Use of Dendritic Cell Receptors as Targets for Enhancing Anti-Cancer Immune Responses


Abstract
:1. Introduction
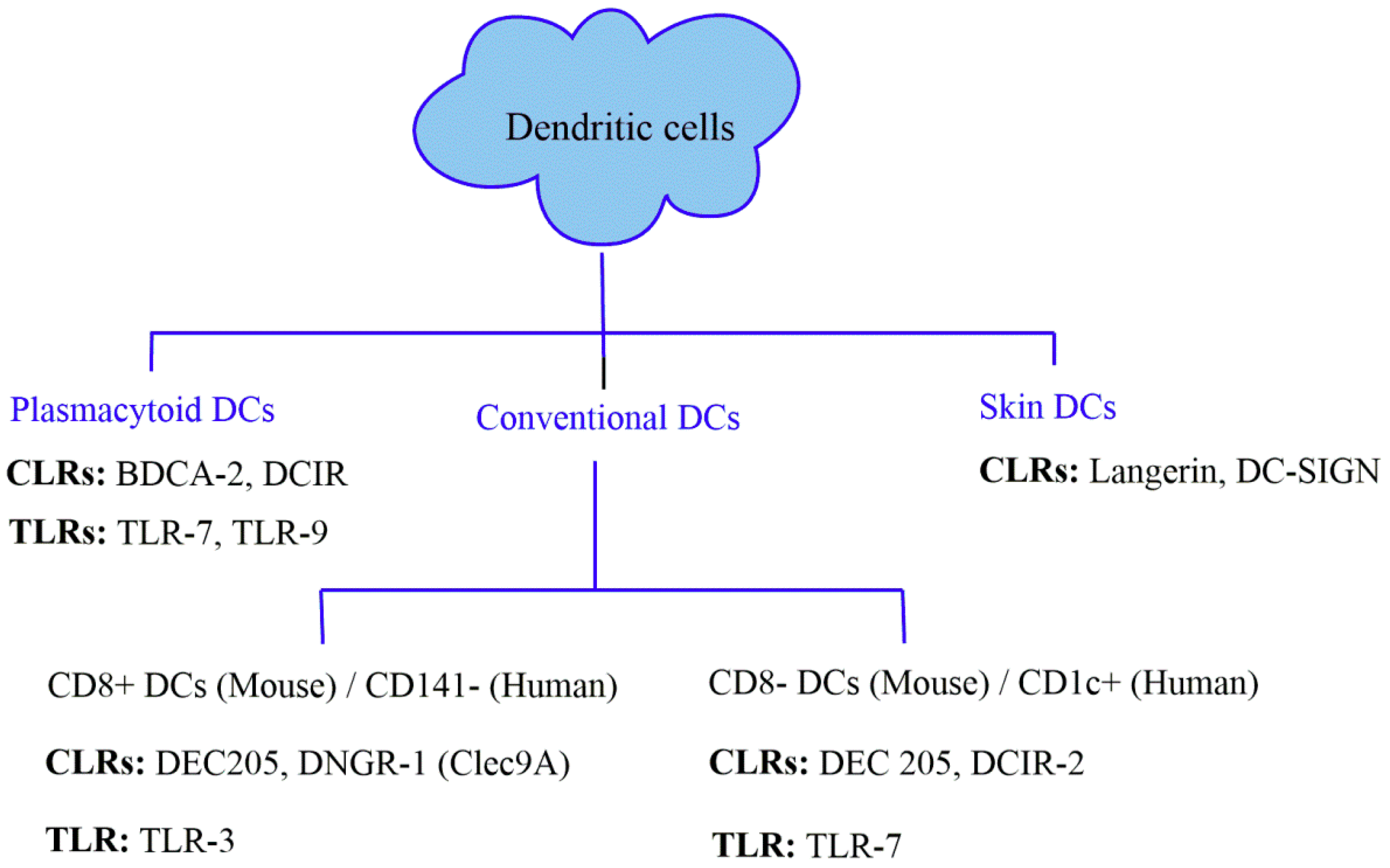
2. Dendritic Cell Subsets and Receptors
3. Targeting Different DCs Receptors for Vaccine Development
3.1. C-type Lectin Receptors
3.1.1. Mannose Receptor (MR)
3.1.2. DEC-205
3.1.3. Dectin-1 and 2
3.1.4. DC-SIGN
3.1.5. DNGR-1
3.2. Fc Receptors
3.3. Targeting Methodologies
4. Conclusions
Funding
Conflicts of Interest
References
- Ljunggren, H.G. Dendritic cells, dendritic cell-based vaccines and Ralph Steinman. J. Intern. Med. 2012, 271, 174–176. [Google Scholar] [CrossRef]
- Steinman, R.M.; Witmer, M.D. Lymphoid dendritic cells are potent stimulators of the primary mixed leukocyte reaction in mice. Proc. Natl. Acad. Sci. USA 1978, 75, 5132–5136. [Google Scholar] [CrossRef]
- McMenamin, P.G. Distribution and phenotype of dendritic cells and resident tissue macrophages in the dura mater, leptomeninges, and choroid plexus of the rat brain as demonstrated in wholemount preparations. J. Comp. Neurol. 1999, 405, 553–562. [Google Scholar] [CrossRef]
- Reis e Sousa, C. Activation of dendritic cells: Translating innate into adaptive immunity. Curr. Opin. Immunol. 2004, 16, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Blum, J.S.; Wearsch, P.A.; Cresswell, P. Pathways of antigen processing. Annu. Rev. Immunol. 2013, 31, 443–473. [Google Scholar] [CrossRef] [PubMed]
- Germain, R.N. MHC-dependent antigen processing and peptide presentation: Providing ligands for T lymphocyte activation. Cell 1994, 76, 287–299. [Google Scholar] [CrossRef]
- Elliott, T. Transporter associated with antigen processing. Adv. Immunol. 1997, 65, 47–109. [Google Scholar] [PubMed]
- Momburg, F.; Hammerling, G.J. Generation and TAP-mediated transport of peptides for major histocompatibility complex class I molecules. Adv. Immunol. 1998, 68, 191–256. [Google Scholar] [PubMed]
- Paulsson, K.; Wang, P. Chaperones and folding of MHC class I molecules in the endoplasmic reticulum. Biochim. Biophys. Acta 2003, 1641, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Williams, D.B. Assembly of MHC class I molecules within the endoplasmic reticulum. Immunol. Res. 2006, 35, 151–162. [Google Scholar] [CrossRef]
- Chen, P.; Liu, X.; Sun, Y.; Zhou, P.; Wang, Y.; Zhang, Y. Dendritic cell targeted vaccines: Recent progresses and challenges. Hum. Vaccin. Immunother. 2016, 12, 612–622. [Google Scholar] [CrossRef]
- Berger, A. Th1 and Th2 responses: What are they? BMJ 2000, 321, 424. [Google Scholar] [CrossRef] [PubMed]
- Wohlleber, D.; Kashkar, H.; Gartner, K.; Frings, M.K.; Odenthal, M.; Hegenbarth, S.; Borner, C.; Arnold, B.; Hammerling, G.; Nieswandt, B.; et al. TNF-induced target cell killing by CTL activated through cross-presentation. Cell Rep. 2012, 2, 478–487. [Google Scholar] [CrossRef]
- Keler, T.; He, L.; Ramakrishna, V.; Champion, B. Antibody-targeted vaccines. Oncogene 2007, 26, 3758–3767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, A.; Panozza, S.E.; Macri, C.; Dumont, C.; Li, J.; Liu, H.; Segura, E.; Vega-Ramos, J.; Gupta, N.; Caminschi, I.; et al. Criteria for dendritic cell receptor selection for efficient antibody-targeted vaccination. J. Immunol. 2015, 194, 2696–2705. [Google Scholar] [CrossRef]
- Tacken, P.J.; Figdor, C.G. Targeted antigen delivery and activation of dendritic cells in vivo: Steps towards cost effective vaccines. Semin. Immunol. 2011, 23, 12–20. [Google Scholar] [CrossRef] [PubMed]
- McKenna, K.; Beignon, A.S.; Bhardwaj, N. Plasmacytoid dendritic cells: Linking innate and adaptive immunity. J. Virol. 2005, 79, 17–27. [Google Scholar] [CrossRef]
- Reizis, B.; Bunin, A.; Ghosh, H.S.; Lewis, K.L.; Sisirak, V. Plasmacytoid dendritic cells: Recent progress and open questions. Annu. Rev. Immunol. 2011, 29, 163–183. [Google Scholar] [CrossRef]
- Swiecki, M.; Colonna, M. The multifaceted biology of plasmacytoid dendritic cells. Nat. Rev. Immunol. 2015, 15, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Young, L.J.; Wilson, N.S.; Schnorrer, P.; Proietto, A.; ten Broeke, T.; Matsuki, Y.; Mount, A.M.; Belz, G.T.; O’Keeffe, M.; Ohmura-Hoshino, M.; et al. Differential MHC class II synthesis and ubiquitination confers distinct antigen-presenting properties on conventional and plasmacytoid dendritic cells. Nat. Immunol. 2008, 9, 1244–1252. [Google Scholar] [CrossRef]
- Hoeffel, G.; Ripoche, A.C.; Matheoud, D.; Nascimbeni, M.; Escriou, N.; Lebon, P.; Heshmati, F.; Guillet, J.G.; Gannage, M.; Caillat-Zucman, S.; et al. Antigen crosspresentation by human plasmacytoid dendritic cells. Immunity 2007, 27, 481–492. [Google Scholar] [CrossRef]
- Ochando, J.C.; Homma, C.; Yang, Y.; Hidalgo, A.; Garin, A.; Tacke, F.; Angeli, V.; Li, Y.; Boros, P.; Ding, Y.; et al. Alloantigen-presenting plasmacytoid dendritic cells mediate tolerance to vascularized grafts. Nat. Immunol. 2006, 7, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Ribas, D.; Adema, G.J.; Winkels, G.; Klasen, I.S.; Punt, C.J.; Figdor, C.G.; de Vries, I.J. Plasmacytoid dendritic cells of melanoma patients present exogenous proteins to CD4+ T cells after Fc gamma RII-mediated uptake. J. Exp. Med. 2006, 203, 1629–1635. [Google Scholar] [CrossRef] [PubMed]
- Dzionek, A.; Fuchs, A.; Schmidt, P.; Cremer, S.; Zysk, M.; Miltenyi, S.; Buck, D.W.; Schmitz, J. BDCA-2, BDCA-3, and BDCA-4: Three markers for distinct subsets of dendritic cells in human peripheral blood. J. Immunol. 2000, 165, 6037–6046. [Google Scholar] [CrossRef]
- Segura, E.; Valladeau-Guilemond, J.; Donnadieu, M.H.; Sastre-Garau, X.; Soumelis, V.; Amigorena, S. Characterization of resident and migratory dendritic cells in human lymph nodes. J. Exp. Med. 2012, 209, 653–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merad, M.; Sathe, P.; Helft, J.; Miller, J.; Mortha, A. The dendritic cell lineage: Ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu. Rev. Immunol. 2013, 31, 563–604. [Google Scholar] [CrossRef]
- Macri, C.; Dumont, C.; Johnston, A.P.; Mintern, J.D. Targeting dendritic cells: A promising strategy to improve vaccine effectiveness. Clin. Transl. Immunol. 2016, 5, e66. [Google Scholar] [CrossRef]
- Caminschi, I.; Proietto, A.I.; Ahmet, F.; Kitsoulis, S.; Shin Teh, J.; Lo, J.C.; Rizzitelli, A.; Wu, L.; Vremec, D.; van Dommelen, S.L.; et al. The dendritic cell subtype-restricted C-type lectin Clec9A is a target for vaccine enhancement. Blood 2008, 112, 3264–3273. [Google Scholar] [CrossRef] [Green Version]
- Shortman, K.; Heath, W.R. The CD8+ dendritic cell subset. Immunol. Rev. 2010, 234, 18–31. [Google Scholar] [CrossRef]
- den Haan, J.M.; Lehar, S.M.; Bevan, M.J. CD8(+) but not CD8(-) dendritic cells cross-prime cytotoxic T cells in vivo. J. Exp. Med. 2000, 192, 1685–1696. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.D.; Diebold, S.S.; Slack, E.M.; Tomizawa, H.; Hemmi, H.; Kaisho, T.; Akira, S.; Reis e Sousa, C. Toll-like receptor expression in murine DC subsets: Lack of TLR7 expression by CD8 alpha+ DC correlates with unresponsiveness to imidazoquinolines. Eur. J. Immunol. 2003, 33, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Signaling to NF-kappaB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef]
- Zhang, Y.; El-Far, M.; Dupuy, F.P.; Abdel-Hakeem, M.S.; He, Z.; Procopio, F.A.; Shi, Y.; Haddad, E.K.; Ancuta, P.; Sekaly, R.P.; et al. HCV RNA activates APCs via TLR7/TLR8 while virus selectively stimulates macrophages without inducing antiviral responses. Sci. Rep. 2016, 6, 29447. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D.H. In vivo function of Langerhans cells and dermal dendritic cells. Trends Immunol. 2010, 31, 446–451. [Google Scholar] [CrossRef]
- van Kooyk, Y.; Rabinovich, G.A. Protein-glycan interactions in the control of innate and adaptive immune responses. Nat. Immunol. 2008, 9, 593–601. [Google Scholar] [CrossRef]
- Sancho, D.; Reis e Sousa, C. Sensing of cell death by myeloid C-type lectin receptors. Curr. Opin. Immunol. 2013, 25, 46–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dambuza, I.M.; Brown, G.D. C-type lectins in immunity: Recent developments. Curr. Opin. Immunol. 2015, 32, 21–27. [Google Scholar] [CrossRef]
- Robinson, M.J.; Sancho, D.; Slack, E.C.; LeibundGut-Landmann, S.; Reis e Sousa, C. Myeloid C-type lectins in innate immunity. Nat. Immunol. 2006, 7, 1258–1265. [Google Scholar] [CrossRef] [PubMed]
- Szolnoky, G.; Bata-Csorgo, Z.; Kenderessy, A.S.; Kiss, M.; Pivarcsi, A.; Novak, Z.; Nagy Newman, K.; Michel, G.; Ruzicka, T.; Marodi, L.; et al. A mannose-binding receptor is expressed on human keratinocytes and mediates killing of Candida albicans. J. Investig. Dermatol. 2001, 117, 205–213. [Google Scholar] [CrossRef]
- Martinez-Pomares, L. The mannose receptor. J. Leukoc. Biol. 2012, 92, 1177–1186. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, P.H.; Doebber, T.W.; Mandell, B.F.; White, R.; DeSchryver, C.; Rodman, J.S.; Miller, M.J.; Stahl, P. Plasma clearance of glycoproteins with terminal mannose and N-acetylglucosamine by liver non-parenchymal cells. Studies with β-glucuronidase, N-acetyl-β-d-glucosaminidase, ribonuclease B and agalacto-orosomucoid. Biochem. J. 1978, 176, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Stahl, P.D.; Ezekowitz, R.A. The mannose receptor is a pattern recognition receptor involved in host defense. Curr. Opin. Immunol. 1998, 10, 50–55. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; Pietersz, G.A.; Loveland, B.E.; Sandrin, M.S.; McKenzie, I.F. Oxidative/reductive conjugation of mannan to antigen selects for T1 or T2 immune responses. Proc. Natl. Acad. Sci. USA 1995, 92, 10128–10132. [Google Scholar] [CrossRef] [PubMed]
- Vassilaros, S.; Tsibanis, A.; Tsikkinis, A.; Pietersz, G.A.; McKenzie, I.F.; Apostolopoulos, V. Up to 15-year clinical follow-up of a pilot Phase III immunotherapy study in stage II breast cancer patients using oxidized mannan-MUC1. Immunotherapy 2013, 5, 1177–1182. [Google Scholar] [CrossRef]
- He, L.Z.; Weidlick, J.; Sisson, C.; Marsh, H.C.; Keler, T. Toll-like receptor agonists shape the immune responses to a mannose receptor-targeted cancer vaccine. Cell. Mol. Immunol. 2015, 12, 719–728. [Google Scholar] [CrossRef]
- Shrimpton, R.E.; Butler, M.; Morel, A.S.; Eren, E.; Hue, S.S.; Ritter, M.A. CD205 (DEC-205): A recognition receptor for apoptotic and necrotic self. Mol. Immunol. 2009, 46, 1229–1239. [Google Scholar] [CrossRef]
- Kato, M.; McDonald, K.J.; Khan, S.; Ross, I.L.; Vuckovic, S.; Chen, K.; Munster, D.; MacDonald, K.P.; Hart, D.N. Expression of human DEC-205 (CD205) multilectin receptor on leukocytes. Int. Immunol. 2006, 18, 857–869. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Swiggard, W.J.; Heufler, C.; Peng, M.; Mirza, A.; Steinman, R.M.; Nussenzweig, M.C. The receptor DEC-205 expressed by dendritic cells and thymic epithelial cells is involved in antigen processing. Nature 1995, 375, 151–155. [Google Scholar] [CrossRef]
- Dudziak, D.; Kamphorst, A.O.; Heidkamp, G.F.; Buchholz, V.R.; Trumpfheller, C.; Yamazaki, S.; Cheong, C.; Liu, K.; Lee, H.W.; Park, C.G.; et al. Differential antigen processing by dendritic cell subsets in vivo. Science 2007, 315, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zaidi, N.; He, L.Z.; Zhang, L.; Kuroiwa, J.M.; Keler, T.; Steinman, R.M. Targeting of the non-mutated tumor antigen HER2/neu to mature dendritic cells induces an integrated immune response that protects against breast cancer in mice. Breast Cancer Res. 2012, 14, R39. [Google Scholar] [CrossRef] [PubMed]
- Idoyaga, J.; Lubkin, A.; Fiorese, C.; Lahoud, M.H.; Caminschi, I.; Huang, Y.; Rodriguez, A.; Clausen, B.E.; Park, C.G.; Trumpfheller, C.; et al. Comparable T helper 1 (Th1) and CD8 T-cell immunity by targeting HIV gag p24 to CD8 dendritic cells within antibodies to Langerin, DEC205, and Clec9A. Proc. Natl. Acad. Sci. USA 2011, 108, 2384–2389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charalambous, A.; Oks, M.; Nchinda, G.; Yamazaki, S.; Steinman, R.M. Dendritic cell targeting of survivin protein in a xenogeneic form elicits strong CD4+ T cell immunity to mouse survivin. J. Immunol. 2006, 177, 8410–8421. [Google Scholar] [CrossRef] [PubMed]
- Bonifaz, L.C.; Bonnyay, D.P.; Charalambous, A.; Darguste, D.I.; Fujii, S.; Soares, H.; Brimnes, M.K.; Moltedo, B.; Moran, T.M.; Steinman, R.M. In vivo targeting of antigens to maturing dendritic cells via the DEC-205 receptor improves T cell vaccination. J. Exp. Med. 2004, 199, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Hawiger, D.; Inaba, K.; Dorsett, Y.; Guo, M.; Mahnke, K.; Rivera, M.; Ravetch, J.V.; Steinman, R.M.; Nussenzweig, M.C. Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo. J. Exp. Med. 2001, 194, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Cheong, C.; Choi, J.H.; Vitale, L.; He, L.Z.; Trumpfheller, C.; Bozzacco, L.; Do, Y.; Nchinda, G.; Park, S.H.; Dandamudi, D.B.; et al. Improved cellular and humoral immune responses in vivo following targeting of HIV Gag to dendritic cells within human anti-human DEC205 monoclonal antibody. Blood 2010, 116, 3828–3838. [Google Scholar] [CrossRef] [PubMed]
- Lahoud, M.H.; Ahmet, F.; Zhang, J.G.; Meuter, S.; Policheni, A.N.; Kitsoulis, S.; Lee, C.N.; O’Keeffe, M.; Sullivan, L.C.; Brooks, A.G.; et al. DEC-205 is a cell surface receptor for CpG oligonucleotides. Proc. Natl. Acad. Sci. USA 2012, 109, 16270–16275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.G.; Rodriguez, A.; Ueta, H.; Lee, H.; Pack, M.; Matsuno, K.; Steinman, R.M. Generation of anti-human DEC205/CD205 monoclonal antibodies that recognize epitopes conserved in different mammals. J. Immunol. Methods 2012, 377, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.Y.; Zhou, G.; Li, Q.L.; Lu, J.S.; Shi, D.Y.; Pang, X.B.; Zhou, X.W.; Yu, Y.Z.; Huang, P.T. Enhanced effects of DNA vaccine against botulinum neurotoxin serotype A by targeting antigen to dendritic cells. Immunol. Lett. 2017, 190, 118–124. [Google Scholar] [CrossRef]
- Beckman, R.A.; Weiner, L.M.; Davis, H.M. Antibody constructs in cancer therapy: Protein engineering strategies to improve exposure in solid tumors. Cancer 2007, 109, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Birkholz, K.; Schwenkert, M.; Kellner, C.; Gross, S.; Fey, G.; Schuler-Thurner, B.; Schuler, G.; Schaft, N.; Dorrie, J. Targeting of DEC-205 on human dendritic cells results in efficient MHC class II-restricted antigen presentation. Blood 2010, 116, 2277–2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakhrif, Z.; Moreau, A.; Herault, B.; Di-Tommaso, A.; Juste, M.; Moire, N.; Dimier-Poisson, I.; Mevelec, M.N.; Aubrey, N. Targeted delivery of Toxoplasma gondii antigens to dendritic cells promote immunogenicity and protective efficiency against Toxoplasmosis. Front. Immunol. 2018, 9, 317. [Google Scholar] [CrossRef] [PubMed]
- Ngu, L.N.; Nji, N.N.; Ambada, G.E.; Sagnia, B.; Sake, C.N.; Tchadji, J.C.; Njambe Priso, G.D.; Lissom, A.; Tchouangueu, T.F.; Manga Tebit, D.; et al. In vivo targeting of protein antigens to dendritic cells using anti-DEC-205 single chain antibody improves HIV Gag specific CD4(+) T cell responses protecting from airway challenge with recombinant vaccinia-gag virus. Immun. Inflamm. Dis. 2017. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Kane, J.R.; Panek, W.K.; Young, J.S.; Rashidi, A.; Yu, D.; Kanojia, D.; Hasan, T.; Miska, J.; Gomez-Lim, M.A.; et al. A dendritic cell-targeted adenoviral vector facilitates adaptive immune response against human glioma antigen (CMV-IE) and prolongs survival in a human glioma tumor model. Neurotherapeutics 2018, 15, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Antonio-Herrera, L.; Badillo-Godinez, O.; Medina-Contreras, O.; Tepale-Segura, A.; Garcia-Lozano, A.; Gutierrez-Xicotencatl, L.; Soldevila, G.; Esquivel-Guadarrama, F.R.; Idoyaga, J.; Bonifaz, L.C. The nontoxic cholera B subunit Is a potent adjuvant for intradermal DC-targeted vaccination. Front. Immunol. 2018, 9, 2212. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.M.; Gow, N.A.; Brown, G.D. Pattern recognition: Recent insights from Dectin-1. Curr. Opin. Immunol. 2009, 21, 30–37. [Google Scholar] [CrossRef]
- Brown, G.D. Dectin-1: A signalling non-TLR pattern-recognition receptor. Nat. Rev. Immunol. 2006, 6, 33–43. [Google Scholar] [CrossRef]
- Brown, G.D.; Gordon, S. Immune recognition. A new receptor for β-glucans. Nature 2001, 413, 36–37. [Google Scholar] [CrossRef]
- Monteiro, J.T.; Lepenies, B. Myeloid C-type lectin receptors in viral recognition and antiviral immunity. Viruses 2017, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Dennehy, K.M.; Brown, G.D. The role of the β-glucan receptor Dectin-1 in control of fungal infection. J. Leukoc. Biol. 2007, 82, 253–258. [Google Scholar] [CrossRef]
- Rivera, A.; Hohl, T.M.; Collins, N.; Leiner, I.; Gallegos, A.; Saijo, S.; Coward, J.W.; Iwakura, Y.; Pamer, E.G. Dectin-1 diversifies Aspergillus fumigatus-specific T cell responses by inhibiting T helper type 1 CD4 T cell differentiation. J. Exp. Med. 2011, 208, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Santos, N.; Gaffen, S.L. Th17 cells in immunity to Candida albicans. Cell Host Microbe 2012, 11, 425–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leibundgut-Landmann, S.; Osorio, F.; Brown, G.D.; Reis e Sousa, C. Stimulation of dendritic cells via the dectin-1/Syk pathway allows priming of cytotoxic T-cell responses. Blood 2008, 112, 4971–4980. [Google Scholar] [CrossRef] [Green Version]
- Graham, L.M.; Brown, G.D. The Dectin-2 family of C-type lectins in immunity and homeostasis. Cytokine 2009, 48, 148–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, Y.; Inoue, A.; Hangai, S.; Saijo, S.; Negishi, H.; Nishio, J.; Yamasaki, S.; Iwakura, Y.; Yanai, H.; Taniguchi, T. The innate immune receptor Dectin-2 mediates the phagocytosis of cancer cells by Kupffer cells for the suppression of liver metastasis. Proc. Natl. Acad. Sci. USA 2016, 113, 14097–14102. [Google Scholar] [CrossRef] [Green Version]
- Chiffoleau, E. C-type lectin-like receptors as emerging orchestrators of sterile inflammation represent potential therapeutic targets. Front. Immunol. 2018, 9, 227. [Google Scholar] [CrossRef]
- Kerrigan, A.M.; Brown, G.D. C-type lectins and phagocytosis. Immunobiology 2009, 214, 562–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appelmelk, B.J.; van Die, I.; van Vliet, S.J.; Vandenbroucke-Grauls, C.M.; Geijtenbeek, T.B.; van Kooyk, Y. Cutting edge: Carbohydrate profiling identifies new pathogens that interact with dendritic cell-specific ICAM-3-grabbing nonintegrin on dendritic cells. J. Immunol. 2003, 170, 1635–1639. [Google Scholar] [CrossRef] [PubMed]
- Engering, A.; Geijtenbeek, T.B.; van Vliet, S.J.; Wijers, M.; van Liempt, E.; Demaurex, N.; Lanzavecchia, A.; Fransen, J.; Figdor, C.G.; Piguet, V.; et al. The dendritic cell-specific adhesion receptor DC-SIGN internalizes antigen for presentation to T cells. J. Immunol. 2002, 168, 2118–2126. [Google Scholar] [CrossRef]
- Geijtenbeek, T.B.; Kwon, D.S.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Middel, J.; Cornelissen, I.L.; Nottet, H.S.; KewalRamani, V.N.; Littman, D.R.; et al. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell 2000, 100, 587–597. [Google Scholar] [CrossRef]
- van Dinther, D.; Stolk, D.A.; van de Ven, R.; van Kooyk, Y.; de Gruijl, T.D.; den Haan, J.M.M. Targeting C-type lectin receptors: A high-carbohydrate diet for dendritic cells to improve cancer vaccines. J. Leukoc. Biol. 2017, 102, 1017–1034. [Google Scholar] [CrossRef]
- Cruz, L.J.; Tacken, P.J.; Fokkink, R.; Joosten, B.; Stuart, M.C.; Albericio, F.; Torensma, R.; Figdor, C.G. Targeted PLGA nano- but not microparticles specifically deliver antigen to human dendritic cells via DC-SIGN in vitro. J. Control. Release 2010, 144, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Hesse, C.; Ginter, W.; Forg, T.; Mayer, C.T.; Baru, A.M.; Arnold-Schrauf, C.; Unger, W.W.; Kalay, H.; van Kooyk, Y.; Berod, L.; et al. In vivo targeting of human DC-SIGN drastically enhances CD8(+) T-cell-mediated protective immunity. Eur. J. Immunol. 2013, 43, 2543–2553. [Google Scholar] [CrossRef]
- Kretz-Rommel, A.; Qin, F.; Dakappagari, N.; Torensma, R.; Faas, S.; Wu, D.; Bowdish, K.S. In vivo targeting of antigens to human dendritic cells through DC-SIGN elicits stimulatory immune responses and inhibits tumor growth in grafted mouse models. J. Immunother. 2007, 30, 715–726. [Google Scholar] [CrossRef]
- Tacken, P.J.; Joosten, B.; Reddy, A.; Wu, D.; Eek, A.; Laverman, P.; Kretz-Rommel, A.; Adema, G.J.; Torensma, R.; Figdor, C.G. No advantage of cell-penetrating peptides over receptor-specific antibodies in targeting antigen to human dendritic cells for cross-presentation. J. Immunol. 2008, 180, 7687–7696. [Google Scholar] [CrossRef]
- Cruz, F.M.; Colbert, J.D.; Merino, E.; Kriegsman, B.A.; Rock, K.L. The biology and underlying mechanisms of cross-presentation of exogenous antigens on MHC-I molecules. Annu. Rev. Immunol. 2017, 35, 149–176. [Google Scholar] [CrossRef]
- Ahrens, S.; Zelenay, S.; Sancho, D.; Hanc, P.; Kjaer, S.; Feest, C.; Fletcher, G.; Durkin, C.; Postigo, A.; Skehel, M.; et al. F-actin is an evolutionarily conserved damage-associated molecular pattern recognized by DNGR-1, a receptor for dead cells. Immunity 2012, 36, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Sancho, D.; Mourao-Sa, D.; Joffre, O.P.; Schulz, O.; Rogers, N.C.; Pennington, D.J.; Carlyle, J.R.; Reis e Sousa, C. Tumor therapy in mice via antigen targeting to a novel, DC-restricted C-type lectin. J. Clin. Investig. 2008, 118, 2098–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picco, G.; Beatson, R.; Taylor-Papadimitriou, J.; Burchell, J.M. Targeting DNGR-1 (CLEC9A) with antibody/MUC1 peptide conjugates as a vaccine for carcinomas. Eur. J. Immunol. 2014, 44, 1947–1955. [Google Scholar] [CrossRef] [Green Version]
- Joffre, O.P.; Sancho, D.; Zelenay, S.; Keller, A.M.; Reis e Sousa, C. Efficient and versatile manipulation of the peripheral CD4+ T-cell compartment by antigen targeting to DNGR-1/CLEC9A. Eur. J. Immunol. 2010, 40, 1255–1265. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ahmet, F.; Sullivan, L.C.; Brooks, A.G.; Kent, S.J.; De Rose, R.; Salazar, A.M.; Reis e Sousa, C.; Shortman, K.; Lahoud, M.H.; et al. Antibodies targeting Clec9A promote strong humoral immunity without adjuvant in mice and non-human primates. Eur. J. Immunol. 2015, 45, 854–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, O.; Hanc, P.; Bottcher, J.P.; Hoogeboom, R.; Diebold, S.S.; Tolar, P.; Reis, E.S.C. Myosin II synergizes with F-Actin to promote DNGR-1-dependent cross-presentation of dead cell-associated antigens. Cell Rep. 2018, 24, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, F.; Ravetch, J.V. Fcgamma receptors as regulators of immune responses. Nat. Rev. Immunol. 2008, 8, 34–47. [Google Scholar] [CrossRef]
- Sanchez-Mejorada, G.; Rosales, C. Signal transduction by immunoglobulin Fc receptors. J. Leukoc. Biol. 1998, 63, 521–533. [Google Scholar] [CrossRef] [Green Version]
- Pincetic, A.; Bournazos, S.; DiLillo, D.J.; Maamary, J.; Wang, T.T.; Dahan, R.; Fiebiger, B.M.; Ravetch, J.V. Type I and type II Fc receptors regulate innate and adaptive immunity. Nat. Immunol. 2014, 15, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Bournazos, S.; Ravetch, J.V. Fcgamma receptor function and the design of vaccination strategies. Immunity 2017, 47, 224–233. [Google Scholar] [CrossRef]
- Swanson, J.A.; Hoppe, A.D. The coordination of signaling during Fc receptor-mediated phagocytosis. J. Leukoc. Biol. 2004, 76, 1093–1103. [Google Scholar] [CrossRef] [Green Version]
- Roopenian, D.C.; Akilesh, S. FcRn: The neonatal Fc receptor comes of age. Nat. Rev. Immunol. 2007, 7, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Mallery, D.L.; McEwan, W.A.; Bidgood, S.R.; Towers, G.J.; Johnson, C.M.; James, L.C. Antibodies mediate intracellular immunity through tripartite motif-containing 21 (TRIM21). Proc. Natl. Acad. Sci. USA 2010, 107, 19985–19990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilliams, M.; Bruhns, P.; Saeys, Y.; Hammad, H.; Lambrecht, B.N. The function of Fcgamma receptors in dendritic cells and macrophages. Nat. Rev. Immunol. 2014, 14, 94–108. [Google Scholar] [CrossRef]
- Pham, G.H.; Iglesias, B.V.; Gosselin, E.J. Fc receptor-targeting of immunogen as a strategy for enhanced antigen loading, vaccination, and protection using intranasally administered antigen-pulsed dendritic cells. Vaccine 2014, 32, 5212–5220. [Google Scholar] [CrossRef]
- Bitsaktsis, C.; Babadjanova, Z.; Gosselin, E.J. In vivo mechanisms involved in enhanced protection utilizing an Fc receptor-targeted mucosal vaccine platform in a bacterial vaccine and challenge model. Infect. Immun. 2015, 83, 77–89. [Google Scholar] [CrossRef]
- de Jong, J.M.; Schuurhuis, D.H.; Ioan-Facsinay, A.; van der Voort, E.I.; Huizinga, T.W.; Ossendorp, F.; Toes, R.E.; Verbeek, J.S. Murine Fc receptors for IgG are redundant in facilitating presentation of immune complex derived antigen to CD8+ T cells in vivo. Mol. Immunol. 2006, 43, 2045–2050. [Google Scholar] [CrossRef]
- Hossain, M.K.; Wall, K.A. Immunological evaluation of recent MUC1 glycopeptide cancer vaccines. Vaccines 2016, 4, 25. [Google Scholar] [CrossRef] [PubMed]
- Lakshminarayanan, V.; Supekar, N.T.; Wei, J.; McCurry, D.B.; Dueck, A.C.; Kosiorek, H.E.; Trivedi, P.P.; Bradley, J.M.; Madsen, C.S.; Pathangey, L.B.; et al. MUC1 vaccines, comprised of glycosylated or non-glycosylated peptides or tumor-derived MUC1, can circumvent immunoediting to control tumor growth in MUC1 transgenic mice. PLoS ONE 2016, 11, e0145920. [Google Scholar] [CrossRef]
- Gong, J.; Apostolopoulos, V.; Chen, D.; Chen, H.; Koido, S.; Gendler, S.J.; McKenzie, I.F.; Kufe, D. Selection and characterization of MUC1-specific CD8+ T cells from MUC1 transgenic mice immunized with dendritic-carcinoma fusion cells. Immunology 2000, 101, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Karmakar, P.; Lee, K.; Sarkar, S.; Wall, K.A.; Sucheck, S.J. Synthesis of a Liposomal MUC1 glycopeptide-based immunotherapeutic and evaluation of the effect of l-Rhamnose targeting on cellular immune responses. Bioconjug. Chem. 2016, 27, 110–120. [Google Scholar] [CrossRef]
- Hossain, M.K.; Vartak, A.; Karmakar, P.; Sucheck, S.J.; Wall, K.A. Augmenting vaccine immunogenicity through the use of natural human anti-rhamnose antibodies. ACS Chem. Biol. 2018, 13, 2130–2142. [Google Scholar] [CrossRef] [PubMed]
- Ngu, L.N.; Nji, N.N.; Ambada, G.; Ngoh, A.A.; Njambe Priso, G.D.; Tchadji, J.C.; Lissom, A.; Magagoum, S.H.; Sake, C.N.; Tchouangueu, T.F.; et al. Dendritic cell targeted HIV-1 gag protein vaccine provides help to a recombinant Newcastle disease virus vectored vaccine including mobilization of protective CD8(+) T cells. Immun. Inflamm. Dis. 2018, 6, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Jiao, Y.Y.; Ma, Y.; Peng, X.L.; Fu, Y.H.; Zhang, X.J.; Zheng, Y.B.; Zheng, Y.P.; Hong, T.; He, J.S. Enhanced humoral and CD8+ T cell immunity in mice vaccinated by DNA vaccine against human respiratory syncytial virus through targeting the encoded F protein to dendritic cells. Int. Immunopharmacol. 2017, 46, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Glaffig, M.; Stergiou, N.; Hartmann, S.; Schmitt, E.; Kunz, H. A synthetic MUC1 anticancer vaccine containing mannose ligands for targeting macrophages and dendritic cells. ChemMedChem 2018, 13, 25–29. [Google Scholar] [CrossRef]
- Silva, J.M.; Zupancic, E.; Vandermeulen, G.; Oliveira, V.G.; Salgado, A.; Videira, M.; Gaspar, M.; Graca, L.; Preat, V.; Florindo, H.F. In vivo delivery of peptides and Toll-like receptor ligands by mannose-functionalized polymeric nanoparticles induces prophylactic and therapeutic anti-tumor immune responses in a melanoma model. J. Control. Release 2015, 198, 91–103. [Google Scholar] [CrossRef]
- Dzharullaeva, A.S.; Tukhvatulin, A.I.; Erokhova, A.S.; Bandelyuk, A.S.; Polyakov, N.B.; Solovyev, A.I.; Nikitenko, N.A.; Shcheblyakov, D.V.; Naroditsky, B.S.; Logunov, D.Y.; et al. Stimulation of Dectin-1 and Dectin-2 during Parenteral Immunization, but Not Mincle, Induces Secretory IgA in Intestinal Mucosa. J. Immunol. Res. 2018, 2018, 3835720. [Google Scholar] [CrossRef]
- Donadei, A.; Gallorini, S.; Berti, F.; O’Hagan, D.T.; Adamo, R.; Baudner, B.C. Rational design of adjuvant for skin delivery: Conjugation of synthetic β-glucan dectin-1 agonist to protein antigen. Mol. Pharm. 2015, 12, 1662–1672. [Google Scholar] [CrossRef]
- Velasquez, L.N.; Stuve, P.; Gentilini, M.V.; Swallow, M.; Bartel, J.; Lycke, N.Y.; Barkan, D.; Martina, M.; Lujan, H.D.; Kalay, H.; et al. Targeting Mycobacterium tuberculosis antigens to dendritic cells via the DC-specific-ICAM3-Grabbing-nonintegrin receptor induces strong T-Helper 1 immune responses. Front. Immunol. 2018, 9, 471. [Google Scholar] [CrossRef] [PubMed]
- Le Moignic, A.; Malard, V.; Benvegnu, T.; Lemiegre, L.; Berchel, M.; Jaffres, P.A.; Baillou, C.; Delost, M.; Macedo, R.; Rochefort, J.; et al. Preclinical evaluation of mRNA trimannosylated lipopolyplexes as therapeutic cancer vaccines targeting dendritic cells. J. Control. Release 2018, 278, 110–121. [Google Scholar] [CrossRef]
- Caminschi, I.; Vremec, D.; Ahmet, F.; Lahoud, M.H.; Villadangos, J.A.; Murphy, K.M.; Heath, W.R.; Shortman, K. Antibody responses initiated by Clec9A-bearing dendritic cells in normal and Batf3(-/-) mice. Mol. Immunol. 2012, 50, 9–17. [Google Scholar] [CrossRef]
- Gall, V.A.; Philips, A.V.; Qiao, N.; Clise-Dwyer, K.; Perakis, A.A.; Zhang, M.; Clifton, G.T.; Sukhumalchandra, P.; Ma, Q.; Reddy, S.M.; et al. Trastuzumab increases HER2 uptake and cross-presentation by dendritic cells. Cancer Res. 2017, 77, 5374–5383. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Motal, U.M.; Wang, S.; Awad, A.; Lu, S.; Wigglesworth, K.; Galili, U. Increased immunogenicity of HIV-1 p24 and gp120 following immunization with gp120/p24 fusion protein vaccine expressing alpha-gal epitopes. Vaccine 2010, 28, 1758–1765. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Motal, U.M.; Guay, H.M.; Wigglesworth, K.; Welsh, R.M.; Galili, U. Immunogenicity of influenza virus vaccine is increased by anti-gal-mediated targeting to antigen-presenting cells. J. Virol. 2007, 81, 9131–9141. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receptors | Antigens | Adjuvant | Mice | Immunization | Results | Ref. |
|---|---|---|---|---|---|---|
| DEC205 | 1. SAG-1 (T. gondii) | 1. Poly (I:C) | 1. CBA/J (H-2k) | 1. IN & SubQ | 1. Th1 (IFN-γ, IL-2, IgG2a, IgA) | [62] |
| 2. HIV-1 gagP24 | 2. Poly-ICLC | 2. CXB6 F1 | 2. SubQ | 2. CD4+ T cells | [109] | |
| 3. Ova | 3. CTB | 3. C57BL/6 and Tg GFP | 3. ID | 3. CD4+ T cells, Th1, Th17 | [65] | |
| 4. AHc | 4. No adjuvant | 4. Balb/c | 4. IM | 4. IgG1, IgG2a, Mature DC | [59] | |
| 5. RSV fusion protein | 5. No adjuvant | 5. Balb/c | 5. IM | 5. Th1, CD8+ T cells | [110] | |
| MR | 1. MUC1 | 1. IFA | 1. Balb/c | 1. IP | 1. IgG1, IgG2a, IgG2b | [111] |
| 2. MAA | 2. CpG and Poly(I:C) | 2. C57Bl/6 and Tg OT-II | 2. SubQ | 2. IgG, IgG1, IgG2c, Th1 | [112] | |
| 3. hCGβ | 3. CpG and Poly-ICLC | 3. hMR-Tg and WT | 3. SubQ | 3. Th1 | [46] | |
| Dectin-1 | 1. Ova | 1. Curdlan | 1. C57BL/6 | 1. SubQ | 1. IgG1, IgG2a, IgG3, IgA, CD4+ and CD8+ T cells | [113] |
| 2. Diphtheria toxin (CRM197) | 2. β-glucans hexamer | 2. Balb/c | 2. ID | 2. IgG1, IgG2a | [114] | |
| DC-SIGN | 1. Ag85B (Mtb) | 1. CTA1-DD and zymosan | 1. hSIGN and P25ktk | 1. IP | 1. CD4+ T cells, IFN-γ | [115] |
| 2. triMN-LPR | 2. triMN-LPR as intrinsic adjuvant | 2. C57BL/6J (H-2b) CD11c-YFP | 2. ID | 2. DC upregulation, CD8+ T cells | [116] | |
| DNGR-1 | 1. Ova | 1. Poly (I:C) | 1. C57BL/6J WEHI, Clec9A−/−, IRF8−/−, Batf3−/− mice | 1. IV | 1. IgG, CD4+ and CD8+ T cells | [117] |
| 2. Anti-Clec9A | 2. With or without Poly (I:C) | 2. C57BL/6 | 2. IV | 2. Ig | [91] | |
| 3. Ova | 3. With or without anti-CD40 | 3. C57BL/6, B6.SJL and OT-I × rag−/− | 3. IV | 3. CD8+ T cells, IFN-γ | [88] | |
| 4. MUC1 | 4. Anti-CD40 and Poly (I:C) | 4. MUC1xA2K/b Tg | 4. SubQ | 4. CD8+ T cells, IFN-γ | [89] | |
| FcγR | 1. MUC1-Tn | 1. Pam3CysSK4 | 1. C57BL/6 | 1. IP | 1. IgG, IFN-γ, CD4+ and CD8+ T cells | [108] |
| 2. E75 (HER-2) | 2. GM-CSF | 2. FVB/N-Tg(MMTV-neu), Balb/c | 2. IP | 2. CTL | [118] | |
| 3. iFT | 3. No Adjuant | 3. C57BL/6, B6.129S1 Il12atm1Jm/J | 3. IN | 3. DC upregulation, IFN-γ secreting CD4+ T cells | [102] | |
| 4. gp120αgal/p24 | 4. Ribi adjuvant | 4. α1,3GT KO | 4. IP | 4. IgG, CD4+ and CD8+ T cells | [119] | |
| 5. α-gal | 5. Ribi adjuvant | 5. α1,3GT KO | 5. IP | 5. IgG, CD4+ and CD8+ T cells | [120] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.K.; Wall, K.A. Use of Dendritic Cell Receptors as Targets for Enhancing Anti-Cancer Immune Responses. Cancers 2019, 11, 418. https://doi.org/10.3390/cancers11030418
Hossain MK, Wall KA. Use of Dendritic Cell Receptors as Targets for Enhancing Anti-Cancer Immune Responses. Cancers. 2019; 11(3):418. https://doi.org/10.3390/cancers11030418
Chicago/Turabian StyleHossain, Md Kamal, and Katherine A. Wall. 2019. "Use of Dendritic Cell Receptors as Targets for Enhancing Anti-Cancer Immune Responses" Cancers 11, no. 3: 418. https://doi.org/10.3390/cancers11030418
APA StyleHossain, M. K., & Wall, K. A. (2019). Use of Dendritic Cell Receptors as Targets for Enhancing Anti-Cancer Immune Responses. Cancers, 11(3), 418. https://doi.org/10.3390/cancers11030418