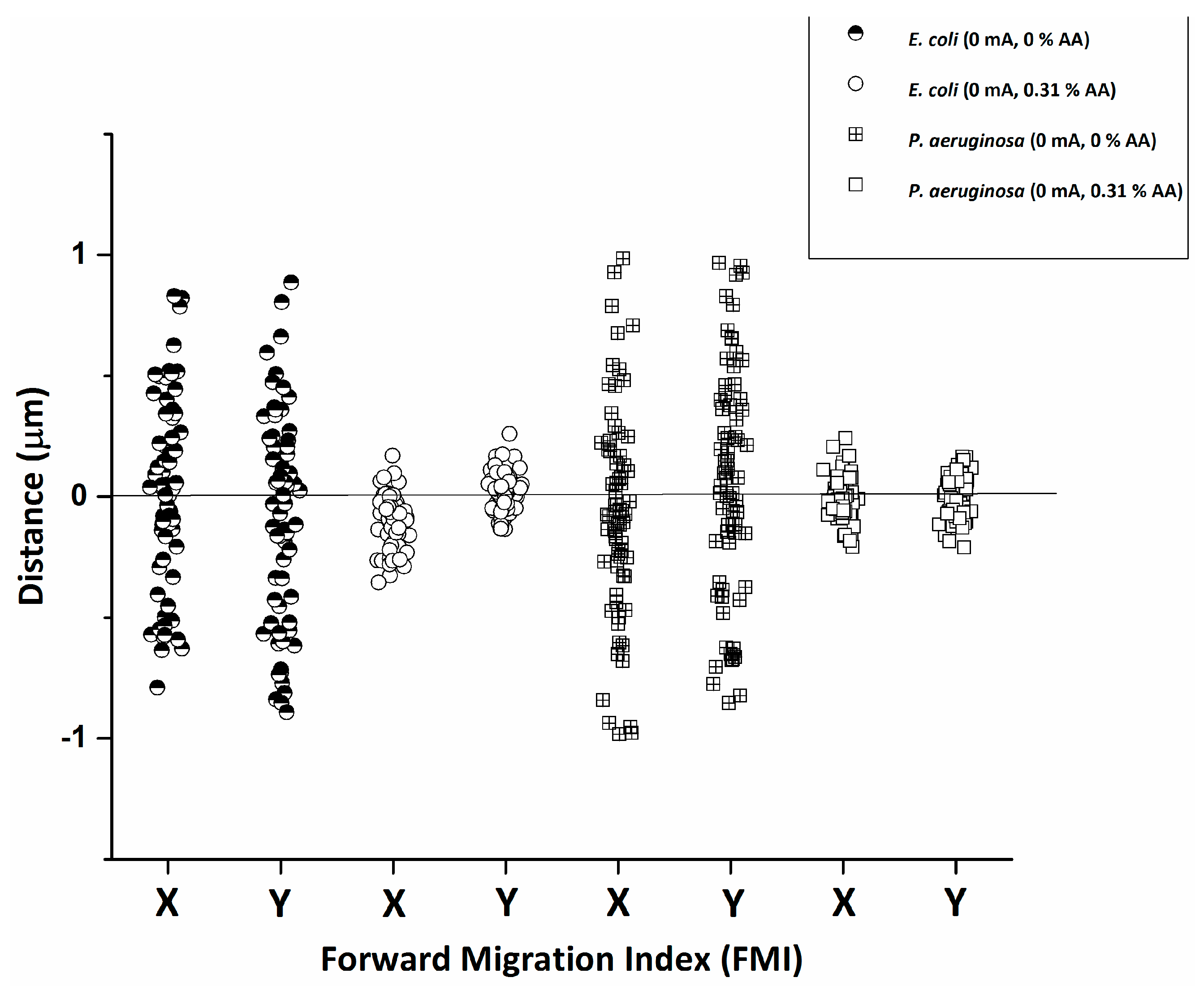
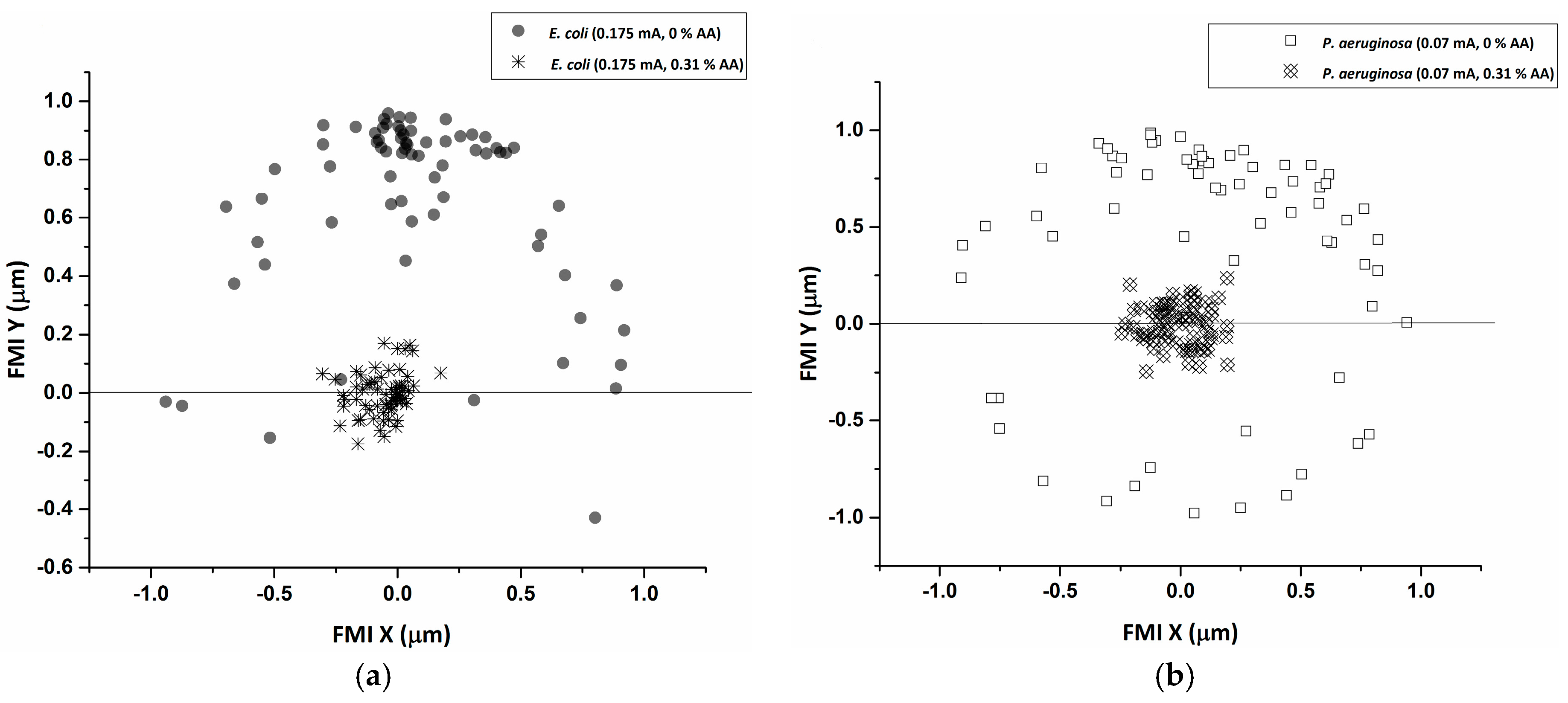
2.4. Statistical Tracking and Data Analysis
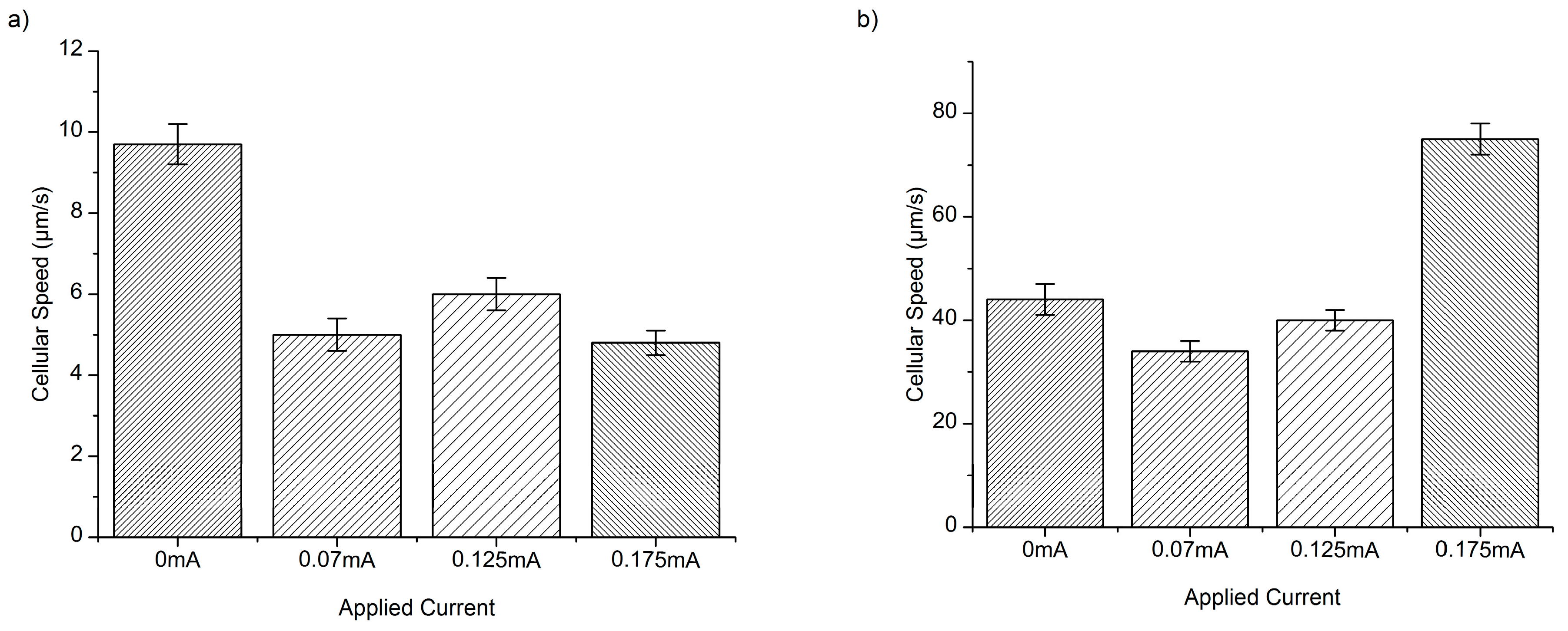
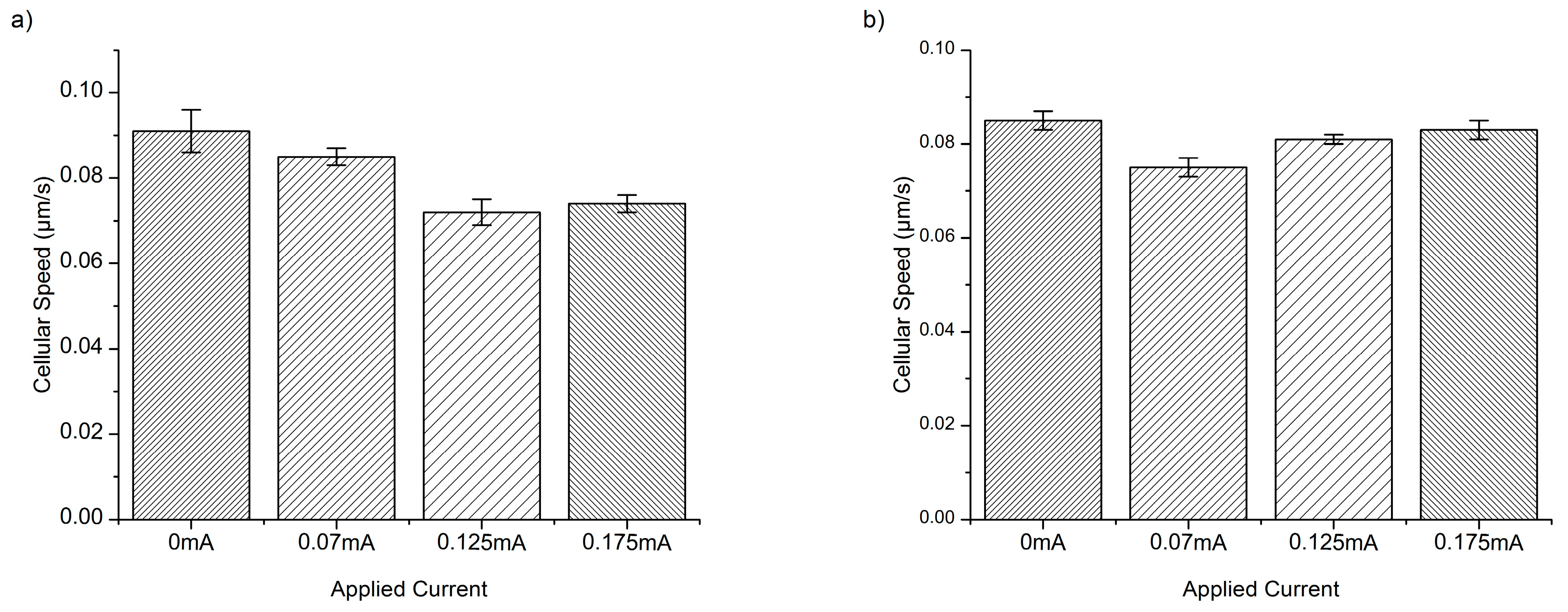
3. Results and Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Centers for Disease Control and Prevention (CDC). Antibiotic Resistance Threats in the United States; Centers for Disease Control and Prevention (CDC): Atlanta, GA, USA, 2013.
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The Calgary biofilm device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar] [PubMed]
- Liu, Z.; Chen, W.; Papadopoulos, K.D. Electrokinetic movement of Escherichia coli in capillaries. Environ. Microbiol. 1999, 1, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kim, P.S.S.; Lee, K.; Kim, J.; Kim, M.J. Galvanotactic behavior of Tetrahymena pyriformis under electric fields. J. Micromech. Microeng. 2013, 23, 1–9. [Google Scholar] [CrossRef]
- Asadi, M.R.; Torkaman, G. Bacterial inhibition by electrical stimulation. Adv. Wound Care 2014, 3, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Rowley, B.A.; McKenna, J.M.; Chase, G.R.; Wolcott, L.E. The influence of electrical current on an infecting microorganism in wounds. Ann. N. Y. Acad. Sci. 1974, 238, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Subramanian, S.; Gerasopoulos, K.; Ben-Yoav, H.; Wu, H.C.; Quan, D.; Carter, K.; Meyer, M.T.; Bentley, W.E.; Ghodssi, R. Effect of electrical energy on the efficacy of biofilm treatment using the bioelectric effect. npj Biofilms Microbiomes 2015, 15016, 1–8. [Google Scholar] [CrossRef]
- Gall, I.; Herzberg, M.; Oren, Y. The effect of electric fields on bacterial attachment to conductive surfaces. Soft Matter 2013, 9, 2443–2452. [Google Scholar] [CrossRef]
- Schmidt-Malan, S.M.; Karau, M.J.; Cede, J.; Greenwood-Quaintance, K.E.; Brinkman, C.L.; Mandrekar, J.N.; Patel, R. Antibiofilm activity of low-amperage continuous and intermittent direct electrical current. Antimicrob. Agents Chemother. 2015, 59, 4610–4615. [Google Scholar] [CrossRef] [PubMed]
- Sandvik, E.L.; McLeod, B.R.; Parker, A.E.; Stewart, P.S. Direct electric current treatment under physiologic saline conditions kills Staphylococcus epidermidis biofilms via electrolytic generation of hypochlorous acid. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lin, F. Microfluidic devices for studying chemotaxis and electrotaxis. Trends Cell Biol. 2011, 21, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Song, B.; Pu, J.; Wada, T.; Reid, B.; Tai, G.; Wang, F.; Guo, A.; Walczysko, P.; Gu, Y.; et al. Electrical signals control wound healing through phosphatidylinositol-3-OH kinase-γ and PTEN. Nature 2006, 442, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Sauer, K.; Camper, A.K.; Ehrlich, G.D.; Costerton, J.W.; Davies, D.G. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm. J. Bacteriol. 2002, 184, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Voegele, P.; Badiola, J.; Schmidt-Malan, S.M.; Karau, M.J.; Greenwood-Quaintance, K.E.; Mandrekar, J.N.; Patel, R. Antibiofilm activity of electrical current in a catheter model. Antimicrob. Agents Chemother. 2016, 60, 1476–1480. [Google Scholar] [CrossRef] [PubMed]
- Halstead, F.D.; Rauf, M.; Moiemen, N.S.; Bamford, A.; Wearn, C.M.; Fraise, A.P.; Lund, P.A.; Oppenheim, B.A.; Webber, M.A. The antibacterial activity of acetic acid against biofilm-producing pathogens of relevance to burns patients. PLoS ONE 2015, 10, e0136190. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.B.D.; Favarin, L.; Luchese, R.H.; McIntosh, D. Psychrotrophic bacteria in milk: How much do we really know? Braz. J. Microbiol. 2015, 46, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Ronen, A.; Duan, W.; Wheeldon, I.; Walker, S.; Jassby, D. Microbial attachment inhibition through low-voltage electrochemical reactions on electrically conducting membranes. Environ. Sci. Technol. 2015, 49, 12741–12750. [Google Scholar] [CrossRef] [PubMed]
- Ghatak, P.D.; Schlanger, R.; Ganesh, K.; Lambert, L.; Gordillo, G.M.; Martinsek, P.; Roy, S. A wireless electroceutical dressing lowers cost of negative pressure wound therapy. Adv. Wound Care 2015, 4, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T.; Alhede, M.; Jensen, P.Ø.; Nielsen, A.K.; Johansen, H.K.; Homøe, P.; Høiby, N.; Givskov, M.; Kirketerp-Møller, K. Antibiofilm properties of acetic acid. Adv. Wound Care 2015, 4, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Watters, C.; Kay, M. Eradication of Wound Biofilms by Electrical Stimulation; Springer: Heidelberg, Germany, 2014; pp. 425–447. [Google Scholar]
- Preiss, L.; Neethirajan, S. Single cells of Pseudomonas aeruginosa exhibit electrotaxis and durotaxis behaviours. bioRxiv 2015. [Google Scholar] [CrossRef]
- Maadi, H.; Haghi, M.; Delshad, R.; Kangarloo, H.; Mohammadnezhady, M.A.; Hemmatyar, G.R. Effect of alternating and direct currents on Pseudomonas aeruginosa growth in vitro. Afr. J. Biotechnol. 2010, 9, 6373–6379. [Google Scholar]
- Chung, S.; Sudo, R.; Mack, P.J.; Wan, C.R.; Vickerman, V.; Kamm, R.D. Cell migration into scaffolds under co-culture conditions in a microfluidic platform. Lab Chip 2009, 9, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, W.; Yuan, B.; Xiao, L.; Liu, D.; Jin, Y.; Quan, B.; Wang, J.O.; Ibrahim, K.; Wang, Z.; et al. In vitro model on glass surfaces for complex interactions between different types of cells. Langmuir 2010, 26, 17790–17794. [Google Scholar] [CrossRef] [PubMed]
- Abhyankar, V.V.; Lokuta, M.A.; Huttenlocher, A.; Beebe, D.J. Characterization of a membrane-based gradient generator for use in cell-signaling studies. Lab Chip 2006, 6, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.G.; Lin, F.; Jeon, N.L. A microfluidic multi-injector for gradient generation. Lab Chip 2006, 6, 764–768. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, H.J.; Jeon, N.L. Biological applications of microfluidic gradient devices. Integr. Biol. 2010, 2, 584–603. [Google Scholar] [CrossRef] [PubMed]
- Cooper, G.M.; Hausman, R.E. The Cell: A Molecular Approach, 6th ed.; Sinauer Associates: Sunderland, MA, USA, 2013; pp. 58–62, 557–564. [Google Scholar]
- Wright, E.; Neethirajan, S.; Warriner, K.; Retterer, S.; Srijanto, B. Single cell swimming dynamics of Listeria monocytogenes using a nanoporous microfluidic platform. Lab Chip 2014, 14, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Wuichet, K.; Zhulin, I.B. Origins and diversification of a complex signal transduction system in prokaryotes. Sci. Signal. 2010, 3, ra50. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Wake, H.; Ozawa, K.; Kodama, H.; Nakamura, N.; Matsunaga, T. Use of a titanium nitride for electrochemical inactivation of marine bacteria. Environ. Sci. Technol. 1998, 32, 798–801. [Google Scholar] [CrossRef]
- Hong, S.H.; Jeong, J.; Shim, S.; Kang, H.; Kwon, S.; Ahn, K.H.; Yoon, J. Effect of electric currents on bacterial detachment and inactivation. Biotechnol. Bioeng. 2008, 100, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A.; Hassett, D.J. Oxidative and nitrosative stress defence systems in Escherichia coli and Pseudomonas aeruginosa: A model organism of study versus a human opportunistic pathogen. In Stress Response in Pathogenic Bacteria; Kidd, S.P., Henderson, B., Wilson, M., Eds.; Library of Congress Cataloging-in-Publication Data: Oxfordshire, UK, 2011. [Google Scholar]
- Darnton, N.C.; Turner, L.; Rojevsky, S.; Berg, H.C. On torque and tumbling in swimming Escherichia coli. J. Bacteriol. 2007, 189, 1756–1764. [Google Scholar] [CrossRef] [PubMed]
- Niepa, T.H.; Gilbert, J.L.; Ren, D. Controlling Pseudomonas aeruginosa persister cells by weak electrochemical currents and synergistic effects with tobramycin. Biomaterials 2012, 33, 7356–7365. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Baldessari, F.; Gyenge, C.C.; Sato, T.; Chambers, R.D.; Santiago, J.G.; Butcher, E.C. Lymphocyte electrotaxis in vitro and in vivo. J. Immunol. 2008, 181, 2465–2471. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acetic Acid (AA) Concentration | Bacteria Type | 0 mA | 0.07 mA | 0.125 mA | 0.175 mA |
|---|---|---|---|---|---|
| 0% | E. coli | 0.74 ± 0.03 | 0.90 ± 0.02 | 0.82 ± 0.02 | 0.83 ± 0.02 |
| P. aeruginosa | 0.79 ± 0.03 | 0.82 ± 0.02 | 0.87 ± 0.02 | 0.86 ± 0.05 | |
| 0.31% | E. coli | 0.16 ± 0.01 | 0.13 ± 0.01 | 0.10 ± 0.01 | 0.10 ± 0.01 |
| P. aeruginosa | 0.11 ± 0.01 | 0.14 ± 0.01 | 0.12 ± 0.01 | 0.11 ± 0.01 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berthelot, R.; Doxsee, K.; Neethirajan, S. Electroceutical Approach for Impairing the Motility of Pathogenic Bacterium Using a Microfluidic Platform. Micromachines 2017, 8, 207. https://doi.org/10.3390/mi8070207
Berthelot R, Doxsee K, Neethirajan S. Electroceutical Approach for Impairing the Motility of Pathogenic Bacterium Using a Microfluidic Platform. Micromachines. 2017; 8(7):207. https://doi.org/10.3390/mi8070207
Chicago/Turabian StyleBerthelot, Ryan, Kristina Doxsee, and Suresh Neethirajan. 2017. "Electroceutical Approach for Impairing the Motility of Pathogenic Bacterium Using a Microfluidic Platform" Micromachines 8, no. 7: 207. https://doi.org/10.3390/mi8070207
APA StyleBerthelot, R., Doxsee, K., & Neethirajan, S. (2017). Electroceutical Approach for Impairing the Motility of Pathogenic Bacterium Using a Microfluidic Platform. Micromachines, 8(7), 207. https://doi.org/10.3390/mi8070207