DNA Replication Licensing Protein MCM10 Promotes Tumor Progression and Is a Novel Prognostic Biomarker and Potential Therapeutic Target in Breast Cancer

 , , , and
, , , and _Kwok.png)
Abstract
:1. Introduction
2. Results
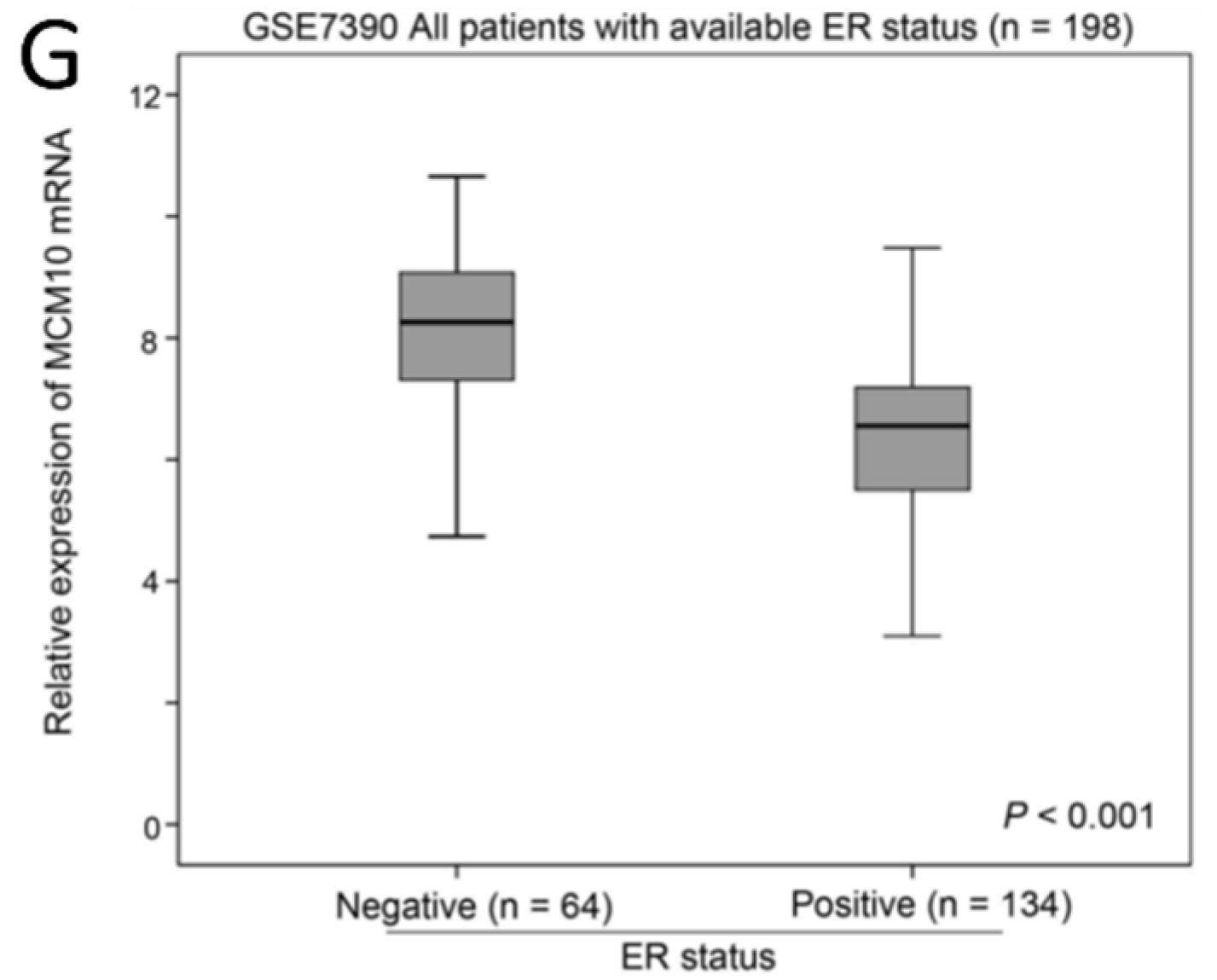
2.1. Association between MCM10 Expression and Tumor Grade
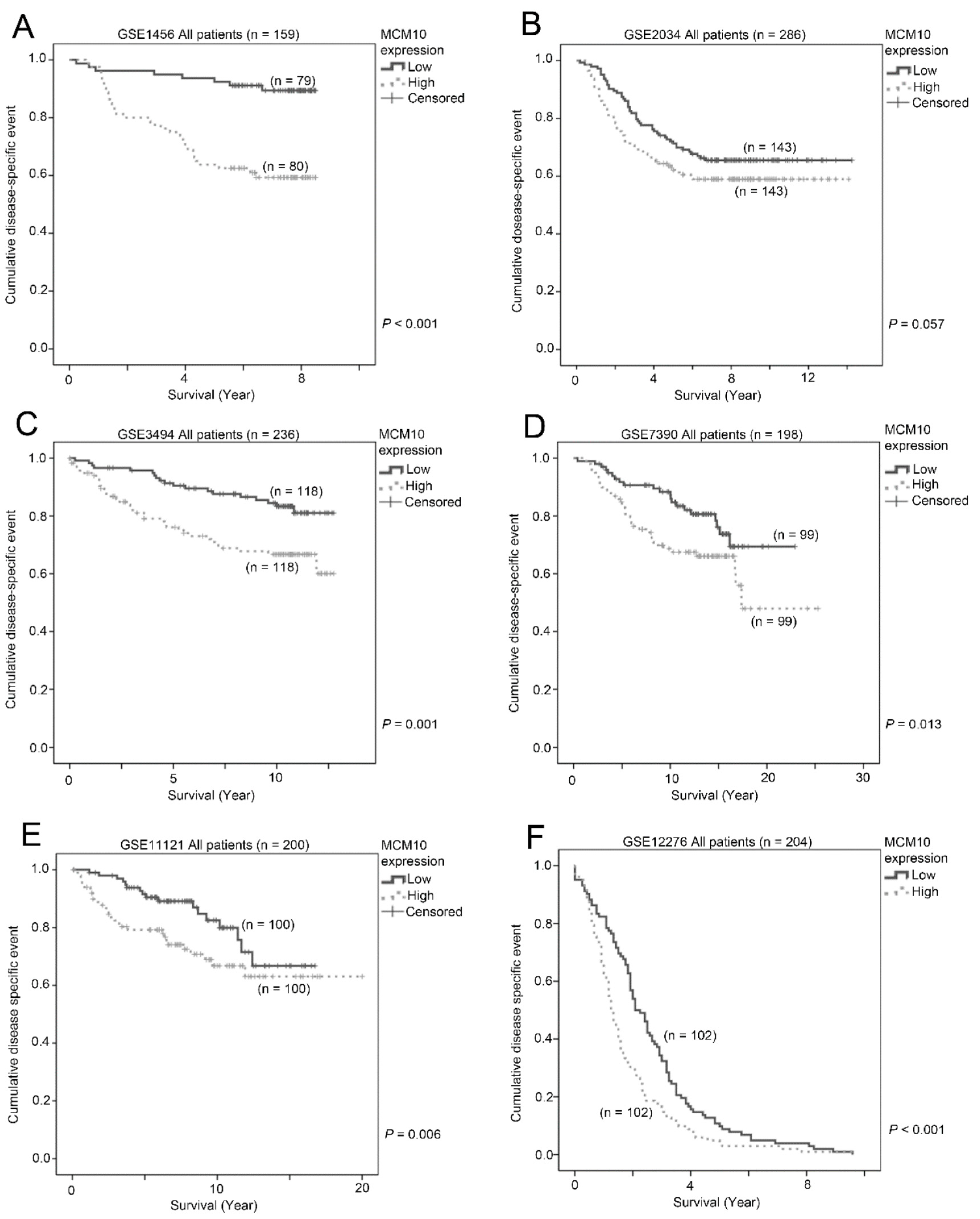
2.2. Association between MCM10 Expression and Patient Survival
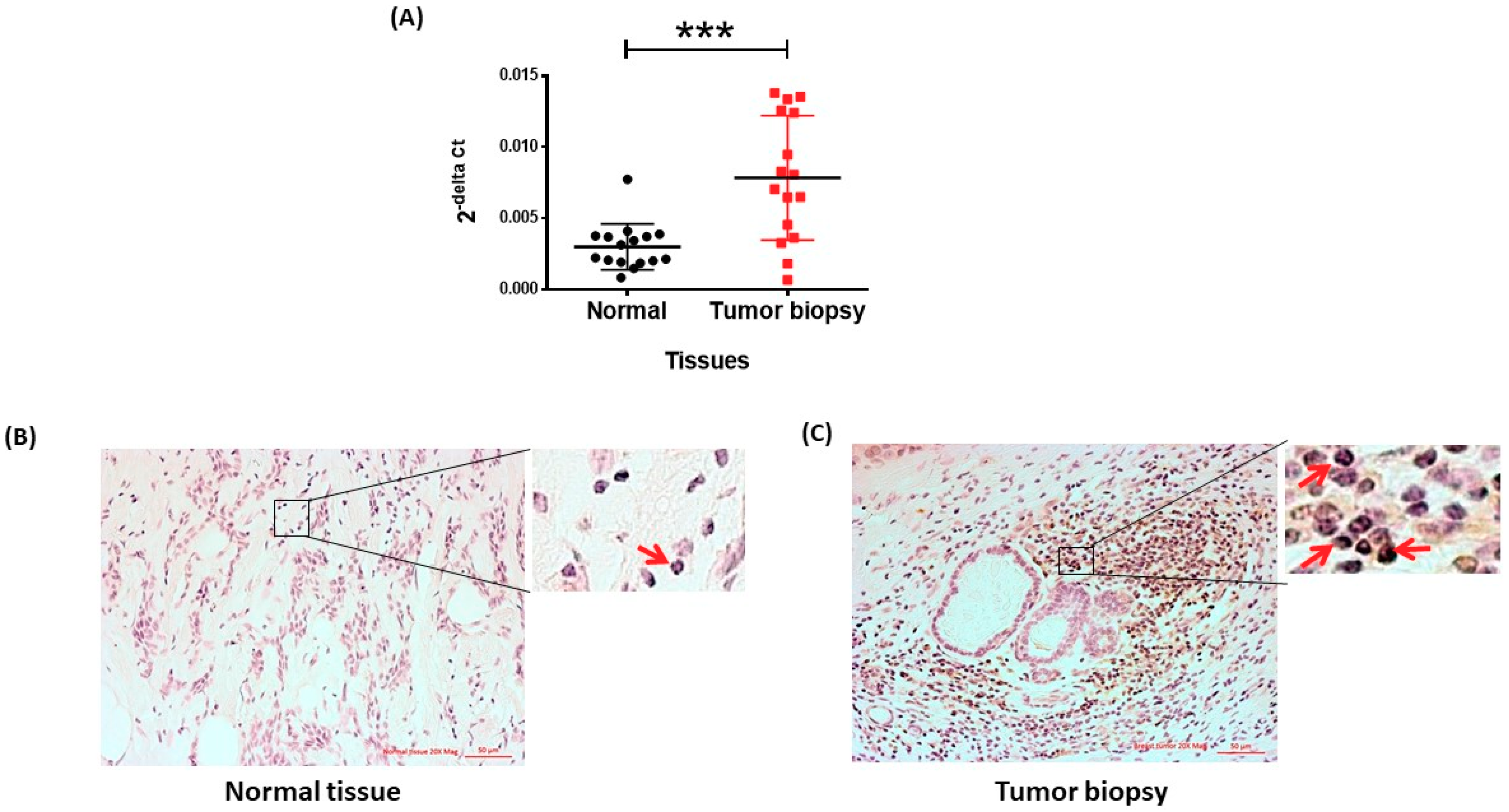
2.3. MCM10 Protein Level Was Increased in Human Breast Cancer Specimens
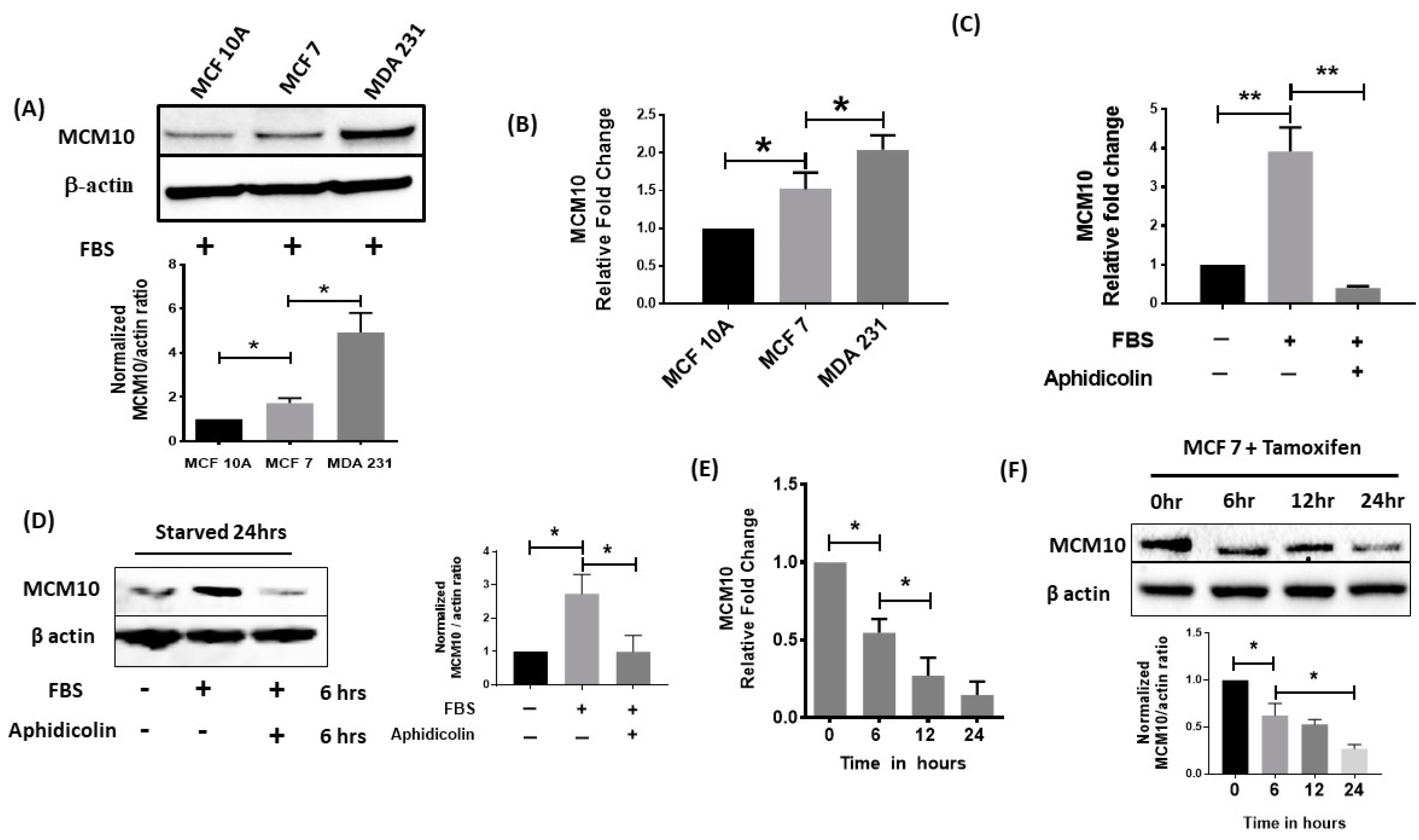
2.4. MCM10 Expression Was Increased in Breast Cancer Cell Lines
2.5. MCM10 Expression Was Regulated by Growth Signalling
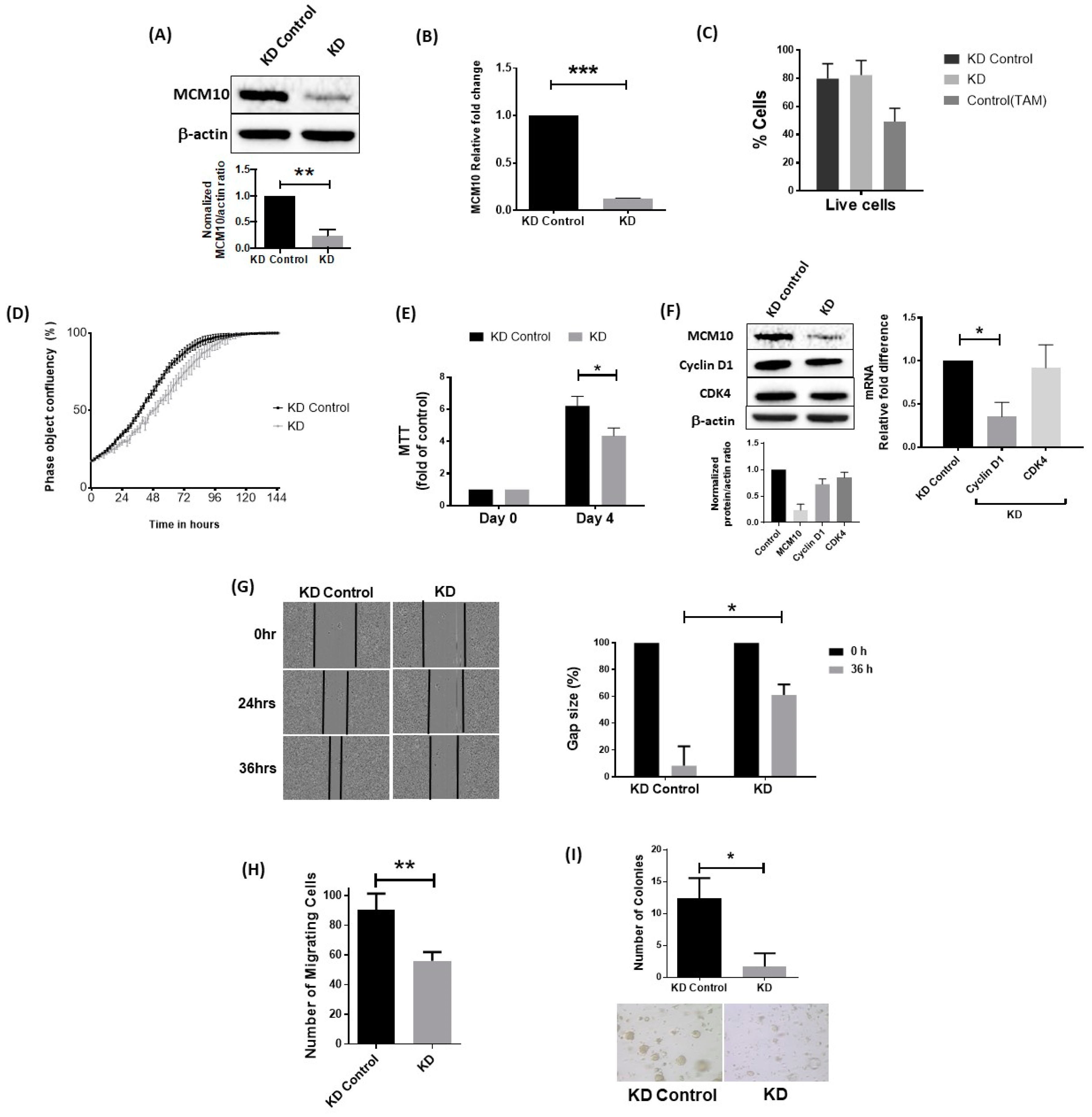
2.6. Knockdown of MCM10 Decreased Cell Proliferation, Cell Migration and Anchorage Independent Growth
2.7. MCM10 Expression Correlated with Tumor Growth In Vivo
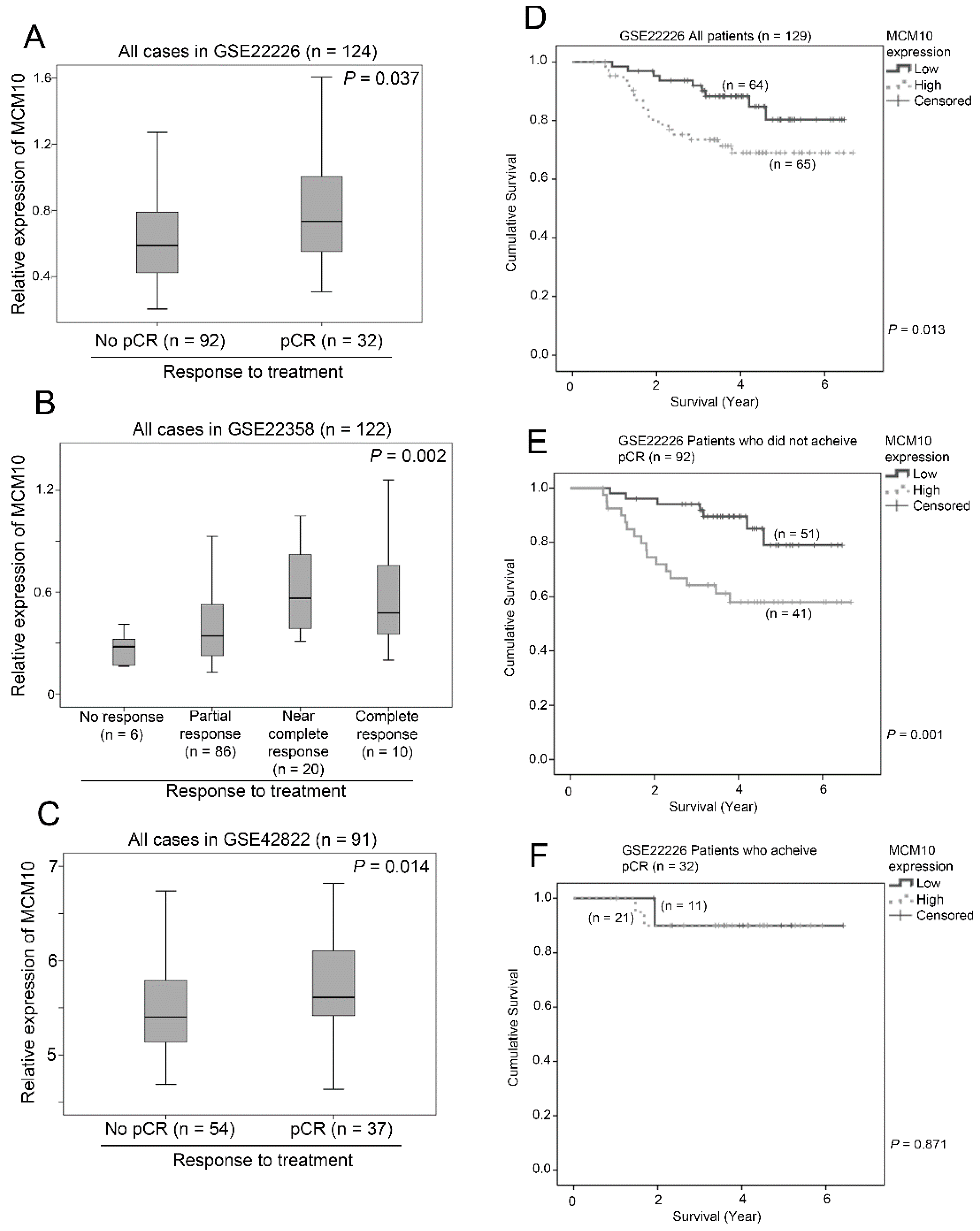
2.8. Association between MCM10 Expression and Treatment Response
3. Discussion
4. Materials and Methods
4.1. Extraction of Clinical and Microarray Gene Expression Data from Breast Cancer Patient Datasets
4.2. Correlations of Gene Expression Levels and Clinical Data
4.3. Cell Culture and Reagents
4.4. Transfection
4.5. Quantitative qPCR
4.6. Western Blotting
4.7. Cell Proliferation by Incucyte Zoom
4.8. MTT Assay
4.9. Wound Healing Assay
4.10. Transwell Migration Assay
4.11. Soft Agar Colony Formation Assay
4.12. Drug Assay
4.13. Assessment of Apoptosis
4.14. Immunohistology
4.15. In Vivo Studies
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DNA | Deoxyribonucleic acid |
| MCM | Minichromosome maintenance proteins |
| GEO | Gene Expression Omnibus |
| CDC6 | Cell division cycle 6 |
| CDT1 | Chromatin licensing and DNA replication factor 1 |
| PCNA | Proliferating cell nuclear antigen |
| HER2 | Human Epidermal Growth Factor Receptor 2 |
| GADPH | Glyceraldehyde 3-phosphate dehydrogenase |
| HRP | Horseradish peroxidase |
| CI | Confidence interval |
| PT | Pre-treatment |
| TGE | Tris-glycine electrophoresis |
| DAB | 3,3′-diaminobenzidine |
| ER | Estrogen receptor |
| qPCR | Quantitative real time polymerase chain reaction |
| pCR | Pathological complete response |
References
- DeSantis, C.; Ma, J.; Bryan, L.; Jemal, A. Breast cancer statistics, 2013. CA Cancer J. Clin. 2014, 64, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Neves, H.; Kwok, H.F. In sickness and in health: The many roles of the minichromosome maintenance proteins. Biochim. Biophys. Acta 2017, 1868, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Mahadevappa, R.; Neves, H.; Yuen, S.M.; Bai, Y.; McCrudden, C.M.; Yuen, H.F.; Wen, Q.; Zhang, S.D.; Kwok, H.F. The prognostic significance of CDC6 and CDT1 in breast cancer. Sci. Rep. 2017, 7, 985. [Google Scholar]
- Kwok, H.F.; Zhang, S.-D.; McCrudden, C.M.; Yuen, H.-F.; Ting, K.-P.; Wen, Q.; Khoo, U.-S.; Chan, K.Y.-K. Prognostic significance of minichromosome maintenance proteins in breast cancer. Am. J. Cancer Res. 2015, 5, 52–71. [Google Scholar] [PubMed]
- Loeb, L.A.; Springgate, C.F.; Battula, N. Errors in DNA replication as a basis of malignant changes. Cancer Res. 1974, 34, 2311–2321. [Google Scholar] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; He, G.; Wang, Y.; Guan, X.; Pang, X.; Zhang, B. MCM-2 is a therapeutic target of trichostatin a in colon cancer cells. Toxicol. Lett. 2013, 221, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Xiong, A.; Zhang, Z.; Li, S.; Huang, H.; Yu, T.T.; Cao, X.; Cheng, S.Y. MicroRNA-31 suppresses medulloblastoma cell growth by inhibiting DNA replication through minichromosome maintenance 2. Oncotarget 2014, 5, 4821–4833. [Google Scholar] [CrossRef] [Green Version]
- Murphy, N.; Ring, M.; Heffron, C.C.; Martin, C.M.; McGuinness, E.; Sheils, O.; O’Leary, J.J. Quantitation of CDC6 and MCM5 mrna in cervical intraepithelial neoplasia and invasive squamous cell carcinoma of the cervix. Mod. Pathol. 2005, 18, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Darnell, J.E., Jr. Transcription factors as targets for cancer therapy. Nat. Rev. Cancer 2002, 2, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, K.M.; Jones, R.M.; Petermann, E.; Jeggo, P.A. Diminished origin-licensing capacity specifically sensitizes tumor cells to replication stress. Mol. Cancer Res. 2013, 11, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Bryant, V.L.; Elias, R.M.; McCarthy, S.M.; Yeatman, T.J.; Alexandrow, M.G. Suppression of reserve MCM complexes chemosensitizes to gemcitabine and 5-fluorouracil. Mol. Cancer Res. 2015, 13, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Yeeles, J.T.; Deegan, T.D.; Janska, A.; Early, A.; Diffley, J.F. Regulated eukaryotic DNA replication origin firing with purified proteins. Nature 2015, 519, 431–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricke, R.M.; Bielinsky, A.K. MCM10 regulates the stability and chromatin association of DNA polymerase-alpha. Mol. Cell 2004, 16, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, S.; Bielinsky, A.K. Human MCM10 regulates the catalytic subunit of DNA polymerase-alpha and prevents DNA damage during replication. Mol. Biol. Cell 2007, 18, 4085–4095. [Google Scholar] [CrossRef] [PubMed]
- Macheret, M.; Halazonetis, T.D. DNA replication stress as a hallmark of cancer. Annu. Rev. Pathol. 2015, 10, 425–448. [Google Scholar] [CrossRef] [PubMed]
- Koppen, A.; Ait-Aissa, R.; Koster, J.; van Sluis, P.G.; Ora, I.; Caron, H.N.; Volckmann, R.; Versteeg, R.; Valentijn, L.J. Direct regulation of the minichromosome maintenance complex by MYCN in neuroblastoma. Eur. J. Cancer 2007, 43, 2413–2422. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Prasad, S.B.; Yadav, S.S.; Govardhan, H.B.; Pandey, L.K.; Singh, S.; Pradhan, S.; Narayan, G. Over expression of minichromosome maintenance genes is clinically correlated to cervical carcinogenesis. PLoS ONE 2013, 8, e69607. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Qiao, J.; He, W.; Wang, J.; Jia, Y.; Sun, Y.; Tang, S.; Fu, L.; Qin, Y. Genome-wide gene expression profile analyses identify CTTN as a potential prognostic marker in esophageal cancer. PLoS ONE 2014, 9, e88918. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.P.; Zhu, Y.; Yin, L.D.; Zhang, J.J.; Guo, S.; Fu, Y.; Miao, Y.; Wei, J.S. The expression and prognostic roles of MCMS in pancreatic cancer. PLoS ONE 2016, 11, e0164150. [Google Scholar] [CrossRef] [PubMed]
- Li, W.M.; Huang, C.N.; Ke, H.L.; Li, C.C.; Wei, Y.C.; Yeh, H.C.; Chang, L.L.; Huang, C.H.; Liang, P.I.; Yeh, B.W.; et al. MCM10 overexpression implicates adverse prognosis in urothelial carcinoma. Oncotarget 2016, 7, 77777–77792. [Google Scholar] [CrossRef] [PubMed]
- Kang, G.; Hwang, W.C.; Do, I.G.; Wang, K.; Kang, S.Y.; Lee, J.; Park, S.H.; Park, J.O.; Kang, W.K.; Jang, J.; et al. Exome sequencing identifies early gastric carcinoma as an early stage of advanced gastric cancer. PLoS ONE 2013, 8, e82770. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhu, J.; Zhang, X. Integrating gene expression and protein-protein interaction network to prioritize cancer-associated genes. BMC Bioinform. 2012, 13, 182. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, A.; Schwob, E.; Mendez, J. Excess mcm proteins protect human cells from replicative stress by licensing backup origins of replication. Proc. Natl. Acad. Sci. USA 2008, 105, 8956–8961. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, H.; Garcia-Muse, T.; Aguilera, A. Replication stress and cancer. Nat. Rev. Cancer 2015, 15, 276–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calaf, G.M.; Roy, D. Gene and protein expressions induced by 17β-estradiol and parathion in cultured breast epithelial cells. Mol. Med. 2007, 13, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ma, N.; Zhou, Z.; Wang, Z.; Hu, Q.; Luo, J.; Mei, X.; Yang, Z.; Zhang, L.; Wang, X.; et al. Estrogen receptor mediates the radiosensitivity of triple-negative breast cancer cells. Med. Sci. Monit. 2017, 23, 2674–2683. [Google Scholar] [CrossRef] [PubMed]
- Pawitan, Y.; Bjohle, J.; Amler, L.; Borg, A.L.; Egyhazi, S.; Hall, P.; Han, X.; Holmberg, L.; Huang, F.; Klaar, S.; et al. Gene expression profiling spares early breast cancer patients from adjuvant therapy: Derived and validated in two population-based cohorts. Breast Cancer Res. 2005, 7, R953–R964. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Klijn, J.G.; Zhang, Y.; Sieuwerts, A.M.; Look, M.P.; Yang, F.; Talantov, D.; Timmermans, M.; Meijer-van Gelder, M.E.; Yu, J.; et al. Gene-expression profiles to predict distant metastasis of lymph-node-negative primary breast cancer. Lancet 2005, 365, 671–679. [Google Scholar] [CrossRef]
- Miller, L.D.; Smeds, J.; George, J.; Vega, V.B.; Vergara, L.; Ploner, A.; Pawitan, Y.; Hall, P.; Klaar, S.; Liu, E.T.; et al. An expression signature for p53 status in human breast cancer predicts mutation status, transcriptional effects, and patient survival. Proc. Natl. Acad. Sci. USA 2005, 102, 13550–13555. [Google Scholar] [CrossRef] [PubMed]
- Desmedt, C.; Piette, F.; Loi, S.; Wang, Y.; Lallemand, F.; Haibe-Kains, B.; Viale, G.; Delorenzi, M.; Zhang, Y.; d’Assignies, M.S.; et al. Strong time dependence of the 76-gene prognostic signature for node-negative breast cancer patients in the transbig multicenter independent validation series. Clin. Cancer Res. 2007, 13, 3207–3214. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Bang, S.W.; Kim, S.H.; Hwang, D.S. Knockdown of human mcm10 activates g2 checkpoint pathway. Biochem. Biophys. Res. Commun. 2008, 365, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Izumi, M.; Yatagai, F.; Hanaoka, F. Localization of human MCM10 is spatially and temporally regulated during the S phase. J. Biol. Chem. 2004, 279, 32569–32577. [Google Scholar] [CrossRef] [PubMed]
- van Deursen, F.; Sengupta, S.; De Piccoli, G.; Sanchez-Diaz, A.; Labib, K. MCM10 associates with the loaded DNA helicase at replication origins and defines a novel step in its activation. EMBO J. 2012, 31, 2195–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, X.Q.; Jackson, D.A.; Blow, J.J. Dormant origins licensed by excess MCM2-7 are required for human cells to survive replicative stress. Genes Dev. 2007, 21, 3331–3341. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Pollard, J.W. KLF15 negatively regulates estrogen-induced epithelial cell proliferation by inhibition of DNA replication licensing. Proc. Natl. Acad. Sci. USA 2012, 109, E1334–E1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, S.; Zhao, J.; Zhou, J.; Zhao, S.; Hu, Y.; Hou, Y. Modulation of 17β-estradiol on the number and cytotoxicity of NK cells in vivo related to mcm and activating receptors. Int. Immunopharmacol. 2007, 7, 1765–1775. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Bohm, D.; von Torne, C.; Steiner, E.; Puhl, A.; Pilch, H.; Lehr, H.A.; Hengstler, J.G.; Kolbl, H.; Gehrmann, M. The humoral immune system has a key prognostic impact in node-negative breast cancer. Cancer Res. 2008, 68, 5405–5413. [Google Scholar] [CrossRef] [PubMed]
- Bos, P.D.; Zhang, X.H.; Nadal, C.; Shu, W.; Gomis, R.R.; Nguyen, D.X.; Minn, A.J.; van de Vijver, M.J.; Gerald, W.L.; Foekens, J.A.; et al. Genes that mediate breast cancer metastasis to the brain. Nature 2009, 459, 1005–1009. [Google Scholar] [CrossRef] [PubMed]
- Esserman, L.J.; Berry, D.A.; Cheang, M.C.; Yau, C.; Perou, C.M.; Carey, L.; DeMichele, A.; Gray, J.W.; Conway-Dorsey, K.; Lenburg, M.E.; et al. Chemotherapy response and recurrence-free survival in neoadjuvant breast cancer depends on biomarker profiles: Results from the I-spy 1 trial (calgb 150007/150012; acrin 6657). Breast Cancer Res. Treat. 2012, 132, 1049–1062. [Google Scholar] [CrossRef] [PubMed]
- Gluck, S.; Ross, J.S.; Royce, M.; McKenna, E.F., Jr.; Perou, C.M.; Avisar, E.; Wu, L. Tp53 genomics predict higher clinical and pathologic tumor response in operable early-stage breast cancer treated with docetaxel-capecitabine +/− trastuzumab. Breast Cancer Res. Treat. 2012, 132, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.; Qi, Y.; Song, N.; Tian, C.; Rice, S.D.; Gabrin, M.J.; Brower, S.L.; Symmans, W.F.; O’Shaughnessy, J.A.; Holmes, F.A.; et al. Cell line derived multi-gene predictor of pathologic response to neoadjuvant chemotherapy in breast cancer: A validation study on us oncology 02-103 clinical trial. BMC Med. Genom. 2012, 5, 51. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Sample Size | Year of the Associated Publication | Available Clinical Parameters | Patient Characteristics (Percent) |
|---|---|---|---|---|
| GSE1456 [26] | 159 | 2005 | Grade | Grade 1 (18) Grade 2 (37) Grade 3 (38) Missing (7) |
| Subtype | Luminal A (25) Luminal B (15) Basal (16) HER2 (9) Normal-like (23) Missing (13) | |||
| GSE2034 [27] | 286 | 2005 | ER status | ER negative (27) ER positive (73) |
| Brain relapse status | With brain relapse (97) Without brain relapse (3) | |||
| GSE3494 [28] | 251 | 2005 | Grade | Grade 1 (27) Grade 2 (51) Grade 3 (22) Missing (1) |
| ER status | ER negative (14) ER positive (85) Missing (2) | |||
| PR status | PR negative (24) PR positive (76) | |||
| Age at diagnosis | Mean = 62 year (SD = 14) | |||
| Tumor size | Mean = 22 mm (SD = 13) | |||
| Lymph node involvement | Negative (63) Positive (34) Missing (4) | |||
| GSE7390 [29] | 198 | 2007 | ER status | ER Negative (32) ER Positive (68) |
| Grade | Grade 1 (15) Grade 2 (42) Grade 3 (42) Missing (1) | |||
| Age | Mean = 46 (SD = 7) | |||
| Tumor size | Mean = 22 mm (SD = 8) | |||
| GSE11121 [30] | 200 | 2008 | Grade | Grade 1 (15) Grade 2 (68) Grade 3 (18) |
| Tumor size | Mean = 21 (SD = 10) | |||
| GSE12276 [31] | 204 | 2009 | Site of relapse | Local (9) Other (83) Brain (4) Brain and other (4) |
| Dataset | No. of Patients | p-Value |
|---|---|---|
| GSE1456 | 159 | <0.001 |
| GSE2034 | 286 | 0.057 |
| GSE3494 | 236 | 0.001 |
| GSE7390 | 198 | 0.013 |
| GSE11121 | 200 | 0.006 |
| GSE12276 | 204 | <0.001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahadevappa, R.; Neves, H.; Yuen, S.M.; Jameel, M.; Bai, Y.; Yuen, H.-F.; Zhang, S.-D.; Zhu, Y.; Lin, Y.; Kwok, H.F. DNA Replication Licensing Protein MCM10 Promotes Tumor Progression and Is a Novel Prognostic Biomarker and Potential Therapeutic Target in Breast Cancer. Cancers 2018, 10, 282. https://doi.org/10.3390/cancers10090282
Mahadevappa R, Neves H, Yuen SM, Jameel M, Bai Y, Yuen H-F, Zhang S-D, Zhu Y, Lin Y, Kwok HF. DNA Replication Licensing Protein MCM10 Promotes Tumor Progression and Is a Novel Prognostic Biomarker and Potential Therapeutic Target in Breast Cancer. Cancers. 2018; 10(9):282. https://doi.org/10.3390/cancers10090282
Chicago/Turabian StyleMahadevappa, Ravikiran, Henrique Neves, Shun Ming Yuen, Muhammad Jameel, Yuchen Bai, Hiu-Fung Yuen, Shu-Dong Zhang, Youzhi Zhu, Yao Lin, and Hang Fai Kwok. 2018. "DNA Replication Licensing Protein MCM10 Promotes Tumor Progression and Is a Novel Prognostic Biomarker and Potential Therapeutic Target in Breast Cancer" Cancers 10, no. 9: 282. https://doi.org/10.3390/cancers10090282
APA StyleMahadevappa, R., Neves, H., Yuen, S. M., Jameel, M., Bai, Y., Yuen, H. -F., Zhang, S. -D., Zhu, Y., Lin, Y., & Kwok, H. F. (2018). DNA Replication Licensing Protein MCM10 Promotes Tumor Progression and Is a Novel Prognostic Biomarker and Potential Therapeutic Target in Breast Cancer. Cancers, 10(9), 282. https://doi.org/10.3390/cancers10090282