Epilepsy Associates with Decreased HIF-1α/STAT5b Signaling in Glioblastoma

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
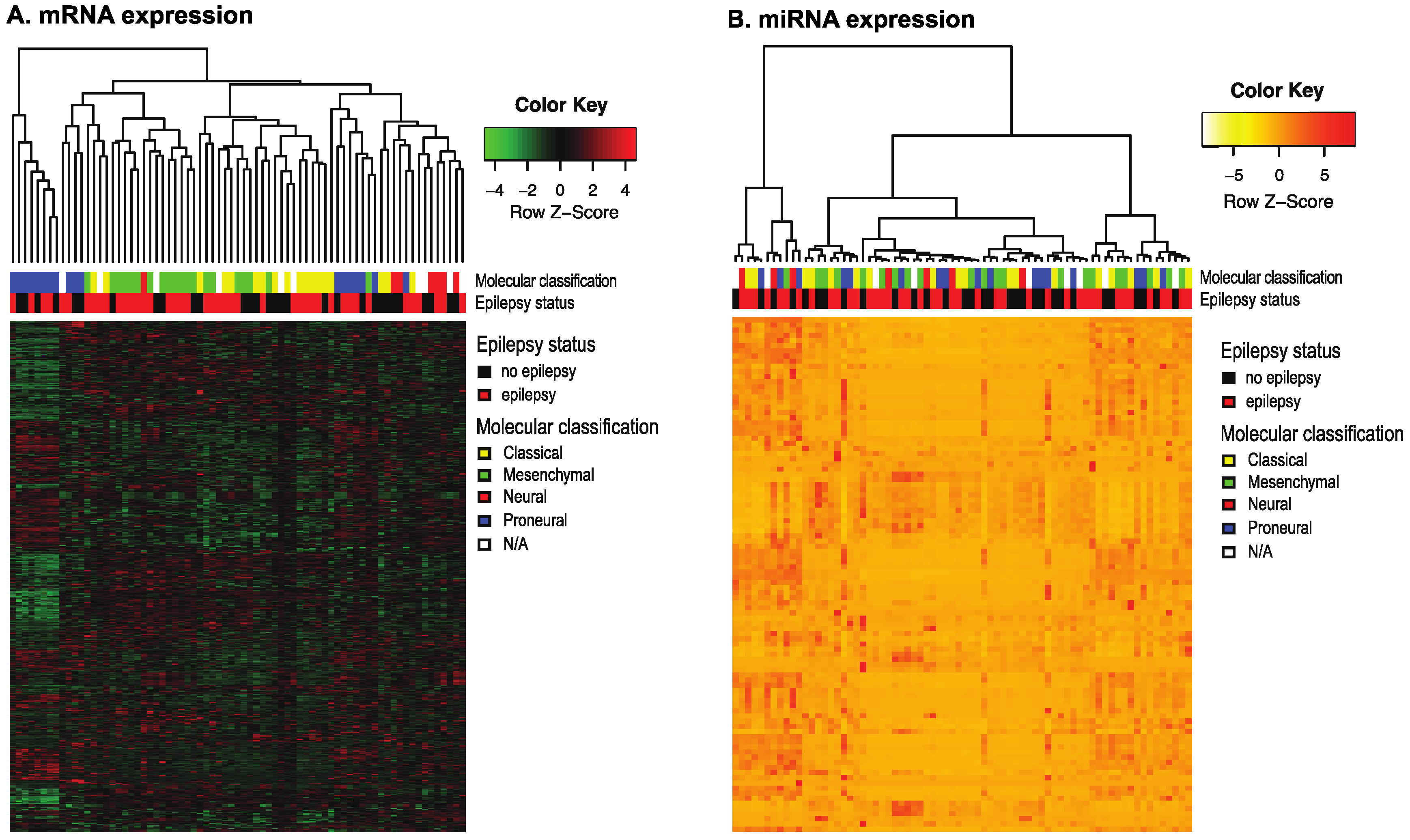
2.1. Epileptogenic GBMs Show Downregulation of HIF1a/STAT5b Signaling
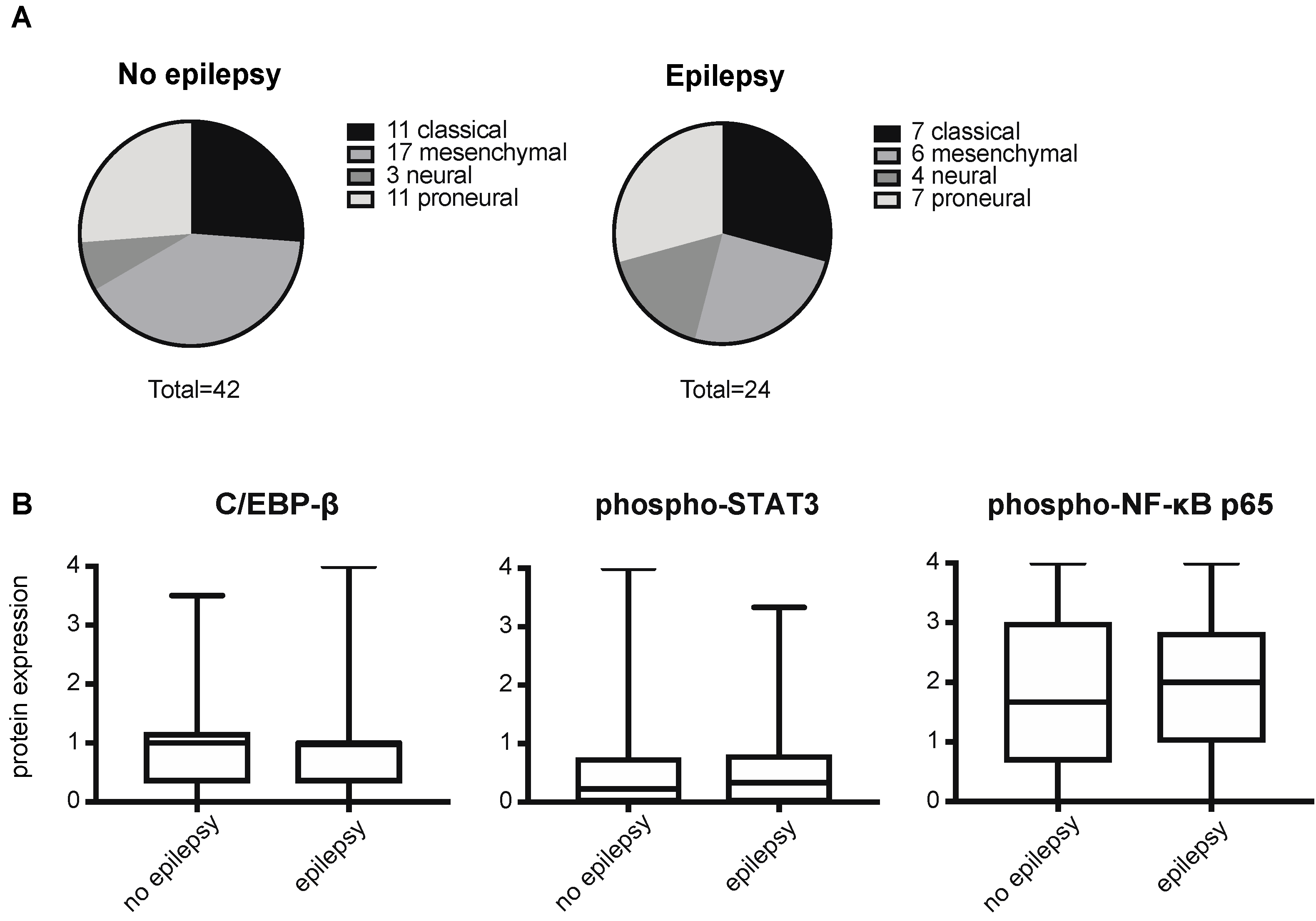
2.2. Epilepsy Does Not Correlate with a Mesenchymal Signature in GBMs
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Clinical data and Tumor Tissues
4.3. mRNA Expression Analysis
4.4. miRNA Expression Analysis
4.5. Class Prediction
4.6. qPCR Analyses
4.7. Tissue Microarrays and Immunohistochemistry
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACTB | Actin β |
| CEBP-β | CCAAT/Enhancer Binding Protein β |
| D2HG | (D)-2-hydroxyglutarate |
| EGFRvIII | Epidermal growth factor receptor variant III |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GUSB | β-glucuronidase |
| HIF-1α | Hypoxia-inducible factor-1α |
| IDH1 | Isocitrate dehydrogenase 1 |
| JAK | Janus kinase |
| LPS | Lipopolysaccharide |
| NF-kB | Nuclear Factor binding near the κ light chain gene in B cells |
| NMDA | N-methyl-D-aspartate |
| NR2B | N-methyl D-aspartate receptor subtype 2B |
| OLIG2 | Oligodendrocyte transcription factor |
| SRF | Serum response factor |
| STAT3 | Signal transducer of activation 3 |
| STAT5 | Signal transducer of activation 5 |
| TAZ | Tafazzin |
| VEGF | Vascular endothelial growth factor |
References
- Stupp, R.; Taillibert, S.; Kanner, A.A.; Kesari, S.; Steinberg, D.M.; Toms, S.A. Maintenance Therapy With Tumor-Treating Fields Plus Temozolomide vs. Temozolomide Alone for Glioblastoma: A Randomized Clinical Trial. JAMA 2015, 314, 2535–2543. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.W.; Verhaak, R.G.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R. The somatic genomic landscape of glioblastoma. Cell 2013, 155, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Carro, M.S.; Lim, W.K.; Alvarez, M.J.; Bollo, R.J.; Zhao, X.; Snyder, E.Y. The transcriptional network for mesenchymal transformation of brain tumours. Nature 2010, 463, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Bhat, K.P.; Balasubramaniyan, V.; Vaillant, B.; Ezhilarasan, R.; Hummelink, K.; Hollingsworth, F. Mesenchymal differentiation mediated by NF-kappaB promotes radiation resistance in glioblastoma. Cancer Cell 2013, 24, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Kerkhof, M.; Dielemans, J.C.; van Breemen, M.S.; Zwinkels, H.; Walchenbach, R.; Taphoorn, M.J.; Vecht, C.J. Effect of valproic acid on seizure control and on survival in patients with glioblastoma multiforme. Neuro-Oncology 2013, 15, 961–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berendsen, S.; Varkila, M.; Kroonen, J.; Seute, T.; Snijders, T.J.; Kauw, F. Prognostic relevance of epilepsy at presentation in glioblastoma patients. Neuro-Oncology 2016, 18, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Happold, C.; Gorlia, T.; Chinot, O.; Gilbert, M.R.; Nabors, L.B.; Wick, W. Does Valproic Acid or Levetiracetam Improve Survival in Glioblastoma? A Pooled Analysis of Prospective Clinical Trials in Newly Diagnosed Glioblastoma. J. Clin. Oncol. 2016, 34, 731–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Breemen, M.S.; Wilms, E.B.; Vecht, C.J. Epilepsy in patients with brain tumours: Epidemiology, mechanisms, and management. Lancet Neurol. 2007, 6, 421–430. [Google Scholar] [CrossRef]
- Dang, L.; White, D.W.; Gross, S.; Bennett, B.D.; Bittinger, M.A.; Driggers, E.M. Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature 2009, 462, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, H.; Cai, S.; Saadatzadeh, M.R.; Hanenberg, H.; Pollok, K.E. Phosphorylation of NMDA 2B at S1303 in human glioma peritumoral tissue: Implications for glioma epileptogenesis. Neurosurg. Focus 2014, 37, E17. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, H.; Pollok, K.E.; Chen, J.; Cohen-Gadol, A.A. Activation of death-associated protein kinase in human peritumoral tissue: A potential therapeutic target. J. Clin. Neurosci. 2015, 22, 1655–1660. [Google Scholar] [CrossRef] [PubMed]
- Yuen, T.I.; Morokoff, A.P.; Bjorksten, A.; D’abaco, G.; Paradiso, L.; Finch, S. Glutamate is associated with a higher risk of seizures in patients with gliomas. Neurology 2012, 79, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Robert, S.M.; Buckingham, S.C.; Campbell, S.L.; Robel, S.; Holt, K.T.; Ogunrinu-Babarinde, T. SLC7A11 expression is associated with seizures and predicts poor survival in patients with malignant glioma. Sci. Transl. Med. 2015, 7, 289ra86. [Google Scholar] [CrossRef] [PubMed]
- Rosati, A. Epilepsy in glioblastoma multiforme: Correlation with glutamine synthetase levels. J. Neurooncol. 2009, 93, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Neal, A. IDH1 and IDH2 mutations in postoperative diffuse glioma-associated epilepsy. Epilepsy Behav. 2018, 78, 30–36. [Google Scholar] [CrossRef]
- Li, Y. IDH1 mutation is associated with a higher preoperative seizure incidence in low-grade glioma: A systematic review and meta-analysis. Seizure 2018, 55, 76–82. [Google Scholar] [CrossRef]
- Chen, H. Mutant IDH1 and seizures in patients with glioma. Neurology 2017, 88, 1805–1813. [Google Scholar] [CrossRef]
- Lee, J.W.; Norden, A.D.; Ligon, K.L.; Golby, A.J.; Beroukhim, R.; Quackenbush, J. Tumor associated seizures in glioblastomas are influenced by survival gene expression in a region-specific manner: A gene expression imaging study. Epilepsy Res. 2014, 108, 843–852. [Google Scholar] [CrossRef] [Green Version]
- Pak, S.H. Hypoxia upregulates Hsp90alpha expression via STAT5b in cancer cells. Int. J. Oncol. 2012, 41, 161–168. [Google Scholar]
- Joung, Y.H. Hypoxia activates the IGF-1 expression through STAT5b in human HepG2 cells. Biochem. Biophys. Res. Commun. 2007, 358, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Joung, Y.H.; Lim, E.J.; Lee, M.Y.; Park, J.H.; Ye, S.K.; Park, E.U. Hypoxia activates the cyclin D1 promoter via the Jak2/STAT5b pathway in breast cancer cells. Exp. Mol. Med. 2005, 37, 353–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.Y. Phosphorylation and activation of STAT proteins by hypoxia in breast cancer cells. Breast 2006, 15, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-kappaB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef]
- van der Krieken, S.E. CCAAT/enhancer binding protein beta in relation to ER stress, inflammation, and metabolic disturbances. Biomed. Res. Int. 2015, 2015, 324815. [Google Scholar] [CrossRef]
- Villarino, A.V. Mechanisms of Jak/STAT signaling in immunity and disease. J. Immunol. 2015, 194, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Cooper, L.A.; Gutman, D.A.; Chisolm, C.; Appin, C.; Kong, J.; Rong, Y. The tumor microenvironment strongly impacts master transcriptional regulators and gene expression class of glioblastoma. Am. J. Pathol. 2012, 180, 2108–2119. [Google Scholar] [CrossRef] [PubMed]
- Bredel, M. Tumor necrosis factor-alpha-induced protein 3 as a putative regulator of nuclear factor-kappaB-mediated resistance to O6-alkylating agents in human glioblastomas. J. Clin. Oncol. 2006, 24, 274–287. [Google Scholar] [CrossRef]
- Carmeliet, P.; Dor, Y.; Herbert, J.M.; Fukumura, D.; Brusselmans, K.; Dewerchin, M. Role of HIF-1alpha in hypoxia-mediated apoptosis, cell proliferation and tumour angiogenesis. Nature 1998, 394, 485–490. [Google Scholar] [CrossRef]
- Joseph, J.V.; Conroy, S.; Pavlov, K.; Sontakke, P.; Tomar, T.; Eggens-Meijer, E. Hypoxia enhances migration and invasion in glioblastoma by promoting a mesenchymal shift mediated by the HIF1alpha-ZEB1 axis. Cancer Lett. 2015, 359, 107–116. [Google Scholar] [CrossRef]
- Talasila, K.M. The angiogenic switch leads to a metabolic shift in human glioblastoma. Neuro-Oncology 2017, 19, 383–393. [Google Scholar] [CrossRef]
- Murat, A. Modulation of angiogenic and inflammatory response in glioblastoma by hypoxia. PLoS ONE 2009, 4, e5947. [Google Scholar] [CrossRef]
- Jung, J.E.; Lee, H.G.; Cho, I.H.; Chung, D.H.; Yoon, S.H.; Yang, Y.M. STAT3 is a potential modulator of HIF-1-mediated VEGF expression in human renal carcinoma cells. FASEB J. 2005, 19, 1296–1298. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, D. The critical role that STAT3 plays in glioma-initiating cells: STAT3 addiction in glioma. Oncotarget 2018, 9, 22095–22112. [Google Scholar] [CrossRef] [Green Version]
- Alkharusi, A. Stimulation of prolactin receptor induces STAT-5 phosphorylation and cellular invasion in glioblastoma multiforme. Oncotarget 2016, 7, 79572–79583. [Google Scholar] [CrossRef] [PubMed]
- Gressot, L.V.; Doucette, T.A.; Yang, Y.; Fuller, G.N.; Heimberger, A.B.; Bögler, O. Signal transducer and activator of transcription 5b drives malignant progression in a PDGFB-dependent proneural glioma model by suppressing apoptosis. Int. J. Cancer 2015, 136, 2047–2054. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.C.; Xiong, H.; Zhao, Z.W.; Jia, D.; Li, W.X.; Qin, H.Z. Inhibition of transcription factor STAT5b suppresses proliferation, induces G1 cell cycle arrest and reduces tumor cell invasion in human glioblastoma multiforme cells. Cancer Lett. 2009, 273, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wang, C.; Zheng, Q.; Qiao, Y.; Xu, K.; Jiang, T.; Wu, A. STAT5 regulates glioma cell invasion by pathways dependent and independent of STAT5 DNA binding. Neurosci. Lett. 2011, 487, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.W. EGFR phosphorylates tumor-derived EGFRvIII driving STAT3/5 and progression in glioblastoma. Cancer Cell 2013, 24, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Roos, A. EGFRvIII-Stat5 Signaling Enhances Glioblastoma Cell Migration and Survival. Mol. Cancer Res. 2018, 16, 1185–1195. [Google Scholar] [CrossRef]
- Latha, K.; Li, M.; Chumbalkar, V.; Gururaj, A.; Hwang, Y.; Dakeng, S. Nuclear EGFRvIII-STAT5b complex contributes to glioblastoma cell survival by direct activation of the Bcl-XL promoter. Int. J. Cancer 2013, 132, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Yamashita, H.; Rui, H.; Waxman, D.J. Serine phosphorylation of GH-activated signal transducer and activator of transcription 5a (STAT5a) and STAT5b: Impact on STAT5 transcriptional activity. Mol. Endocrinol. 2001, 15, 2157–2171. [Google Scholar] [CrossRef] [PubMed]
- Robe, P.A.; University Medical Center Utrecht, Utrecht, The Netherlands. Personal communication, 2018.
- Miyoshi, K.; Shillingford, J.M.; Smith, G.H.; Grimm, S.L.; Wagner, K.U.; Oka, T. Signal transducer and activator of transcription (Stat) 5 controls the proliferation and differentiation of mammary alveolar epithelium. J. Cell Biol. 2001, 155, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Valle-Casuso, J.C. HIF-1 and c-Src mediate increased glucose uptake induced by endothelin-1 and connexin43 in astrocytes. PLoS ONE 2012, 7, e32448. [Google Scholar] [CrossRef]
- Dong, H. Complex role of connexin 43 in astrocytic tumors and possible promotion of gliomaassociated epileptic discharge (Review). Mol. Med. Rep. 2017, 16, 7890–7900. [Google Scholar] [CrossRef]
- Collignon, F. Altered expression of connexin subtypes in mesial temporal lobe epilepsy in humans. J. Neurosurg. 2006, 105, 77–87. [Google Scholar] [CrossRef]
- Linher-Melville, K. Chronic Inhibition of STAT3/STAT5 in Treatment-Resistant Human Breast Cancer Cell Subtypes: Convergence on the ROS/SUMO Pathway and Its Effects on xCT Expression and System xc- Activity. PLoS ONE 2016, 11, e0161202. [Google Scholar] [CrossRef] [PubMed]
- Lösing, P.; Niturad, C.E.; Harrer, M.; zu Reckendorf, C.M.; Schatz, T.; Sinske, D. SRF modulates seizure occurrence, activity induced gene transcription and hippocampal circuit reorganization in the mouse pilocarpine epilepsy model. Mol. Brain 2017, 10, 30. [Google Scholar] [CrossRef]
- Kuzniewska, B. Adult Deletion of SRF Increases Epileptogenesis and Decreases Activity-Induced Gene Expression. Mol. Neurobiol. 2016, 53, 1478–1493. [Google Scholar] [CrossRef]
- Ohrnberger, S. Dysregulated serum response factor triggers formation of hepatocellular carcinoma. Hepatology 2015, 61, 979–989. [Google Scholar] [CrossRef] [Green Version]
- Qiao, J. SRF promotes gastric cancer metastasis through stromal fibroblasts in an SDF1-CXCR4-dependent manner. Oncotarget 2016, 7, 46088–46099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundon, D.J.; Boland, A.; Prencipe, M.; Hurley, G.; O’Neill, A.; Kay, E. The prognostic utility of the transcription factor SRF in docetaxel-resistant prostate cancer: In-vitro discovery and in-vivo validation. BMC Cancer 2017, 17, 163. [Google Scholar] [CrossRef] [PubMed]
- Turkheimer, F.E.; Roncaroli, F.; Hennuy, B.; Herens, C.; Nguyen, M.; Martin, D. Chromosomal patterns of gene expression from microarray data: Methodology, validation and clinical relevance in gliomas. BMC Bioinform. 2006, 7, 526. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, M. GenePattern 2.0. Nat. Genet. 2006, 38, 500–501. [Google Scholar] [CrossRef] [PubMed]
- Wislet-Gendebien, S.; Poulet, C.; Neirinckx, V.; Hennuy, B.; Swingland, J.T.; Laudet, E. In vivo tumorigenesis was observed after injection of in vitro expanded neural crest stem cells isolated from adult bone marrow. PLoS ONE 2012, 7, e46425. [Google Scholar] [CrossRef] [PubMed]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene Sets | NES | p-Value | FDR |
|---|---|---|---|
| C2 collection—curated gene sets | |||
| ELVIDGE_HIF1A_TARGETS_DN | −2.355 | <0.0001 | <0.0001 |
| ELVIDGE_HYPOXIA_BY_DMOG_UP | −2.283 | <0.0001 | 0.0007 |
| ELVIDGE_HIF1A_AND_HIF2A_TARGETS_DN | −2.301 | <0.0001 | 0.0007 |
| ELVIDGE_HYPOXIA_UP | −2.259 | <0.0001 | 0.0009 |
| LEONARD_HYPOXIA | −2.285 | <0.0001 | 0.001 |
| FARDIN_HYPOXIA_11 | −2.198 | <0.0001 | 0.007 |
| PID_HIF1_TFPATHWAY | −2.149 | <0.0001 | 0.02 |
| GROSS_HIF1A_TARGETS_DN | −2.136 | <0.0001 | 0.02 |
| GROSS_HYPOXIA_VIA_ELK3_AND_HIF1A_UP | −2.116 | <0.0001 | 0.03 |
| C3 collection—transcription factor targets | |||
| V$ROAZ_01 | −1.69 | 0.03 | 0.11 |
| V$SRF_01 | −1.71 | 0.02 | 0.13 |
| CCAWWNAAGG_V$SRF_Q4 | −1.71 | 0.01 | 0.16 |
| TTGCWCAAY_V$CEBPB_02 | −1.73 | 0.004 | 0.18 |
| GGGNRMNNYCAT_UNKNOWN | −1.62 | 0.006 | 0.20 |
| KRCTCNNNNMANAGC_UNKNOWN | −1.77 | 0.013 | 0.25 |
| C7 collection—immunologic signatures | |||
| GSE14769_UNSTIM_VS_40MIN_LPS_BMDM_DN | −1.90 | 0.006 | 0.13 |
| GSE37416_CTRL_VS_12H_F_TULARENSIS_LVS_NEUTROPHIL_DN | −1.90 | 0.006 | 0.15 |
| GSE14769_UNSTIM_VS_80MIN_LPS_BMDM_DN | −1.91 | 0.008 | 0.17 |
| GSE14769_UNSTIM_VS_60MIN_LPS_BMDM_DN | −1.91 | 0.006 | 0.21 |
| GSE37416_CTRL_VS_3H_F_TULARENSIS_LVS_NEUTROPHIL_DN | −1.93 | 0.004 | 0.24 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berendsen, S.; Spliet, W.G.M.; Geurts, M.; Van Hecke, W.; Seute, T.; Snijders, T.J.; Bours, V.; Bell, E.H.; Chakravarti, A.; Robe, P.A. Epilepsy Associates with Decreased HIF-1α/STAT5b Signaling in Glioblastoma. Cancers 2019, 11, 41. https://doi.org/10.3390/cancers11010041
Berendsen S, Spliet WGM, Geurts M, Van Hecke W, Seute T, Snijders TJ, Bours V, Bell EH, Chakravarti A, Robe PA. Epilepsy Associates with Decreased HIF-1α/STAT5b Signaling in Glioblastoma. Cancers. 2019; 11(1):41. https://doi.org/10.3390/cancers11010041
Chicago/Turabian StyleBerendsen, Sharon, Wim G. M. Spliet, Marjolein Geurts, Wim Van Hecke, Tatjana Seute, Tom J. Snijders, Vincent Bours, Erica H. Bell, Arnab Chakravarti, and Pierre A. Robe. 2019. "Epilepsy Associates with Decreased HIF-1α/STAT5b Signaling in Glioblastoma" Cancers 11, no. 1: 41. https://doi.org/10.3390/cancers11010041
APA StyleBerendsen, S., Spliet, W. G. M., Geurts, M., Van Hecke, W., Seute, T., Snijders, T. J., Bours, V., Bell, E. H., Chakravarti, A., & Robe, P. A. (2019). Epilepsy Associates with Decreased HIF-1α/STAT5b Signaling in Glioblastoma. Cancers, 11(1), 41. https://doi.org/10.3390/cancers11010041