ANLN and TLE2 in Muscle Invasive Bladder Cancer: A Functional and Clinical Evaluation Based on In Silico and In Vitro Data

 ,
,  and
and
Abstract
:1. Introduction
2. Results
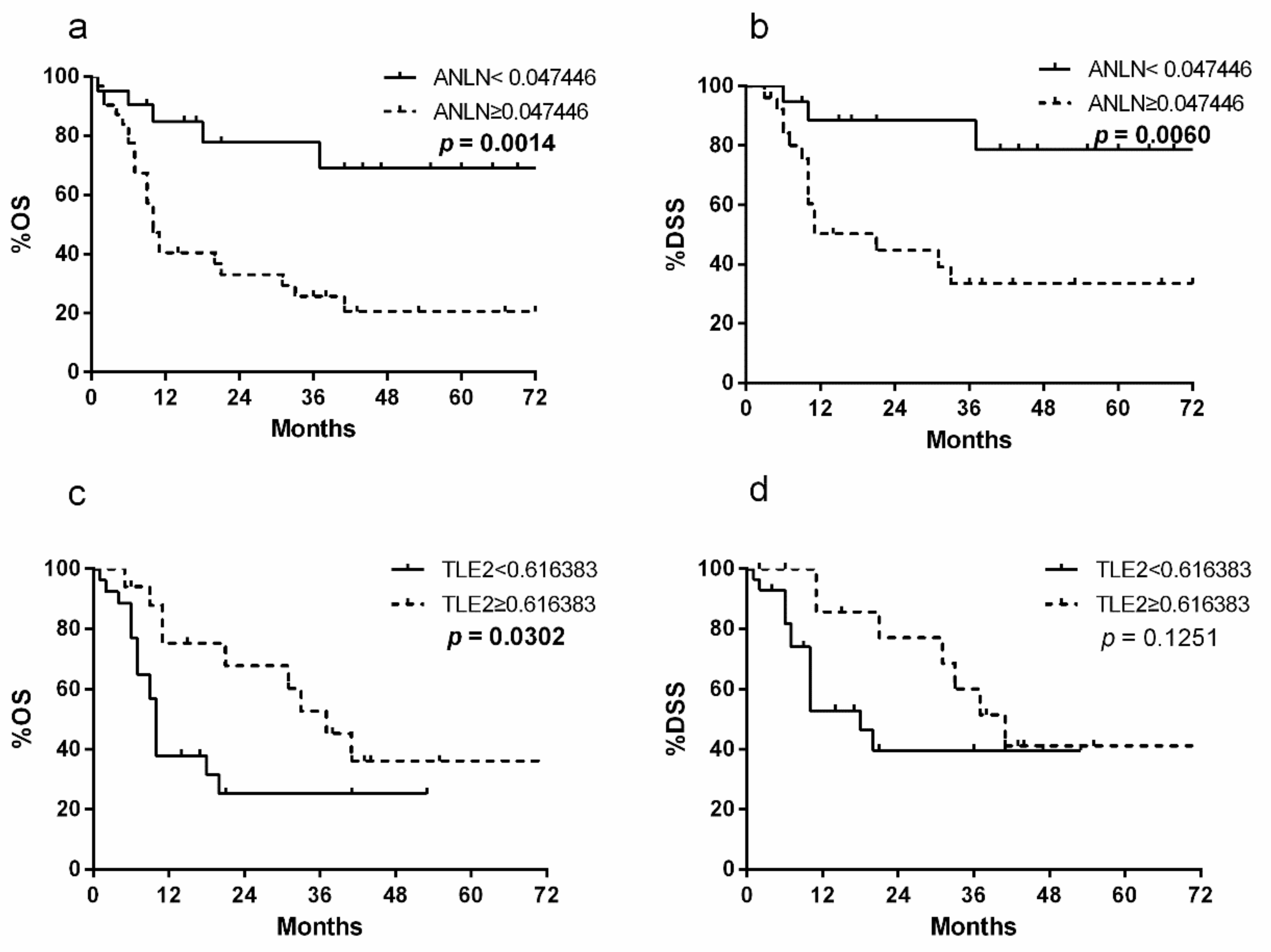
2.1. Patient Population and Survival Analysis
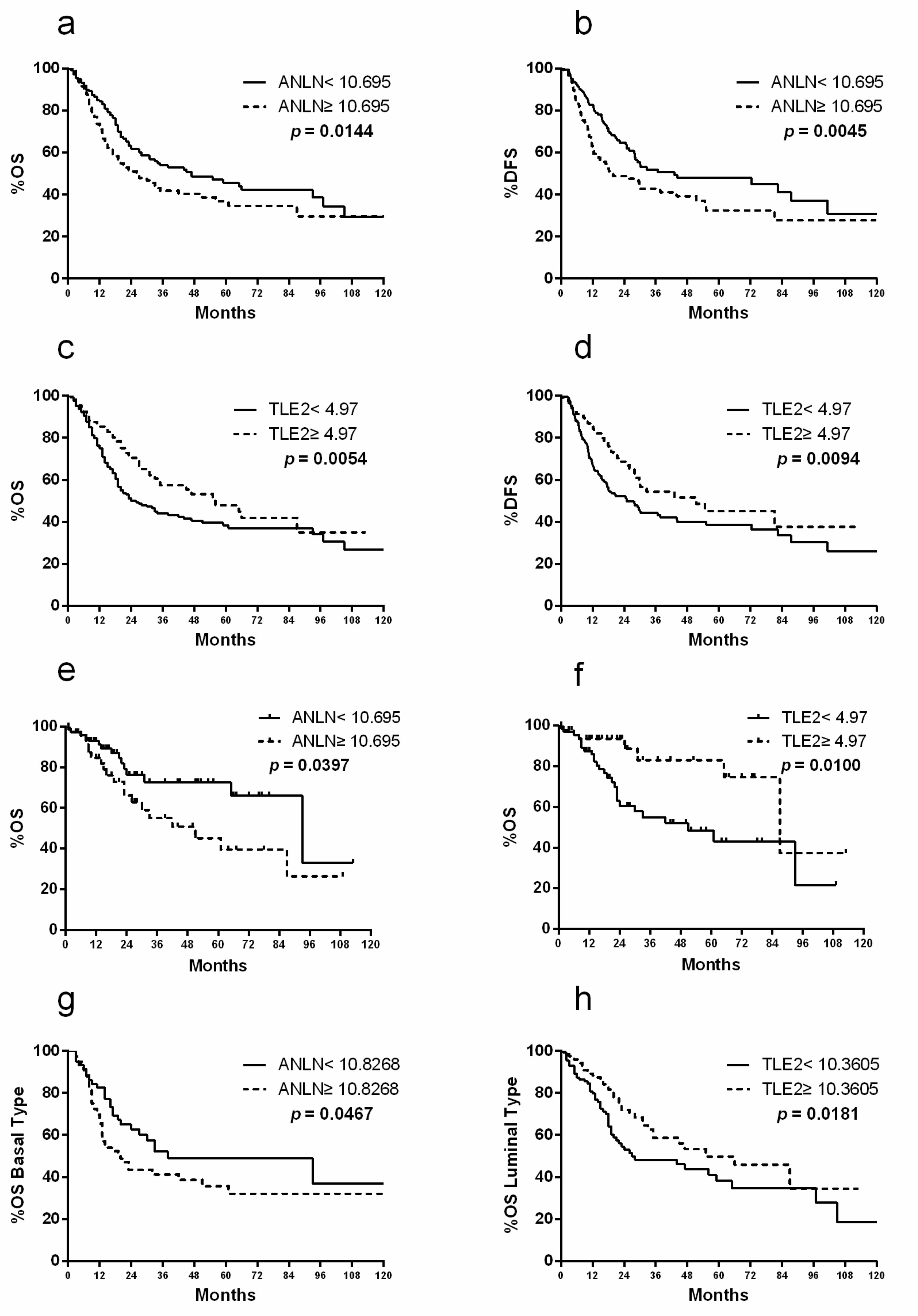
2.2. ANLN and TLE2 as Risk Markers for Prognostic Prediction After RC
2.3. Gene Expression Profiling of ANLN and TLE2 in Comparison with other Tumor Entities
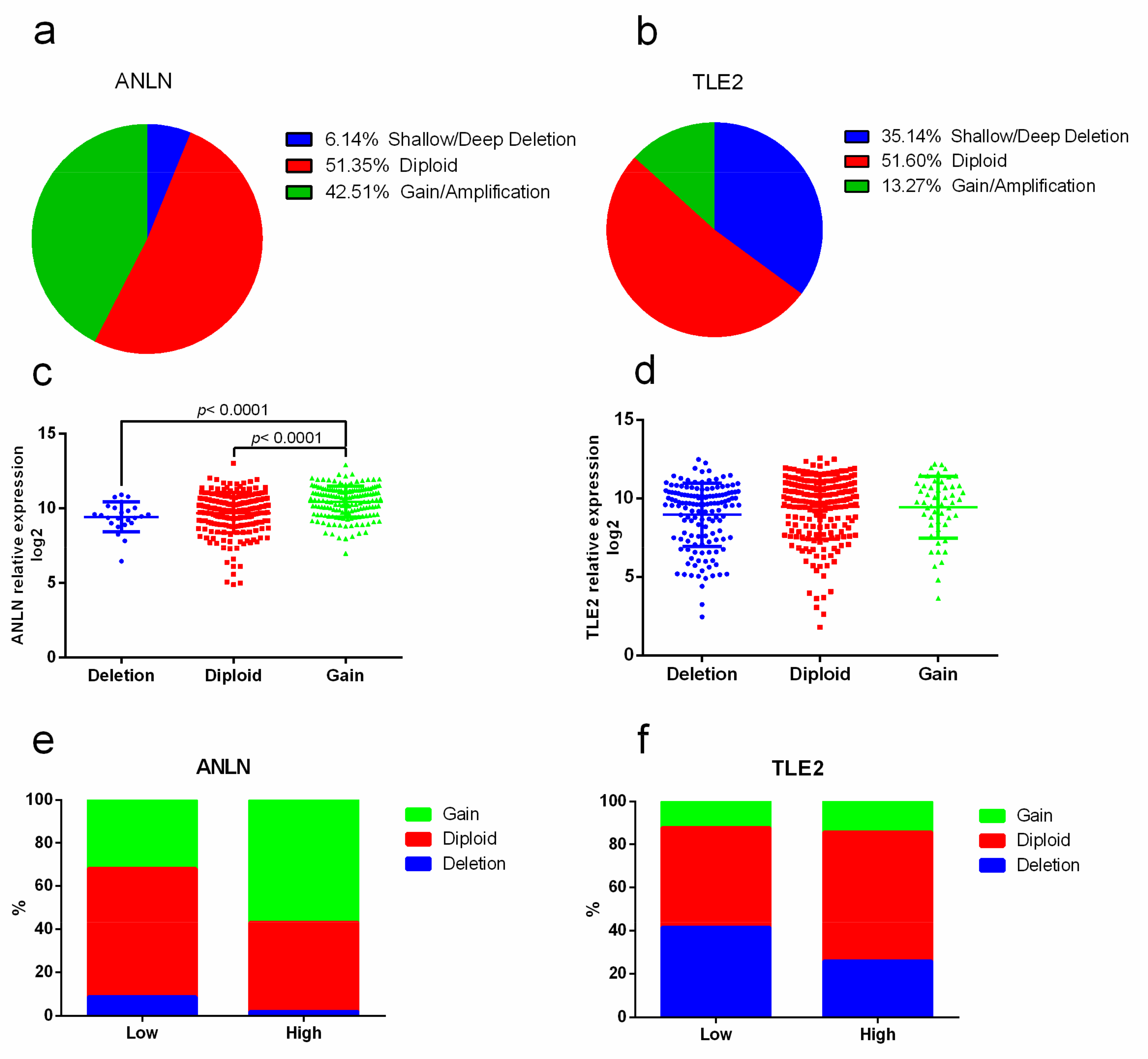
2.4. Association of Copy-number Alterations with ANLN and TLE2 Gene Expression
2.5. Correlation with Signaling Pathways and Therapeutic Targets in Bladder Cancer
2.6. Molecular Subtype Specificity of ANLN and TLE2
3. Discussion
4. Materials and Methods
4.1. Patients and Tissue Samples
4.2. Database
4.3. Cell Lines
4.4. RNA Extraction and qRT-PCR
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA A Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef]
- Babjuk, M.; Bohle, A.; Burger, M.; Capoun, O.; Cohen, D.; Comperat, E.M.; Hernandez, V.; Kaasinen, E.; Palou, J.; Roupret, M.; et al. EAU Guidelines on Non-Muscle-invasive Urothelial Carcinoma of the Bladder: Update 2016. Eur. Urol. 2017, 71, 447–461. [Google Scholar] [CrossRef]
- Shah, J.B.; McConkey, D.J.; Dinney, C.P. New strategies in muscle-invasive bladder cancer: On the road to personalized medicine. Clin. Cancer Res. 2011, 17, 2608–2612. [Google Scholar] [CrossRef] [PubMed]
- Svatek, R.S.; Shariat, S.F.; Novara, G.; Skinner, E.C.; Fradet, Y.; Bastian, P.J.; Kamat, A.M.; Kassouf, W.; Karakiewicz, P.I.; Fritsche, H.M.; et al. Discrepancy between clinical and pathological stage: External validation of the impact on prognosis in an international radical cystectomy cohort. BJU. Int. 2011, 107, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.G.; Kim, J.; Al-Ahmadie, H.; Bellmunt, J.; Guo, G.; Cherniack, A.D.; Hinoue, T.; Laird, P.W.; Hoadley, K.A.; Akbani, R. Comprehensive molecular characterization of muscle-invasive bladder cancer. Cell 2017, 171, 540–556. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Porten, S.; Kim, S.; Willis, D.; Plimack, E.R.; Hoffman-Censits, J.; Roth, B.; Cheng, T.; Tran, M.; Lee, I.-L. Identification of distinct basal and luminal subtypes of muscle-invasive bladder cancer with different sensitivities to frontline chemotherapy. Cancer Cell 2014, 25, 152–165. [Google Scholar] [CrossRef]
- Martens-Uzunova, E.S.; Böttcher, R.; Croce, C.M.; Jenster, G.; Visakorpi, T.; Calin, G.A. Long noncoding RNA in prostate, bladder, and kidney cancer. Eur. Urol. 2014, 65, 1140–1151. [Google Scholar] [CrossRef]
- Kandimalla, R.; Van Tilborg, A.A.; Zwarthoff, E.C. DNA methylation-based biomarkers in bladder cancer. Nat. Rev. Urol. 2013, 10, 327–335. [Google Scholar] [CrossRef]
- Miller, K.G.; Field, C.M.; Alberts, B.M. Actin-binding proteins from Drosophila embryos: A complex network of interacting proteins detected by F-actin affinity chromatography. J. Cell Biol. 1989, 109, 2963–2975. [Google Scholar] [CrossRef]
- Hall, P.A.; Todd, C.B.; Hyland, P.L.; McDade, S.S.; Grabsch, H.; Dattani, M.; Hillan, K.J.; Russell, S.E. The septin-binding protein anillin is overexpressed in diverse human tumors. Clin. Cancer Res. 2005, 11, 6780–6786. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, J.; Zhong, M.-Z.; Huang, J.; Hu, Y.-P.; Feng, D.-Y.; Zhou, Z.-J.; Luo, X.; Liu, Z.-Q.; Jiang, W.-Z. Overexpression of ANLN contributed to poor prognosis of anthracycline-based chemotherapy in breast cancer patients. Cancer Chemother. Pharmacol. 2017, 79, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Schiewek, J.; Schumacher, U.; Lange, T.; Joosse, S.A.; Wikman, H.; Pantel, K.; Mikhaylova, M.; Kneussel, M.; Linder, S.; Schmalfeldt, B. Clinical relevance of cytoskeleton associated proteins for ovarian cancer. J. Cancer Res. Clin. Oncol. 2018, 144, 2195–2205. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Shen, W.; Cui, L.; Chen, W.; Hu, X.; Fu, J. Overexpression of Anillin (ANLN) is correlated with colorectal cancer progression and poor prognosis. Cancer Biomark. 2016, 16, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, C.; Daigo, Y.; Ishikawa, N.; Kato, T.; Hayama, S.; Ito, T.; Tsuchiya, E.; Nakamura, Y. ANLN plays a critical role in human lung carcinogenesis through the activation of RHOA and by involvement in the phosphoinositide 3-kinase/AKT pathway. Cancer Res. 2005, 65, 11314–11325. [Google Scholar] [CrossRef]
- Olakowski, M.; Tyszkiewicz, T.; Jarzab, M.; Krol, R.; Oczko-Wojciechowska, M.; Kowalska, M.; Kowal, M.; Gala, G.M.; Kajor, M.; Lange, D.; et al. NBL1 and anillin (ANLN) genes over-expression in pancreatic carcinoma. Folia Histochem. Cytobiol. 2009, 47, 249–255. [Google Scholar] [CrossRef]
- Zeng, S.; Yu, X.; Ma, C.; Song, R.; Zhang, Z.; Zi, X.; Chen, X.; Wang, Y.; Yu, Y.; Zhao, J. Transcriptome sequencing identifies ANLN as a promising prognostic biomarker in bladder urothelial carcinoma. Sci. Rep. 2017, 7, 3151. [Google Scholar] [CrossRef]
- Grbavec, D.; Lo, R.; Liu, Y.; Stifani, S. Transducin-like Enhancer of split 2, a mammalian homologue of Drosophila Groucho, acts as a transcriptional repressor, interacts with Hairy/Enhancer of split proteins, and is expressed during neuronal development. Eur. J. Biochem. 1998, 258, 339–349. [Google Scholar] [CrossRef]
- Hoffman, B.G.; Zavaglia, B.; Beach, M.; Helgason, C.D. Expression of Groucho/TLE proteins during pancreas development. BMC Dev. Biol. 2008, 8, 81. [Google Scholar] [CrossRef]
- He, Z.; Liu, Y.; Liang, D.; Wang, Z.; Robertson, E.S.; Lan, K. Cellular corepressor TLE2 inhibits replication-and-transcription-activator-mediated transactivation and lytic reactivation of Kaposi’s sarcoma-associated herpesvirus. J. Virol. 2010, 84, 2047–2062. [Google Scholar] [CrossRef]
- Pandi, N.S.; Manimuthu, M.; Harunipriya, P.; Murugesan, M.; Asha, G.V.; Rajendran, S. In silico analysis of expression pattern of a Wnt/beta-catenin responsive gene ANLN in gastric cancer. Gene 2014, 545, 23–29. [Google Scholar] [CrossRef]
- Cavallo, R.A.; Cox, R.T.; Moline, M.M.; Roose, J.; Polevoy, G.A.; Clevers, H.; Peifer, M.; Bejsovec, A. Drosophila Tcf and Groucho interact to repress Wingless signalling activity. Nature 1998, 395, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Cinnamon, E.; Paroush, Z. Context-dependent regulation of Groucho/TLE-mediated repression. Curr. Opin. Genet. Dev. 2008, 18, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.-I.; Chen, W.T.; Li, C.-F.; Li, C.-C.; Li, W.-M.; Huang, C.-N.; Yeh, H.-C.; Ke, H.-L.; Wu, W.-J.; Chai, C.-Y. Subcellular localisation of anillin is associated with different survival outcomes in upper urinary tract urothelial carcinoma. J. Clin. Pathol. 2015, 68, 1026–1032. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Nguyen, L.H.; Zhou, K.; Tu, H.-C.; Sehgal, A.; Nassour, I.; Li, L.; Gopal, P.; Goodman, J.; Singal, A.G. Knockdown of anillin actin binding protein blocks cytokinesis in hepatocytes and reduces liver tumor development in mice without affecting regeneration. Gastroenterology 2018, 154, 1421–1434. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, M.; Feng, C.; Gao, P.; Ding, G.; Zhou, Z.; Jiang, H.; Wu, Z.; Ding, Q. Prognostic value of Ki67 and p63 expressions in bladder cancer patients who underwent radical cystectomy. Int. Urol. Nephrol. 2016, 48, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.; Sabichi, A.L.; Gill, J.; Lee, I.-L.; Adegboyega, P.; Dai, M.S.; Loganantharaj, R.; Trutschl, M.; Cvek, U.; Clifford, J.L. Identification of genes correlated with early-stage bladder cancer progression. Cancer Prev. Res. 2010, 3, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Breyer, J.; Wirtz, R.M.; Laible, M.; Schlombs, K.; Erben, P.; Kriegmair, M.C.; Stoehr, R.; Eidt, S.; Denzinger, S.; Burger, M. ESR1, ERBB2, and Ki67 mRNA expression predicts stage and grade of non-muscle-invasive bladder carcinoma (NMIBC). Virchows Arch. 2016, 469, 547–552. [Google Scholar] [CrossRef]
- Martini, T.; Heinkele, J.; Mayr, R.; Weis, C.-A.; Wezel, F.; Wahby, S.; Eckstein, M.; Schnöller, T.; Breyer, J.; Wirtz, R. Predictive value of lymphangiogenesis and proliferation markers on mRNA level in urothelial carcinoma of the bladder after radical cystectomy. In Urologic Oncology: Seminars and Original Investigations; Elsevier: Amsterdam, The Netherlands, 2018; pp. e530-19–e530-27. [Google Scholar]
- Rinaldetti, S.; Wirtz, R.M.; Worst, T.S.; Eckstein, M.; Weiss, C.A.; Breyer, J.; Otto, W.; Bolenz, C.; Hartmann, A.; Erben, P. FOXM1 predicts overall and disease specific survival in muscle-invasive urothelial carcinoma and presents a differential expression between bladder cancer subtypes. Oncotarget 2017, 8, 47595–47606. [Google Scholar] [CrossRef]
- Pei, Z.; Du, X.; Song, Y.; Fan, L.; Li, F.; Gao, Y.; Wu, R.; Chen, Y.; Li, W.; Zhou, H. Down-regulation of lncRNA CASC2 promotes cell proliferation and metastasis of bladder cancer by activation of the Wnt/β-catenin signaling pathway. Oncotarget 2017, 8, 18145–18153. [Google Scholar] [CrossRef]
- Du, H.-F.; Ou, L.-P.; Lv, C.-K.; Yang, X.; Song, X.-D.; Fan, Y.-R.; Wu, X.-H.; Luo, C.-L. Expression of hepaCAM inhibits bladder cancer cell proliferation via a Wnt/β-catenin-dependent pathway in vitro and in vivo. Cancer Biol. Ther. 2015, 16, 1502–1513. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, L.; Wang, L.; Kazobinka, G.; Zhang, X.; Han, X.; Li, B.; Hou, T. HBO1 promotes cell proliferation in bladder cancer via activation of Wnt/β-catenin signaling. Mol. Carcinog. 2018, 57, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Kurnit, K.C.; Kim, G.N.; Fellman, B.M.; Urbauer, D.L.; Mills, G.B.; Zhang, W.; Broaddus, R.R. CTNNB1 (beta-catenin) mutation identifies low grade, early stage endometrial cancer patients at increased risk of recurrence. Mod. Pathol. 2017, 30, 1032–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crippa, S.; Ancey, P.B.; Vazquez, J.; Angelino, P.; Rougemont, A.L.; Guettier, C.; Zoete, V.; Delorenzi, M.; Michielin, O.; Meylan, E. Mutant CTNNB1 and histological heterogeneity define metabolic subtypes of hepatoblastoma. EMBO Mol. Med. 2017, 9, 1589–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahadova, A.; von Knebel Doeberitz, M.; Bläker, H.; Kloor, M. CTNNB1-mutant colorectal carcinomas with immediate invasive growth: A model of interval cancers in Lynch syndrome. Fam. Cancer 2016, 15, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Ai, R.; Sun, Y.; Guo, Z.; Wei, W.; Zhou, L.; Liu, F.; Hendricks, D.T.; Xu, Y.; Zhao, X. NDRG1 overexpression promotes the progression of esophageal squamous cell carcinoma through modulating Wnt signaling pathway. Cancer Biol. Ther. 2016, 17, 943–954. [Google Scholar] [CrossRef] [Green Version]
- Hambly, N.; Shimbori, C.; Kolb, M. Molecular classification of idiopathic pulmonary fibrosis: Personalized medicine, genetics and biomarkers. Respirology 2015, 20, 1010–1022. [Google Scholar] [CrossRef] [Green Version]
- Pietzak, E.J.; Bagrodia, A.; Cha, E.K.; Drill, E.N.; Iyer, G.; Isharwal, S.; Ostrovnaya, I.; Baez, P.; Li, Q.; Berger, M.F. Next-generation sequencing of nonmuscle invasive bladder cancer reveals potential biomarkers and rational therapeutic targets. Eur. Urol. 2017, 72, 952–959. [Google Scholar] [CrossRef]
- Hedegaard, J.; Lamy, P.; Nordentoft, I.; Algaba, F.; Høyer, S.; Ulhøi, B.P.; Vang, S.; Reinert, T.; Hermann, G.G.; Mogensen, K. Comprehensive transcriptional analysis of early-stage urothelial carcinoma. Cancer Cell 2016, 30, 27–42. [Google Scholar] [CrossRef]
- Damrauer, J.S.; Hoadley, K.A.; Chism, D.D.; Fan, C.; Tiganelli, C.J.; Wobker, S.E.; Yeh, J.J.; Milowsky, M.I.; Iyer, G.; Parker, J.S. Intrinsic subtypes of high-grade bladder cancer reflect the hallmarks of breast cancer biology. Proc. Natl. Acad. Sci. USA. 2014, 111, 3110–3115. [Google Scholar] [CrossRef] [Green Version]
- Fedele, M.; Cerchia, L.; Chiappetta, G. The Epithelial-to-Mesenchymal Transition in Breast Cancer: Focus on Basal-Like Carcinomas. Cancers 2017, 9. [Google Scholar] [CrossRef]
- Li, Y.; Yang, K.; Li, K.; Liu, H.; Zhao, S.; Jiao, M.; Fu, X. Clinical and molecular characteristics of bladder urothelial carcinoma subtypes. J. Cell. Biochem. 2019, 120, 9956–9963. [Google Scholar] [CrossRef] [PubMed]
- Eckstein, M.; Wirtz, R.M.; Gross-Weege, M.; Breyer, J.; Otto, W.; Stoehr, R.; Sikic, D.; Keck, B.; Eidt, S.; Burger, M.; et al. mRNA-Expression of KRT5 and KRT20 Defines Distinct Prognostic Subgroups of Muscle-Invasive Urothelial Bladder Cancer Correlating with Histological Variants. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, Y.W.; Hajjar, J.; Hwu, P.; Naing, A. Targeting the indoleamine 2,3-dioxygenase pathway in cancer. J. Immunother. Cancer 2015, 3, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tumeh, P.C.; Harview, C.L.; Yearley, J.H.; Shintaku, I.P.; Taylor, E.J.; Robert, L.; Chmielowski, B.; Spasic, M.; Henry, G.; Ciobanu, V. PD-1 blockade induces responses by inhibiting adaptive immune resistance. Nature 2014, 515, 568–571. [Google Scholar] [CrossRef]
- Aggen, D.H.; Drake, C.G. Biomarkers for immunotherapy in bladder cancer: A moving target. J. Immunother. Cancer 2017, 5, 94. [Google Scholar] [CrossRef]
- Paner, G.P.; Stadler, W.M.; Hansel, D.E.; Montironi, R.; Lin, D.W.; Amin, M.B. Updates in the Eighth Edition of the Tumor-Node-Metastasis Staging Classification for Urologic Cancers. Eur. Urol. 2018, 73, 560–569. [Google Scholar] [CrossRef]
- Woldu, S.L.; Bagrodia, A.; Lotan, Y. Guideline of guidelines: Non-muscle-invasive bladder cancer. BJU Int. 2017, 119, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Tramm, T.; Sørensen, B.S.; Overgaard, J.; Alsner, J. Optimal reference genes for normalization of qRT-PCR data from archival formalin-fixed, paraffin-embedded breast tumors controlling for tumor cell content and decay of mRNA. Diagn. Mol. Pathol. 2013, 22, 181–187. [Google Scholar] [CrossRef]
- Breyer, J.; Wirtz, R.M.; Otto, W.; Erben, P.; Kriegmair, M.C.; Stoehr, R.; Eckstein, M.; Eidt, S.; Denzinger, S.; Burger, M. In stage pT1 non-muscle-invasive bladder cancer (NMIBC), high KRT20 and low KRT5 mRNA expression identify the luminal subtype and predict recurrence and survival. Virchows Arch. 2017, 470, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, M.R.D.; Petricelli, C.D.; Alexandre, S.M.; Paschoal, A.; Araujo Júnior, E.; Nakamura, M.U. Determination of a cutoff value for pelvic floor distensibility using the Epi-no balloon to predict perineal integrity in vaginal delivery: ROC curve analysis. Prospective observational single cohort study. Sao Paulo Med. J. 2016, 134, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Features | n | |
|---|---|---|
| Age | <70 | 23 |
| ≥70 | 37 | |
| Gender | Male | 47 |
| Female | 13 | |
| Grade | Low | 5 |
| High | 55 | |
| Stage | T1 | 6 |
| T2 | 11 | |
| T3 | 28 | |
| T4 | 15 | |
| Lymph node metastasis | Negative | 37 |
| Positive | 15 | |
| Factor | Univariate | Multivariate | ||
|---|---|---|---|---|
| HR (95% CI) | p Value | HR (95% CI) | p Value | |
| Diagnosis Age | ||||
| <70 vs. ≥70 | 0.584 (0.269–1.269) | 0.174 | − | − |
| Gender | ||||
| Male vs. Female | 1.128 (0.405–3.140) | 0.957 | − | − |
| Stage | ||||
| T1/T2 vs. T3/4 | 0.168 (0.015–1.832) | 0.128 | − | − |
| LVI | ||||
| Negative vs. Positive | 0.542 (0.359–0.819) | 0.004 | − | − |
| Lymph node Statues | ||||
| Negative vs. Positive | 0.549 (0.371–0.813) | 0.003 | 0.612 (0.399–0.938) | 0.024 |
| ANLN | ||||
| Low vs. High | 0.220 (0.084–0.575) | 0.002 | 0.328 (0.114–0.945) | 0.039 |
| TLE2 | ||||
| Low vs. High | 0.305 (0.121–0.769) | 0.012 | 0.172 (0.057–0.519) | 0.002 |
| Factor | Univariate | Multivariate | ||
|---|---|---|---|---|
| HR (95% CI) | p Value | HR (95% CI) | p Value | |
| Diagnosis Age | ||||
| <70 vs. ≥70 | 1.260 (0.931–1.705) | 0.134 | − | − |
| Gender | ||||
| Male vs. Female | 1.257 (0.902–1.751) | 0.177 | − | − |
| Smoking Status | ||||
| No vs. Yes | 1.335 (0.940–1.897) | 0.106 | − | − |
| Stage | ||||
| T2 vs. T3/4 | 1.950 (1.393–2.731) | <0.001 | 1.646 (1.156–2.342) | 0.006 |
| Lymph node Statues | ||||
| Negative vs. Positive | 2.145 (1.596–2.883) | <0.001 | 1.989 (1.461–2.707) | <0.001 |
| ANLN | ||||
| Low vs. High | 1.439 (1.070–1.934) | 0.016 | 1.438 (1.064–1.943) | 0.018 |
| TLE2 | ||||
| Low vs. High | 0.636 (0.460–0.880) | 0.006 | 1.415 (1.015–1.973) | 0.040 |
| Correlated Gene | ANLN | TLE2 | |||
|---|---|---|---|---|---|
| Correlation Coefficient | p Value | Correlation Coefficient | p Value | ||
| Cell proliferation | CDK1 | 0.594 | 3.98 × 10−40 | −0.338 | 2.63 × 10−12 |
| RACGAP1 | 0.725 | 1.41 × 10−67 | −0.451 | 9.46 × 10−22 | |
| MKI67 | 0.711 | 7.56 × 10−64 | −0.396 | 1.07 × 10−16 | |
| FOXM1 | 0.688 | 2.98 × 10−58 | −0.4 | 4.46 × 10−17 | |
| Notch signaling | NOTCH1 | 0.109 | 0.027294 | −0.19 | 0.000115 |
| RBPJ | −0.196 | 6.6 × 10−5 | 0.05 | 0.311468 | |
| Wnt signaling | TCF7 | 0.067 | 0.175097 | 0.006 | 0.89689 |
| TCF7L1 | 0.162 | 0.001066 | −0.267 | 4.75 × 10−8 | |
| TCF7L2 | −0.186 | 0.000164 | 0.23 | 2.84 × 10−6 | |
| LEF1 | −0.101 | 0.041347 | −0.026 | 0.596286 | |
| CTNNB1 | 0.237 | 1.29 × 10−6 | −0.276 | 1.6 × 10−8 | |
| Hormone receptor signaling | AR | −0.185 | 0.000171 | 0.388 | 4.9 × 10−16 |
| ESR1 | −0.003 | 0.944815 | −0.048 | 0.332592 | |
| ESR2 | −0.191 | 0.00011 | 0.334 | 4.33 × 10−12 | |
| FOXA1 | −0.38 | 1.89 × 10−15 | 0.505 | 9.11 × 10−28 | |
| GATA3 | −0.403 | 2.33 × 10−17 | 0.65 | 2.80 × 10−50 | |
| Therapeutic targets | EGFR | 0.53 | 7.73 × 10−31 | −0.368 | 1.78 × 10−14 |
| ERBB2 | −0.35 | 3.44 × 10−13 | 0.613 | 1.96 × 10−43 | |
| FGFR3 | −0.283 | 6.52 × 10−9 | 0.156 | 0.001575 | |
| PIK3CA | 0.426 | 2.19 × 10−19 | −0.264 | 6.34 × 10−8 | |
| CDK4 | 0.207 | 2.68 × 10−5 | −0.267 | 4.69 × 10−8 | |
| HRAS | −0.023 | 0.648147 | −0.183 | 0.000202 | |
| PDCD1 | 0.0881 | 0.0754 | −0.313 | 1.07 × 10−10 | |
| PD-L1 | 0.414 | 2.89 × 10−18 | −0.419 | 1.07 × 10−18 | |
| CTLA4 | 0.0963 | 0.052 | −0.363 | 3.71 × 10−14 | |
| EZH2 | 0.446 | 2.51 × 10−21 | −0.162 | 0.001009 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Nitschke, K.; Heinkele, J.; Weis, C.-A.; Worst, T.S.; Eckstein, M.; Porubsky, S.; Erben, P. ANLN and TLE2 in Muscle Invasive Bladder Cancer: A Functional and Clinical Evaluation Based on In Silico and In Vitro Data. Cancers 2019, 11, 1840. https://doi.org/10.3390/cancers11121840
Wu S, Nitschke K, Heinkele J, Weis C-A, Worst TS, Eckstein M, Porubsky S, Erben P. ANLN and TLE2 in Muscle Invasive Bladder Cancer: A Functional and Clinical Evaluation Based on In Silico and In Vitro Data. Cancers. 2019; 11(12):1840. https://doi.org/10.3390/cancers11121840
Chicago/Turabian StyleWu, Sheng, Katja Nitschke, Jakob Heinkele, Cleo-Aron Weis, Thomas Stefan Worst, Markus Eckstein, Stefan Porubsky, and Philipp Erben. 2019. "ANLN and TLE2 in Muscle Invasive Bladder Cancer: A Functional and Clinical Evaluation Based on In Silico and In Vitro Data" Cancers 11, no. 12: 1840. https://doi.org/10.3390/cancers11121840
APA StyleWu, S., Nitschke, K., Heinkele, J., Weis, C. -A., Worst, T. S., Eckstein, M., Porubsky, S., & Erben, P. (2019). ANLN and TLE2 in Muscle Invasive Bladder Cancer: A Functional and Clinical Evaluation Based on In Silico and In Vitro Data. Cancers, 11(12), 1840. https://doi.org/10.3390/cancers11121840