Impact of Fibroblast-Derived SPARC on Invasiveness of Colorectal Cancer Cells

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Results
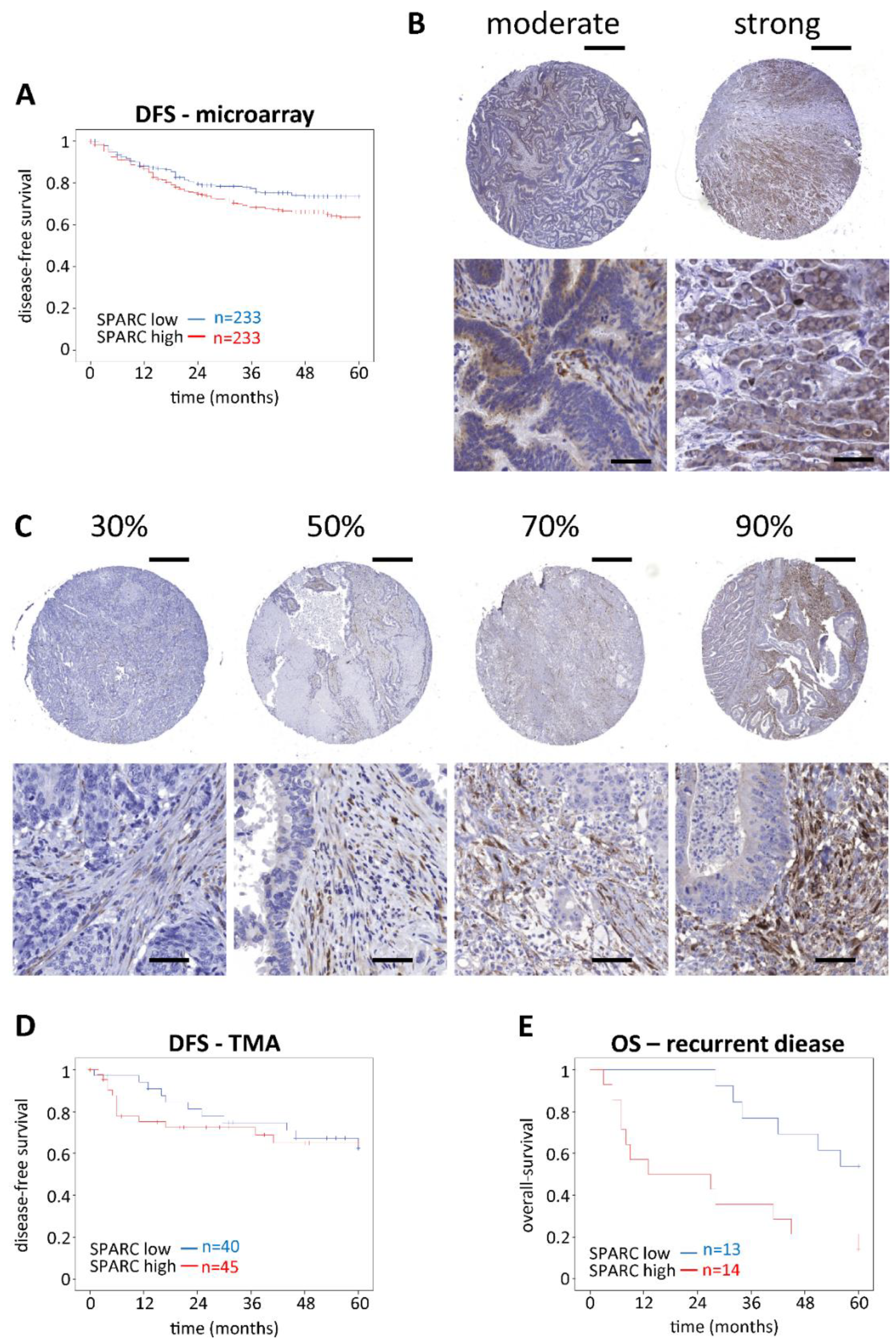
2.1. Impact of SPARC on CRC Outcome
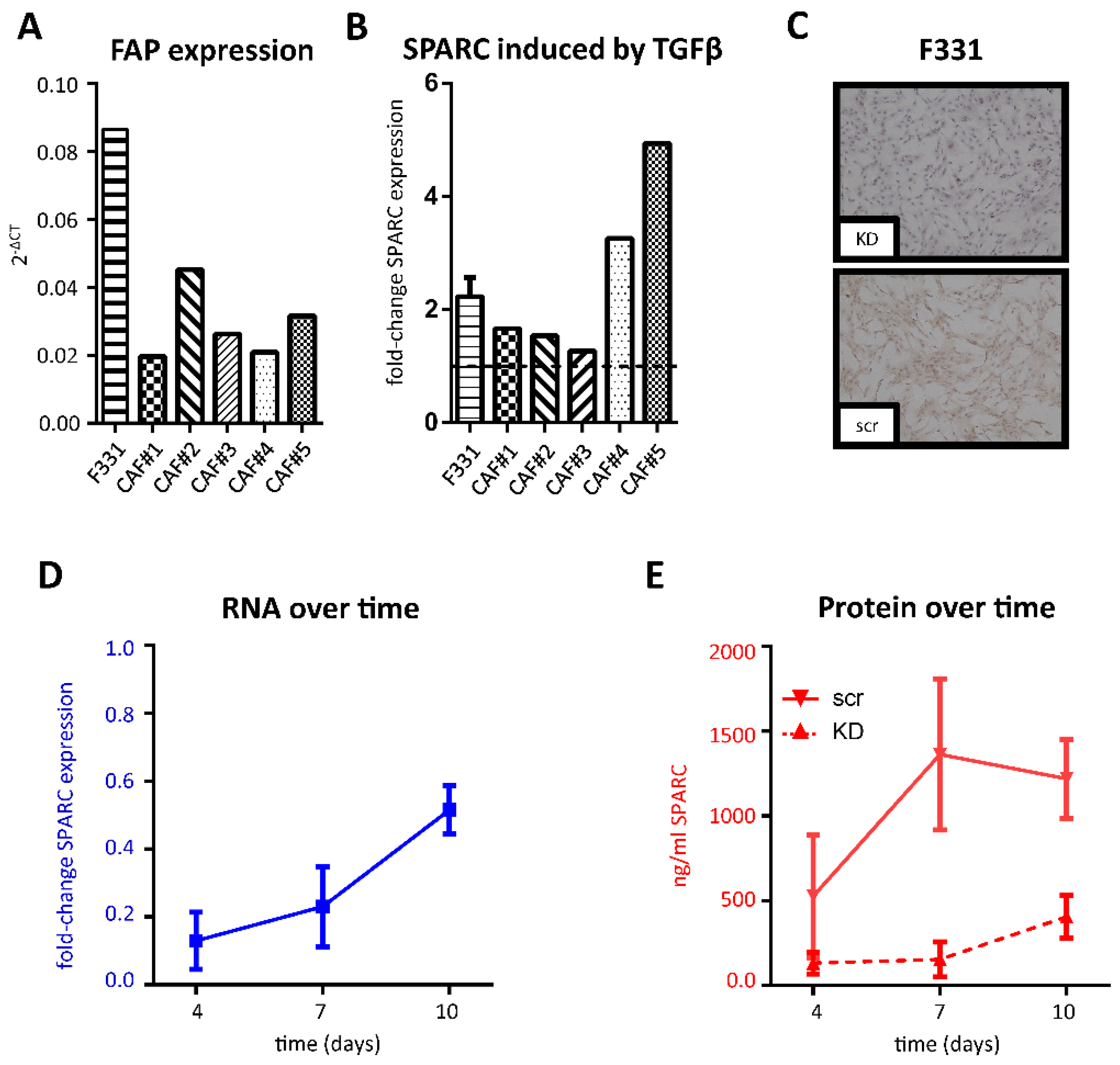
2.2. SPARC Expression in Cancer-Associated Fibroblasts
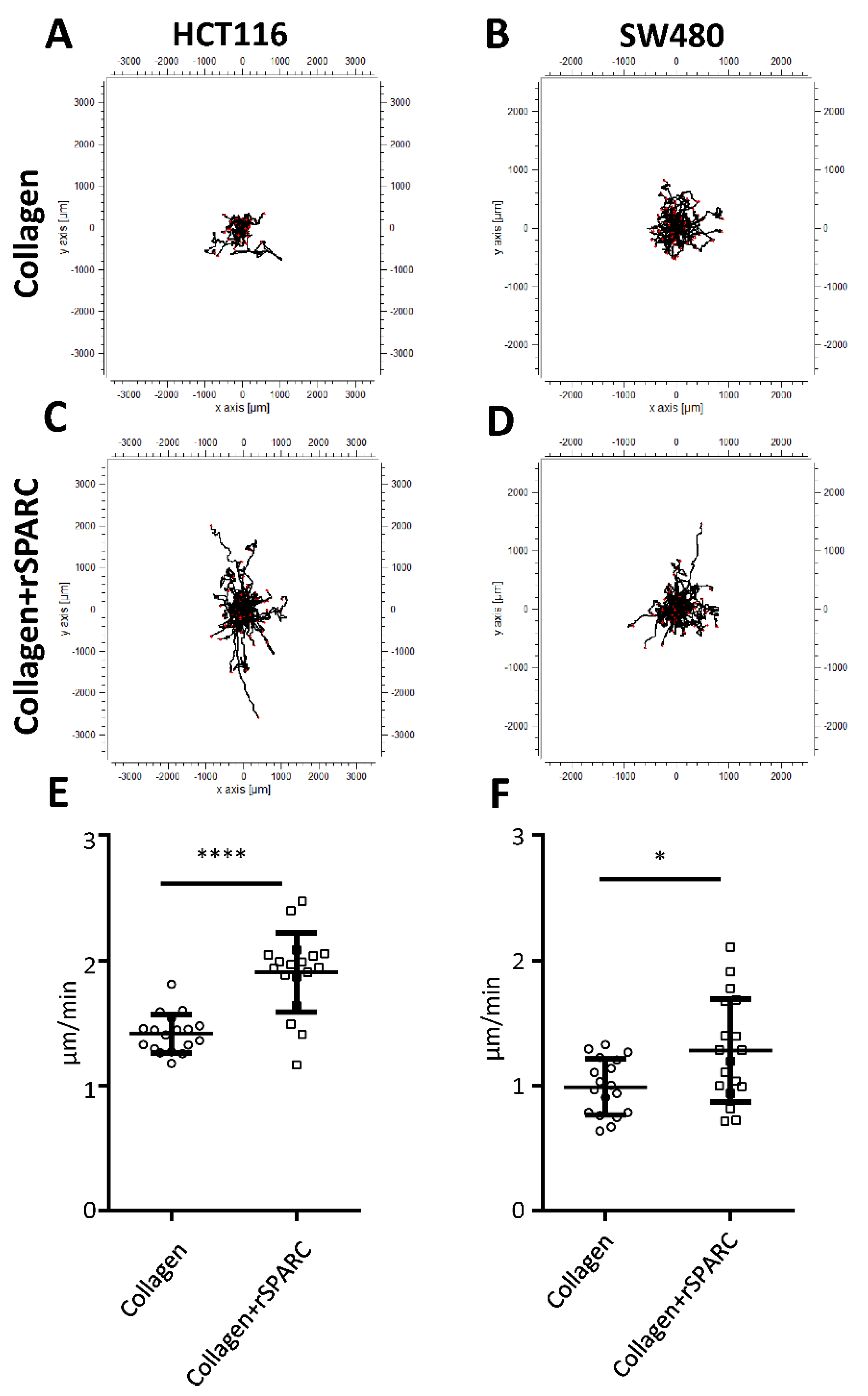
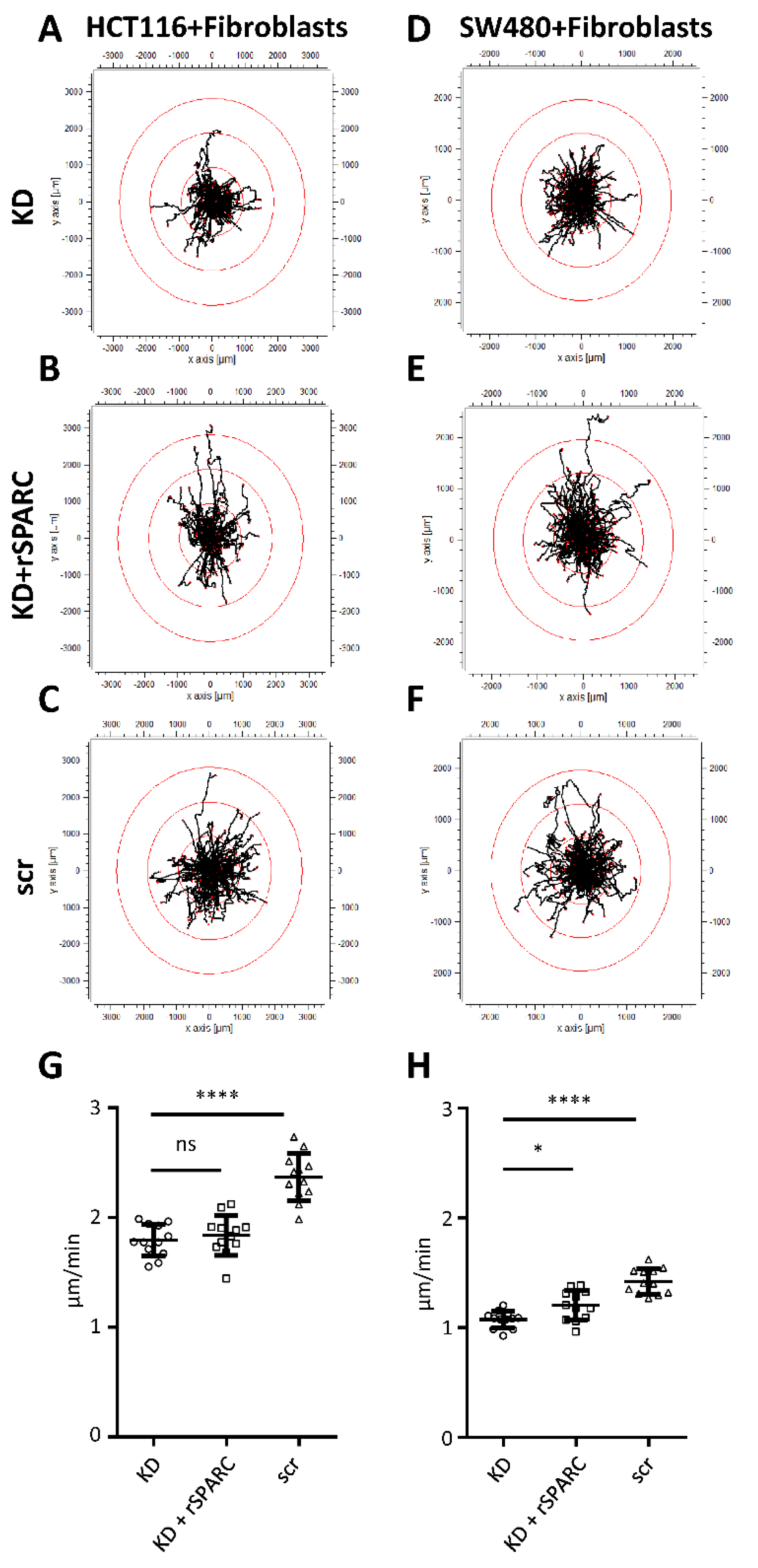
2.3. SPARC and Tumor Cell Migration
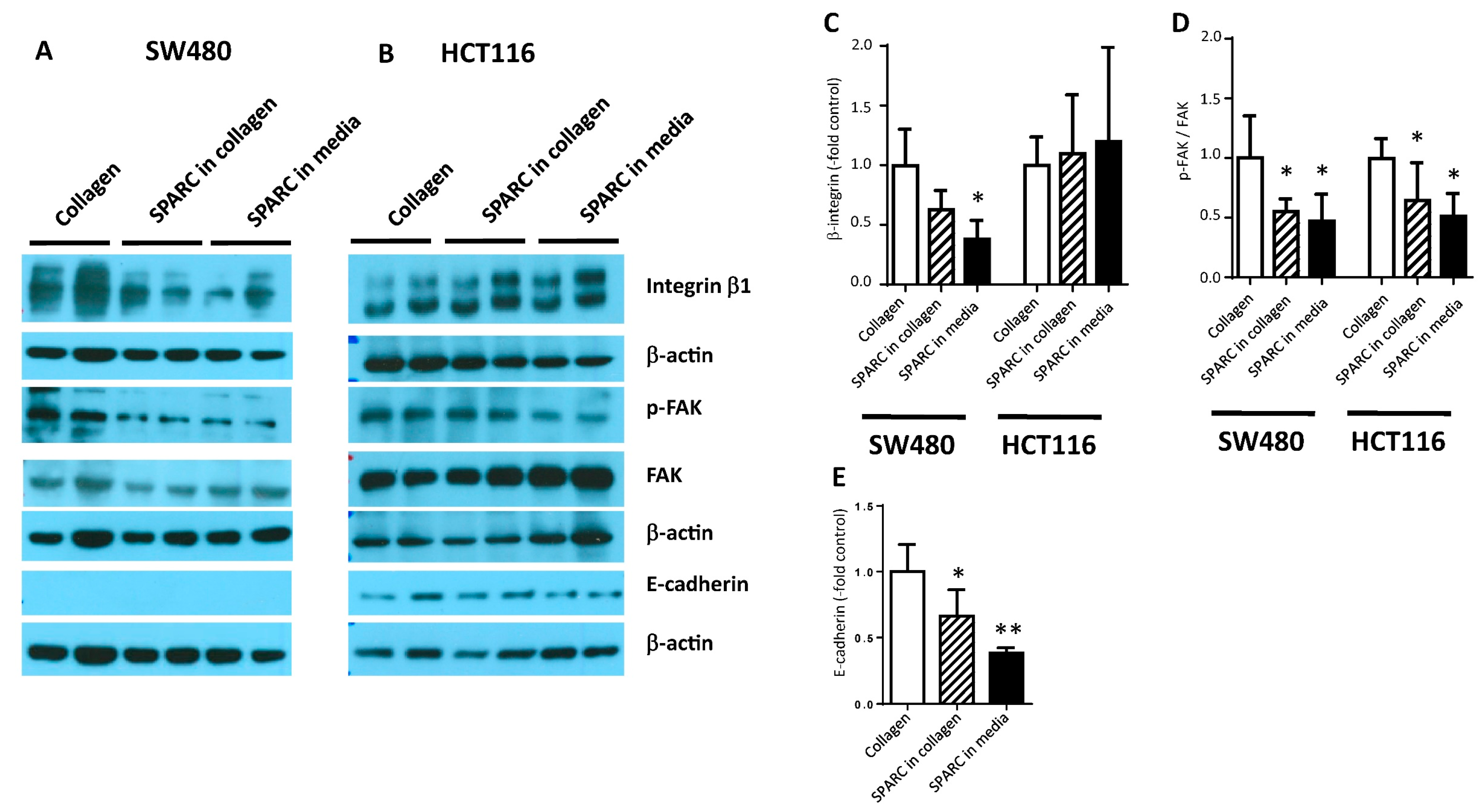
2.4. SPARC Decreased Interaction with the Collagen Matrix
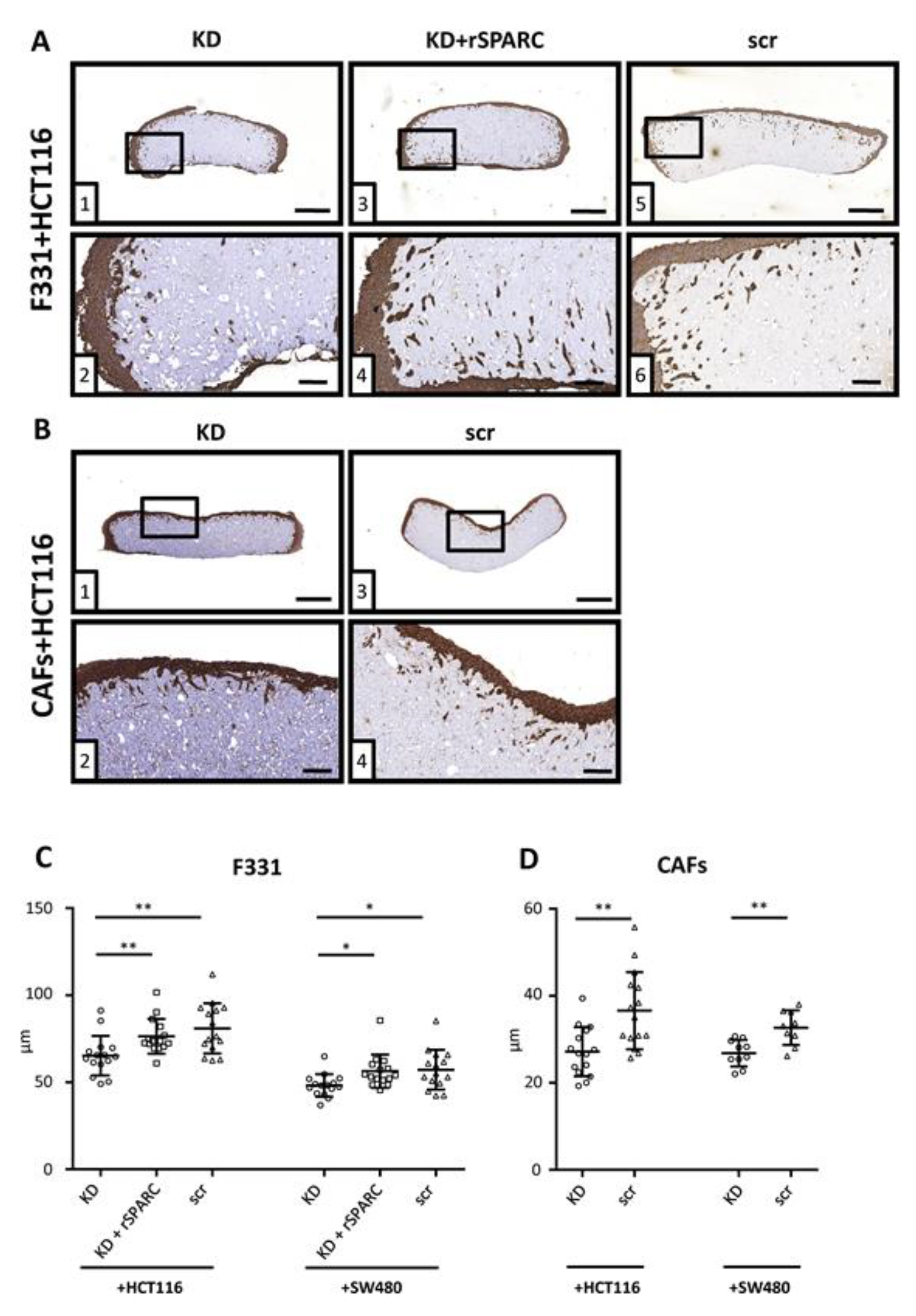
2.5. SPARC and Tumor Cell Invasion
3. Discussion
4. Materials and Methods
4.1. Tissue Acquisition
4.2. Analysis of Gene-Expression Microarray and Tissue Microarray
4.3. Cell Culture
4.4. Isolation of Cancer-Associated Fibroblasts (CAF)
4.5. Transforming Growth Factor Beta (TGFβ) Stimulation Assay
4.6. RNA Isolation and Quantitative Real-Time PCR
4.7. Gene Expression Knock-down
4.8. SPARC Elisa Protein Measurement
4.9. Cancer Cell–Matrix Interactions
4.10. Migration Assay
4.11. Organotypic Cell Culture Model
4.12. Immunohistochemical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lane, T.F.; Sage, E.H. The biology of SPARC, a protein that modulates cell-matrix interactions. FASEB J. 1994, 8, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Kligman, L.H.; Samulewicz, S.J.; Howe, C.C. Impaired wound healing in mice deficient in a matricellular protein SPARC (osteonectin, BM-40). BMC Cell Biol. 2001, 2, 15. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, A.D.; Sage, E.H. SPARC, a matricellular protein that functions in cellular differentiation and tissue response to injury. J. Clin. Investig. 2001, 107, 1049–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaraju, G.P.; Dontula, R.; El-Rayes, B.F.; Lakka, S.S. Molecular mechanisms underlying the divergent roles of SPARC in human carcinogenesis. Carcinogenesis 2014, 35, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Said, N.; Frierson, H.F.; Sanchez-Carbayo, M.; Brekken, R.A.; Theodorescu, D. Loss of SPARC in bladder cancer enhances carcinogenesis and progression. J. Clin. Investig. 2013, 123, 751–766. [Google Scholar] [CrossRef] [PubMed]
- Yiu, G.K.; Chan, W.Y.; Ng, S.W.; Chan, P.S.; Cheung, K.K.; Berkowitz, R.S.; Mok, S.C. SPARC (secreted protein acidic and rich in cysteine) induces apoptosis in ovarian cancer cells. Am. J. Pathol. 2001, 159, 609–622. [Google Scholar] [CrossRef]
- Ledda, M.F.; Adris, S.; Bravo, A.I.; Kairiyama, C.; Bover, L.; Chernajovsky, Y.; Mordoh, J.; Podhajcer, O.L. Suppression of SPARC expression by antisense RNA abrogates the tumorigenicity of human melanoma cells. Nat. Med. 1997, 3, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Lien, H.C.; Hsiao, Y.H.; Lin, Y.S.; Yao, Y.T.; Juan, H.F.; Kuo, W.H.; Hung, M.C.; Chang, K.J.; Hsieh, F.J. Molecular signatures of metaplastic carcinoma of the breast by large-scale transcriptional profiling, Identification of genes potentially related to epithelial-mesenchymal transition. Oncogene 2007, 26, 7859–7871. [Google Scholar] [CrossRef]
- Rempel, S.A.; Golembieski, W.A.; Ge, S.; Lemke, N.; Elisevich, K.; Mikkelsen, T.; Gutierrez, J.A. SPARC: A signal of astrocytic neoplastic transformation and reactive response in human primary and xenograft gliomas. J. Neuropathol. Exp. Neurol. 1998, 57, 1112–1121. [Google Scholar] [CrossRef]
- Tichet, M.; Prod’Homme, V.; Fenouille, N.; Ambrosetti, D.; Mallavialle, A.; Cerezo, M.; Ohanna, M.; Audebert, S.; Rocchi, S.; Giacchero, D.; et al. Tumour-derived SPARC drives vascular permeability and extravasation through endothelial VCAM1 signalling to promote metastasis. Nat. Commun. 2015, 6, 6993. [Google Scholar] [CrossRef] [Green Version]
- Lussier, C.; Sodek, J.; Beaulieu, J.F. Expression of SPARC/osteonectin/BM4O in the human gut, Predominance in the stroma of the remodeling distal intestine. J. Cell. Biochem. 2001, 81, 463–476. [Google Scholar] [CrossRef]
- Chan, S.K.; Griffith, O.L.; Tai, I.T.; Jones, S.J.M. Meta-analysis of Colorectal Cancer Gene Expression Profiling Studies Identifies Consistently Reported Candidate. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 543–552. [Google Scholar] [CrossRef]
- Sansom, O.J.; Mansergh, F.C.; Evans, M.J.; Wilkins, J.A.; Clarke, A.R. Deficiency of SPARC suppresses intestinal tumorigenesis in APCMin/+ mice. Gut 2007, 56, 1410–1414. [Google Scholar] [CrossRef]
- Cheetham, S.; Tang, M.J.; Mesak, F.; Kennecke, H.; Owen, D.; Tai, I.T. SPARC promoter hypermethylation in colorectal cancers can be reversed by 5-Aza-2′deoxycytidine to increase SPARC expression and improve therapy response. Br. J. Cancer 2008, 98, 1810–1819. [Google Scholar] [CrossRef]
- Heitzer, E.; Artl, M.; Filipits, M.; Resel, M.; Graf, R.; Weissenbacher, B.; Lax, S.; Gnant, M.; Wrba, F.; Greil, R.; et al. Differential survival trends of stage II colorectal cancer patients relate to promoter methylation status of PCDH10, SPARC, and UCHL1. Mod. Pathol. 2014, 27, 906–915. [Google Scholar] [CrossRef]
- Yang, E.; Kang, H.J.; Koh, K.H.; Rhee, H.; Kim, N.K.; Kim, H. Frequent inactivation of SPARC by promoter hypermethylation in colon cancers. Int. J. Cancer 2007, 121, 567–575. [Google Scholar] [CrossRef]
- Drev, D.; Bileck, A.; Erdem, Z.N.; Mohr, T.; Timelthaler, G.; Beer, A.; Gerner, C.; Marian, B. Proteomic profiling identifies markers for inflammation-related tumor-fibroblast interaction. Clin. Proteom. 2017, 14, 33. [Google Scholar] [CrossRef]
- Marisa, L.; de Reynies, A.; Duval, A.; Selves, J.; Gaub, M.P.; Vescovo, L.; Etienne-Grimaldi, M.C.; Schiappa, R.; Guenot, D.; Ayadi, M.; et al. Gene expression classification of colon cancer into molecular subtypes, Characterization, validation, and prognostic value. PLoS Med. 2013, 10, e1001453. [Google Scholar] [CrossRef]
- Kalabis, J.; Patterson, M.J.; Enders, G.H.; Marian, B.; Iozzo, R.V.; Rogler, G.; Gimotty, P.A.; Herlyn, M. Stimulation of human colonic epithelial cells by leukemia inhibitory factor is dependent on collagen-embedded fibroblasts in organotypic culture. FASEB J. 2003, 17, 1115–1117. [Google Scholar] [CrossRef]
- Ford, R.; Wang, G.; Jannati, P.; Adler, D.; Racanelli, P.; Higgins, P.J.; Staiano-Coico, L. Modulation of SPARC expression during butyrate-induced terminal differentiation of cultured human keratinocytes, Regulation via a TGF-beta-dependent pathway. Exp. Cell Res. 1993, 206, 261–275. [Google Scholar] [CrossRef]
- Pavasant, P.; Yongchaitrakul, T.; Pattamapun, K.; Arksornnukit, M. The synergistic effect of TGF-beta and 1,25-dihydroxyvitamin D3 on SPARC synthesis and alkaline phosphatase activity in human pulp fibroblasts. Arch. Oral Biol. 2003, 48, 717–722. [Google Scholar] [CrossRef]
- Reed, M.J.; Vernon, R.B.; Abrass, I.B.; Sage, E.H. TGF-beta 1 induces the expression of type I collagen and SPARC, and enhances contraction of collagen gels, by fibroblasts from young and aged donors. J. Cell. Physiol. 1994, 158, 169–179. [Google Scholar] [CrossRef]
- Shibata, S.; Ishiyama, J. Secreted protein acidic and rich in cysteine (SPARC) is upregulated by transforming growth factor (TGF)-beta and is required for TGF-beta-induced hydrogen peroxide production in fibroblasts. Fibrogenesis Tissue Repair 2013, 6, 6. [Google Scholar] [CrossRef]
- Coffey, R.J., Jr.; Goustin, A.s.; Soderquist, A.M.; Shipley, G.D.; Wolfshohl, J.; Carpenter, G.; Moses, H.L. Transforming growth factor alpha and beta expression in human colon cancer lines, Implications for an autocrine model. Cancer Res. 1987, 47, 4590–4594. [Google Scholar]
- Chew, A.; Salama, P.; Robbshaw, A.; Klopcic, B.; Zeps, N.; Platell, C.; Lawrance, I.C. SPARC, FOXP3, CD8 and CD45 correlation with disease recurrence and long-term disease-free survival in colorectal cancer. PLoS ONE 2011, 6, e22047. [Google Scholar] [CrossRef]
- Galamb, O.; Sipos, F.; Spisak, S.; Galamb, B.; Krenacs, T.; Valcz, G.; Tulassay, Z.; Molnar, B. Potential biomarkers of colorectal adenoma-dysplasia-carcinoma progression, mRNA expression profiling and in situ protein detection on TMAs reveal 15 sequentially upregulated and 2 downregulated genes. Cell. Oncol. 2009, 31, 19–29. [Google Scholar]
- Liu, Q.Z.; Gao, X.H.; Chang, W.J.; Wang, H.T.; Wang, H.; Cao, G.W.; Fu, C.G. Secreted protein acidic and rich in cysteine expression in human colorectal cancer predicts postoperative prognosis. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1803–1811. [Google Scholar]
- Tumbarello, D.A.; Andrews, M.R.; Brenton, J.D. SPARC Regulates Transforming Growth Factor Beta Induced (TGFBI) Extracellular Matrix Deposition and Paclitaxel Response in Ovarian Cancer Cells. PLoS ONE 2016, 11, e0162698. [Google Scholar] [CrossRef]
- Tai, I.T.; Dai, M.; Owen, D.A.; Chen, L.B. Genome-wide expression analysis of therapy-resistant tumors reveals SPARC as a novel target for cancer therapy. J. Clin. Investig. 2005, 115, 1492–1502. [Google Scholar] [CrossRef] [Green Version]
- Hynes, R.O. Integrins, Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef]
- Casal, J.I.; Bartolome, R.A. RGD cadherins and alpha2beta1 integrin in cancer metastasis, A dangerous liaison. Biochim. Biophys. Acta Rev. Cancer 2018, 1869, 321–332. [Google Scholar] [CrossRef]
- Klampfl, T.; Bogner, E.; Bednar, W.; Mager, L.; Massudom, D.; Kalny, I.; Heinzle, C.; Berger, W.; Stattner, S.; Karner, J.; et al. Up-regulation of 12(S)-lipoxygenase induces a migratory phenotype in colorectal cancer cells. Exp. Cell Res. 2012, 318, 768–778. [Google Scholar] [CrossRef] [Green Version]
- Sulzmaier, F.J.; Jean, C.; Schlaepfer, D.D. FAK in cancer, Mechanistic findings and clinical applications. Nat. Rev. Cancer 2014, 14, 598–610. [Google Scholar] [CrossRef]
- Shi, Q.; Bao, S.; Song, L.; Wu, Q.; Bigner, D.D.; Hjelmeland, A.B.; Rich, J.N. Targeting SPARC expression decreases glioma cellular survival and invasion associated with reduced activities of FAK and ILK kinases. Oncogene 2007, 26, 4084–4094. [Google Scholar] [CrossRef] [Green Version]
- Smit, D.J.; Gardiner, B.B.; Sturm, R.A. Osteonectin downregulates E-cadherin, induces osteopontin and focal adhesion kinase activity stimulating an invasive melanoma phenotype. Int. J. Cancer 2007, 121, 2653–2660. [Google Scholar] [CrossRef]
- Said, N.; Najwer, I.; Motamed, K. Secreted protein acidic and rich in cysteine (SPARC) inhibits integrin-mediated adhesion and growth factor-dependent survival signaling in ovarian cancer. Am. J. Pathol. 2007, 170, 1054–1063. [Google Scholar] [CrossRef]
- Goyette, M.C.; Cho, K.; Fasching, C.L.; Levy, D.B.; Kinzler, K.W.; Paraskeva, C.; Vogelstein, B.; Stanbridge, E.J. Progression of colorectal cancer is associated with multiple tumor suppressor gene defects but inhibition of tumorigenicity is accomplished by correction of any single defect via chromosome transfer. Mol. Cell. Biol. 1992, 12, 1387–1395. [Google Scholar] [CrossRef]
- Mouradov, D.; Sloggett, C.; Jorissen, R.N.; Love, C.G.; Li, S.; Burgess, A.W.; Arango, D.; Strausberg, R.L.; Buchanan, D.; Wormald, S.; et al. Colorectal cancer cell lines are representative models of the main molecular subtypes of primary cancer. Cancer Res. 2014, 74, 3238–3247. [Google Scholar] [CrossRef]
- Yin, J.; Chen, G.; Liu, Y.; Liu, S.; Wang, P.; Wan, Y.; Wang, X.; Zhu, J.; Gao, H. Downregulation of SPARC expression decreases gastric cancer cellular invasion and survival. J. Exp. Clin. Cancer Res. 2010, 29, 59. [Google Scholar] [CrossRef]
- Barbagallo, C.; Brex, D.; Caponnetto, A.; Cirnigliaro, M.; Scalia, M.; Magnano, A.; Caltabiano, R.; Barbagallo, D.; Biondi, A.; Cappellani, A.; et al. LncRNA UCA1, Upregulated in CRC Biopsies and Downregulated in Serum Exosomes, Controls mRNA Expression by RNA-RNA Interactions. Mol. Ther. Nucleic Acids 2018, 12, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Falzone, L.; Scola, L.; Zanghi, A.; Biondi, A.; Di Cataldo, A.; Libra, M.; Candido, S. Integrated analysis of colorectal cancer microRNA datasets, Identification of microRNAs associated with tumor development. Aging 2018, 10, 1000–1014. [Google Scholar] [CrossRef]
- Deng, B.; Qu, L.; Li, J.; Fang, J.; Yang, S.; Cao, Z.; Mei, Z.; Sun, X. MiRNA-211 suppresses cell proliferation, migration and invasion by targeting SPARC in human hepatocellular carcinoma. Sci. Rep 2016, 6, 26679. [Google Scholar] [CrossRef] [Green Version]
- Sonvilla, G.; Allerstorfer, S.; Heinzle, C.; Stattner, S.; Karner, J.; Klimpfinger, M.; Wrba, F.; Fischer, H.; Gauglhofer, C.; Spiegl-Kreinecker, S.; et al. Fibroblast growth factor receptor 3-IIIc mediates colorectal cancer growth and migration. Br. J. Cancer 2010, 102, 1145–1156. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji, An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Gao, Y.; Yin, S.P.; Xie, X.S.; Xu, D.D.; Du, W.D. The relationship between stromal cell derived SPARC in human gastric cancer tissue and its clinicopathologic significance. Oncotarget 2017, 8, 86240–86252. [Google Scholar] [CrossRef] [Green Version]
- Shintakuya, R.; Kondo, N.; Murakami, Y.; Uemura, K.; Nakagawa, N.; Okano, K.; Takahashi, S.; Sueda, T. The high stromal SPARC expression is independently associated with poor survival of patients with resected pancreatic ductal adenocarcinoma treated with adjuvant gemcitabine in combination with S-1 or adjuvant gemcitabine alone. Pancreatology 2018, 18, 191–197. [Google Scholar] [CrossRef]
- Toyota, K.; Murakami, Y.; Kondo, N.; Uemura, K.; Nakagawa, N.; Takahashi, S.; Sueda, T. Impact of Secreted Protein Acidic and Rich in Cysteine (SPARC) Expression on Prognosis After Surgical Resection for Biliary Carcinoma. J. Gastrointest. Surg. 2017, 21, 990–999. [Google Scholar] [CrossRef]
- Lindner, J.L.; Loibl, S.; Denkert, C.; Ataseven, B.; Fasching, P.A.; Pfitzner, B.M.; Gerber, B.; Gade, S.; Darb-Esfahani, S.; Sinn, B.V.; et al. Expression of secreted protein acidic and rich in cysteine (SPARC) in breast cancer and response to neoadjuvant chemotherapy. Ann. Oncol. 2014, 26, 95–100. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Univariate Cox Analysis | Multivariate Cox Analysis | ||||
|---|---|---|---|---|---|---|
| HR | 95% CI | p-Value | HR | 95% CI | p-Value | |
| Gender (male vs. female) | 1.254 | 0.885–1.777 | 0.202 | |||
| Age | 1.009 | 0.995–1.023 | 0.199 | |||
| Staging (III vs. II) | 1.958 | 1.388–2.764 | <0.0001 | 1.836 | 1.221–3.761 | 0.003 |
| Chemotherapy (yes vs. no) | 1.526 | 1.085–2.146 | 0.015 | 1.096 | 0.732–1.641 | 0.658 |
| Location (proximal vs. distal) | 0.877 | 0.618–1.246 | 0.464 | |||
| SPARC (high vs. low) | 1.410 | 1.001–1.989 | 0.050 | 1.408 | 0.998–1.985 | 0.051 |
| Variables | Univariate Cox Analysis | Multivariate Cox Analysis | ||||
|---|---|---|---|---|---|---|
| HR | 95% CI | p-Value | HR | 95% CI | p-Value | |
| Gender (male vs. female) | 2.256 | 0.800–6.366 | 0.124 | |||
| Age | 1.052 | 1.001–1.105 | 0.045 | 1.053 | 0.999–1.109 | 0.055 |
| Staging (III vs. II) | 1.907 | 0.550–6.608 | 0.309 | |||
| Chemotherapy (yes vs. no) | 0.744 | 0.263–2.107 | 0.578 | |||
| Location (proximal vs. distal) | 2.362 | 0.932–5.992 | 0.070 | 0.338 | 0.123–0.931 | 0.036 |
| SPARC (high vs. low) | 3.607 | 1.337–9.732 | 0.011 | 4.574 | 1.609–13.004 | 0.004 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drev, D.; Harpain, F.; Beer, A.; Stift, A.; Gruber, E.S.; Klimpfinger, M.; Thalhammer, S.; Reti, A.; Kenner, L.; Bergmann, M.; et al. Impact of Fibroblast-Derived SPARC on Invasiveness of Colorectal Cancer Cells. Cancers 2019, 11, 1421. https://doi.org/10.3390/cancers11101421
Drev D, Harpain F, Beer A, Stift A, Gruber ES, Klimpfinger M, Thalhammer S, Reti A, Kenner L, Bergmann M, et al. Impact of Fibroblast-Derived SPARC on Invasiveness of Colorectal Cancer Cells. Cancers. 2019; 11(10):1421. https://doi.org/10.3390/cancers11101421
Chicago/Turabian StyleDrev, Daniel, Felix Harpain, Andrea Beer, Anton Stift, Elisabeth S. Gruber, Martin Klimpfinger, Sabine Thalhammer, Andrea Reti, Lukas Kenner, Michael Bergmann, and et al. 2019. "Impact of Fibroblast-Derived SPARC on Invasiveness of Colorectal Cancer Cells" Cancers 11, no. 10: 1421. https://doi.org/10.3390/cancers11101421
APA StyleDrev, D., Harpain, F., Beer, A., Stift, A., Gruber, E. S., Klimpfinger, M., Thalhammer, S., Reti, A., Kenner, L., Bergmann, M., & Marian, B. (2019). Impact of Fibroblast-Derived SPARC on Invasiveness of Colorectal Cancer Cells. Cancers, 11(10), 1421. https://doi.org/10.3390/cancers11101421