Polymorphism in Toll-Like Receptors and Helicobacter Pylori Motility in Autoimmune Atrophic Gastritis and Gastric Cancer

 ,
,  , , , , , ,
, , , , , ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Design of the Study and Patient’s Characteristics
2.2. Individual TLR-Polymorphisms Showed an Association of TLR5 rs5744174 Genotype-C/T and TLR9 rs5743836 Allele-T in AAG Patients Compared to Healthy Donors
2.3. Targeted H. pylori Flagellin Protein Identification and Quantification by LC-MS/MS Spectrometry
2.4. DNA Sequencing of H. pylori Flagellin A
2.5. Motility Assays of H. pylori Isolates
2.6. Identification of the CagA Virulent Gene in H. pylori Isolates
2.7. Patients Having TLR5 rs5744174 T Allele Show a Tendency to Have a Higher H. pylori Antibody Titer and H. pylori Strains with a Lower Flagellin an Abundance
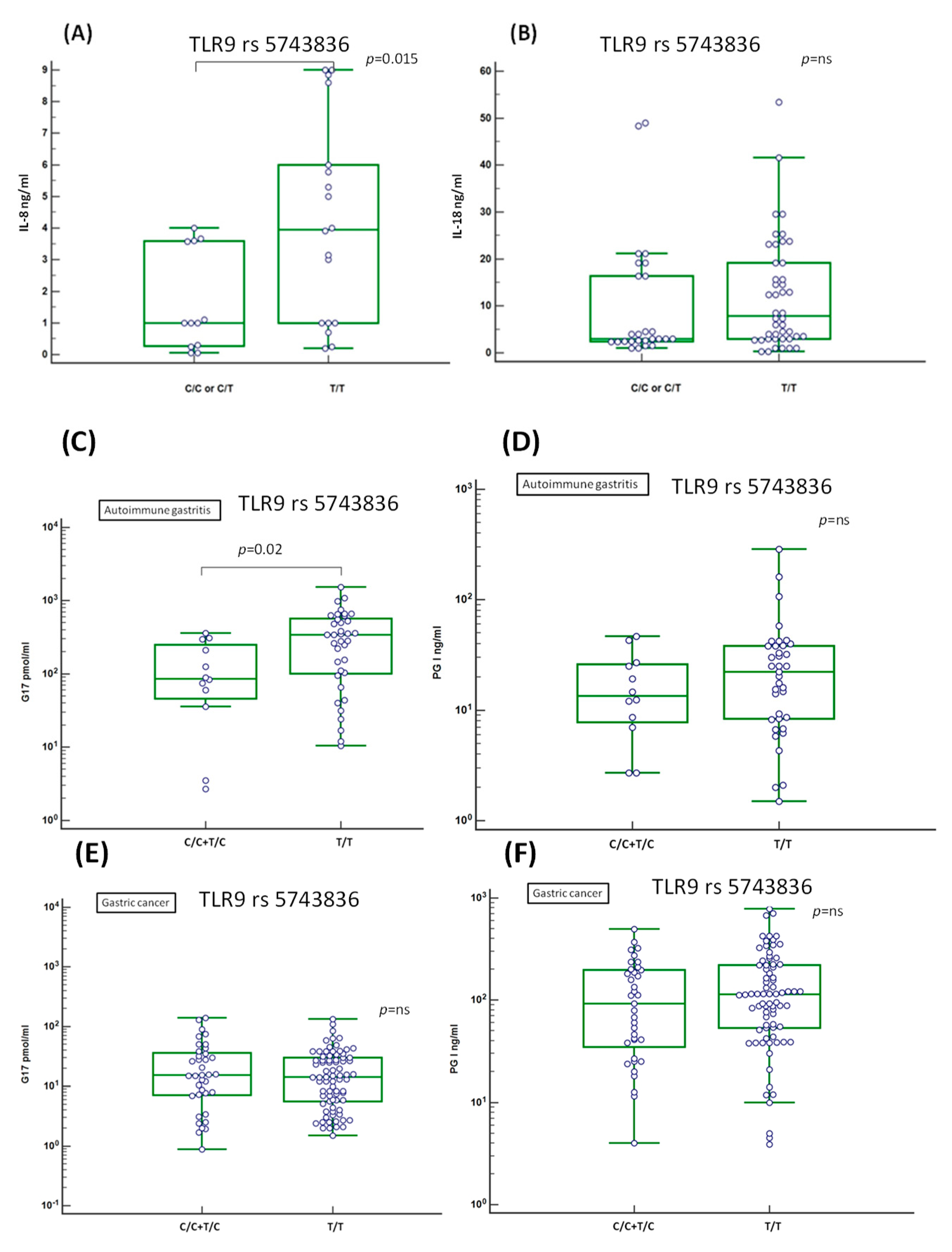
2.8. Evaluation of the Serum Level of Inflammation-Related IL-8 and IL-18 Cytokines
2.9. Association of the Serum Level of Atrophic Markers Gastrin G17 and PGI with TLR9 rs5743836 T Allele
3. Discussion
4. Materials and Methods
4.1. Patient Characteristics and Ethics Statement (IRB-14-2013)
4.2. Selection of TLRs Genetic Variants
4.3. Analysis of TLR Polymorphisms
4.4. Anti-Flagellin Antibody and Liquid MS/MS Spectrometry Validation
4.5. Flagellin A Nucleotide Sequencing
4.6. Motility Assays of H. pylori Isolates
4.7. Identification of the CagA Gene
4.8. IL-8 and IL-18 Cytokines Assay
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jacobson, D.L.; Gange, S.J.; Rose, N.R.; Graham, N.M. Epidemiology and estimated population burden of selected autoimmune diseases in the United States. Clin. Immunol. Immunopathol. 1997, 84, 223–243. [Google Scholar] [CrossRef]
- Di Sabatino, A.; Lenti, M.V.; Giuffrida, P.; Vanoli, A.; Corazza, G.R. New insights into immune mechanismsunderlying autoimmune diseases of the gastrointestinal tract. Autoimmun. Rev. 2015, 14, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, N.; Stashek, K.; Katona, B.W.; Tondon, R.; Shroff, S.G.; Roses, R.; Furth, E.E.; Metz, D.C. Theincidence of neoplasia in patients with autoimmune metaplastic atrophic gastritis: A renewed call forsurveillance. Ann. Gastroenterol. 2019, 32, 67–72. [Google Scholar] [PubMed]
- Neumann, W.L.; Coss, E.; Rugge, M.; Genta, R.M. Autoimmune atrophic gastritis—Pathogenesis, pathologyand management. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 529–541. [Google Scholar] [CrossRef]
- De Re, V.; Repetto, O.; Zanussi, S.; Casarotto, M.; Caggiari, L.; Canzonieri, V.; Cannizzaro, R. Protein signature characterizing Helicobacter pylori strains of patients with autoimmune atrophic gastritis, duodenal ulcer and gastric cancer. Infect. Agents Cancer 2017, 12, 22. [Google Scholar] [CrossRef]
- Repetto, O.; Zanussi, S.; Casarotto, M.; Canzonieri, V.; De Paoli, P.; Cannizzaro, R.; De Re, V. Differential 655 proteomics of Helicobacter pylori associated with autoimmune atrophic gastritis. Mol. Med. 2014, 20, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Rabkin, C.S.; Camargo, M.C. Gastric Cancer: An Evolving Disease. Curr. Treat. Options Gastroenterol. 2018, 16, 561–569. [Google Scholar] [CrossRef]
- Mayerle, J.; den Hoed, C.M.; Schurmann, C.; Stolk, L.; Homuth, G.; Peters, M.J.; Capelle, L.G.; Zimmermann, K.; Rivadeneira, F.; Gruska, S.; et al. Identification of genetic lociassociated with Helicobacter pylori serologic status. JAMA 2013, 309, 1912–1920. [Google Scholar] [CrossRef]
- Castano-Rodriguez, N.; Kaakoush, N.O.; Goh, K.L.; Fock, K.M.; Mitchell, H.M. The role of TLR2, TLR4 andCD14 genetic polymorphisms in gastric carcinogenesis: A case-control study and meta-analysis. PLoS ONE 2013, 8, e60327. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, J.; Xie, W.; Zhang, W.; Chang, Y. Toll-like receptor 2 gene polymorphisms and cancersusceptibility: A meta-analysis. Neoplasma 2013, 60, 459–467. [Google Scholar] [CrossRef]
- Achyut, B.R.; Ghoshal, U.C.; Moorchung, N.; Mittal, B. Association of Toll-like receptor-4 (Asp299Gly and Thr399Ileu) gene polymorphisms with gastritis and precancerous lesions. Hum. Immunol. 2007, 68, 901–907. [Google Scholar] [CrossRef]
- Zeng, H.M.; Pan, K.F.; Zhang, Y.; Zhang, L.; Ma, J.L.; Zhou, T.; Su, H.J.; Li, W.Q.; Li, J.Y.; Gerhard, M.; et al. Genetic variants of toll-like receptor 2 and 5, helicobacter pylori infection, and risk of gastric cancer and itsprecursors in a chinese population. Cancer Epidemiol. Biomark. Prev. 2011, 20, 2594–2602. [Google Scholar] [CrossRef]
- Gantier, M.P.; Irving, A.T.; Kaparakis-Liaskos, M.; Xu, D.; Evans, V.A.; Cameron, P.U.; Bourne, J.A.; Ferrero, R.L.; John, M.; Behlke, M.A.; et al. Genetic modulation of TLR8 response following bacterial phagocytosis. Hum. Mutat. 2010, 31, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.T.; Van’t Hof, R.; Crockett, J.C.; Hope, M.E.; Berry, S.; Thomson, J.; McLean, M.H.; McColl, K.E.; El-Omar, E.M.; Hold, G.L. Increase in NF-kappaB binding affinity of the variant C allele of the toll-like receptor 9 -1237T/C polymorphism is associated with Helicobacter pylori-induced gastric disease. Infect. Immun. 2010, 78, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Swartz, J.R. Functional properties of flagellin as a stimulator of innate immunity. Sci. Rep. 2016, 6, 18379. [Google Scholar] [CrossRef]
- Matusiak, M.; Van, O.N.; Vande, W.L.; Sirard, J.C.; Kanneganti, T.D.; Lamkanfi, M. Flagellin-inducedNLRC4 phosphorylation primes the inflammasome for activation by NAIP5. Proc. Natl. Acad. Sci. USA 2015, 112, 1541–1546. [Google Scholar] [CrossRef]
- Franco, A.T.; Friedman, D.B.; Nagy, T.A.; Romero-Gallo, J.; Krishna, U.; Kendall, A.; Israel, D.A.; Tegtmeyer, N.; Washington, M.K.; Peek, R.M., Jr. Delineation of a carcinogenic Helicobacter pylori proteome. Mol. Cell. Proteom. 2009, 8, 1947–1958. [Google Scholar] [CrossRef]
- Schirm, M.; Soo, E.C.; Aubry, A.J.; Austin, J.; Thibault, P.; Logan, S.M. Structural, genetic and functionalcharacterization of the flagellin glycosylation process in Helicobacter pylori. Mol. Microbiol. 2003, 48, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Song, W.S.; Jeon, Y.J.; Namgung, B.; Hong, M.; Yoon, S.I. A conserved TLR5 binding and activation hotspot on flagellin. Sci. Rep. 2017, 7, 40878. [Google Scholar] [CrossRef]
- De Re, V.; Orzes, E.; Canzonieri, V.; Maiero, S.; Fornasarig, M.; Alessandrini, L.; Cervo, S.; Steffan, A.; Zanette, G.; Mazzon, C.; et al. Pepsinogens to Distinguish Patients with Gastric Intestinal Metaplasia and Helicobacter pylori Infection Among Populations at Risk for Gastric Cancer. Clin. Transl. Gastroenterol. 2016, 7, e183. [Google Scholar] [CrossRef]
- Amedei, A.; Bergman, M.P.; Appelmelk, B.J.; Azzurri, A.; Benagiano, M.; Tamburini, C.; van der Zee, R.; Telford, J.L.; Vandenbroucke-Grauls, C.M.; D’Elios, M.M.; et al. Molecular mimicry between Helicobacter pylori antigens and H+, K+—adenosine triphosphatase in human gastric autoimmunity. J. Exp. Med. 2003, 198, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Toh, B.H.; Chan, J.; Kyaw, T.; Alderuccio, F. Cutting edge issues in autoimmune gastritis. Clin. Rev. Allergy Immunol. 2012, 42, 269–278. [Google Scholar] [CrossRef]
- Elshabrawy, H.A.; Essani, A.E.; Szekanecz, Z.; Fox, D.A.; Shahrara, S. TLRs, future potential therapeutictargets for RA. Autoimmun. Rev. 2017, 16, 103–113. [Google Scholar] [CrossRef]
- Wu, Y.W.; Tang, W.; Zuo, J.P. Toll-like receptors: Potential targets for lupus treatment. Acta Pharmacol. Sin. 2015, 36, 1395–1407. [Google Scholar] [CrossRef] [PubMed]
- Song, E.J.; Kang, M.J.; Kim, Y.S.; Kim, S.M.; Lee, S.E.; Kim, C.H.; Kim, D.J.; Park, J.H. Flagellin promotes the proliferation of gastric cancer cells via the Toll-like receptor 5. Int. J. Mol. Med. 2011, 28, 115–119. [Google Scholar]
- Xu, T.; Fu, D.; Ren, Y.; Dai, Y.; Lin, J.; Tang, L.; Ji, J. Genetic variations of TLR5 gene interacted with Helicobacter pylori infection among carcinogenesis of gastric cancer. Oncotarget 2017, 8, 31016–31022. [Google Scholar] [CrossRef] [PubMed]
- Engler, D.B.; Leonardi, I.; Hartung, M.L.; Kyburz, A.; Spath, S.; Becher, B.; Rogler, G.; Muller, A. Helicobacter pylori-specific protection against inflammatory bowel disease requires the NLRP3 inflammasomeand IL-18. Inflamm. Bowel. Dis. 2015, 21, 854–861. [Google Scholar] [CrossRef] [PubMed]
- Tenthorey, J.L.; Kofoed, E.M.; Daugherty, M.D.; Malik, H.S.; Vance, R.E. Molecular basis for specific recognition of bacterial ligands by NAIP/NLRC4 inflammasomes. Mol. Cell. 2014, 54, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Vijay-Kumar, M.; Carvalho, F.A.; Aitken, J.D.; Fifadara, N.H.; Gewirtz, A.T. TLR5 or NLRC4 is necessary and sufficient for promotion of humoral immunity by flagellin. Eur. J. Immunol. 2010, 40, 3528–3534. [Google Scholar] [CrossRef]
- Letran, S.E.; Lee, S.J.; Atif, S.M.; Flores-Langarica, A.; Uematsu, S.; Akira, S.; Cunningham, A.F.; McSorley, S.J. TLR5-deficient mice lack basal inflammatory and metabolic defects but exhibit impaired CD4 T cell responses to a flagellated pathogen. J. Immunol. 2011, 186, 5406–5412. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, X.; Zhang, B.; Shi, H.; Yuan, X.; Sun, Y.; Pan, Z.; Qian, H.; Xu, W. Exosomes derived from gastric cancer cells activate NF-kappaB pathway in macrophages to promote cancer progression. Tumour. Biol. 2016, 37, 12169–12180. [Google Scholar] [CrossRef] [PubMed]
- Schmausser, B.; Andrulis, M.; Endrich, S.; Muller-Hermelink, H.K.; Eck, M. Toll-like receptors TLR4, TLR5 and TLR9 on gastric carcinoma cells: An implication for interaction with Helicobacter pylori. Int. J. Med. Microbiol. 2005, 295, 179–185. [Google Scholar] [CrossRef]
- Asakura, H.; Churin, Y.; Bauer, B.; Boettcher, J.P.; Bartfeld, S.; Hashii, N.; Kawasaki, N.; Mollenkopf, H.J.; Jungblut, P.R.; Brinkmann, V.; et al. Helicobacter pylori HP0518 affects flagellin glycosylation to alter bacterial motility. Mol. Microbiol. 2010, 78, 1130–1144. [Google Scholar] [CrossRef] [PubMed]
- Gu, H. Role of Flagella in the Pathogenesis of Helicobacter pylori. Curr. Microbiol. 2017, 74, 863–869. [Google Scholar] [CrossRef]
- Kao, C.Y.; Sheu, B.S.; Sheu, S.M.; Yang, H.B.; Chang, W.L.; Cheng, H.C.; Wu, J.J. Higher motility enhancesbacterial density and inflammatory response in dyspeptic patients infected with Helicobacter pylori. Helicobacter 2012, 17, 411–416. [Google Scholar] [CrossRef]
- Kao, C.Y.; Sheu, B.S.; Wu, J.J. Helicobacter pylori infection: An overview of bacterial virulence factors and pathogenesis. Biomed. J. 2016, 39, 14–23. [Google Scholar] [CrossRef]
- Jeong, S.; Choi, E.; Petersen, C.P.; Roland, J.T.; Federico, A.; Ippolito, R.; D’Armiento, F.P.; Nardone, G.; Nagano, O.; Saya, H.; et al. Distinct metaplastic and inflammatory phenotypes in autoimmune andadenocarcinoma-associated chronic atrophic gastritis. United European. Gastroenterol. J. 2017, 5, 37–44. [Google Scholar]
- Sanders, C.J.; Yu, Y.; Moore, D.A., III; Williams, I.R.; Gewirtz, A.T. Humoral immune response to flagellin requires T cells and activation of innate immunity. J. Immunol. 2006, 177, 2810–2818. [Google Scholar] [CrossRef]
- Cullender, T.C.; Chassaing, B.; Janzon, A.; Kumar, K.; Muller, C.E.; Werner, J.J.; Angenent, L.T.; Bell, M.E.; Hay, A.G.; Peterson, D.A.; et al. Innate and adaptive immunity interact to quench microbiome flagellar motility in the gut. Cell Host Microbe 2013, 14, 571–581. [Google Scholar] [CrossRef]
- Klimosch, S.N.; Forsti, A.; Eckert, J.; Knezevic, J.; Bevier, M.; von Schönfels, W.; Heits, N.; Walter, J.; Hinz, S.; Lascorz, J.; et al. Functional TLR5 genetic variants affect human colorectal cancer survival. Cancer Res. 2013, 73, 7232–7242. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, N.; Ovsyannikova, I.G.; Vierkant, R.A.; Ryan, J.E.; Pankratz, V.S.; Jacobson, R.M.; Poland, G.A. Associations between SNPs in toll-like receptors and related intracellular signaling molecules and immune 681 responses to measles vaccine: Preliminary results. Vaccine 2008, 26, 1731–1736. [Google Scholar] [CrossRef]
- Grossman, S.R.; Andersen, K.G.; Shlyakhter, I.; Tabrizi, S.; Winnicki, S.; Yen, A.; Park, D.J.; Griesemer, D.; Karlsson, E.K.; Wong, S.H.; et al. Identifying recent adaptations in large-scale genomic data. Cell 2013, 152, 703–713. [Google Scholar] [CrossRef]
- Sun, S.C.; Chang, J.H.; Jin, J. Regulation of nuclear factor-kappaB in autoimmunity. Trends Immunol. 2013, 34, 282–289. [Google Scholar] [CrossRef]
- Lerner, A.; Jeremias, P.; Matthias, T. The World Incidence and Prevalence of Autoimmune Diseases is Increasing. Int. J. Celiac Disease 2015, 3, 151–155. [Google Scholar] [CrossRef]
- Ngo, S.T.; Steyn, F.J.; McCombe, P.A. Gender differences in autoimmune disease. Front. Neuroendocrinol. 2014, 35, 347–369. [Google Scholar] [CrossRef]
- Webb, K.; Butler, G.; Ciurtin, C.; Peckham, H.; Radziszewska, A.; Wedderburn, L.R.; Ioannou, Y. Sex differences in autoimmune disease: Testosterone is associated with a decrease in expression of key anti-viral genes during puberty, which may decrease the risk of autoimmunity in males. ESPE Abstracts 2018, 89, FC14.6. [Google Scholar]
- Berger, R.; Fiegl, H.; Goebel, G.; Obexer, P.; Ausserlechner, M.; Doppler, W.; Hauser-Kronberger, C.; Reitsamer, R.; Egle, D.; Reimer, D.; et al. Toll-like receptor 9 expression in breast and ovarian cancer is associated 722 with poorly differentiated tumors. Cancer Sci. 2010, 101, 1059–1066. [Google Scholar] [CrossRef] [PubMed]
- Krieg, A.M. Therapeutic potential of Toll-like receptor 9 activation. Nat. Rev. Drug Discov. 2006, 5, 471–484. [Google Scholar] [CrossRef]
- Rad, R.; Ballhorn, W.; Voland, P.; Eisenacher, K.; Mages, J.; Rad, L.; Ferstl, R.; Lang, R.; Wagner, H.; Schmid, R.M.; et al. Extracellular and intracellular pattern recognition receptors cooperate in the recognition of Helicobacter pylori. Gastroenterology 2009, 136, 2247–2257. [Google Scholar] [CrossRef]
- Otani, K.; Tanigawa, T.; Watanabe, T.; Nadatani, Y.; Sogawa, M.; Yamagami, H.; Shiba, M.; Watanabe, K.; Tominaga, K.; Fujiwara, Y.; et al. Toll-like receptor 9 signaling has anti-inflammatory effects on the early phase of Helicobacter pylori-induced gastritis. Biochem. Biophys. Res. Commun. 2012, 426, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Varga, M.G.; Peek, R.M. DNA Transfer and Toll-like Receptor Modulation by Helicobacter pylori. Curr. Top. Microbiol. Immunol. 2017, 400, 169–193. [Google Scholar]
- Varga, M.G.; Shaffer, C.L.; Sierra, J.C.; Suarez, G.; Piazuelo, M.B.; Whitaker, M.E.; Romero-Gallo, J.; Krishna, U.S.; Delgado, A.; Gomez, M.A.; et al. Pathogenic Helicobacter pylori strains translocate DNA and activate TLR9 via the cancer-associated cag type IV secretion system. Oncogene 2016, 35, 6262–6269. [Google Scholar] [CrossRef]
- Nagase, H.; Takeoka, T.; Urakawa, S.; Morimoto-Okazawa, A.; Kawashima, A.; Iwahori, K.; Takiguchi, S.; Nishikawa, H.; Sato, E.; Sakaguchi, S.; et al. ICOS(+) Foxp3(+) TILs in gastric cancer are prognostic markers and effector regulatory T cells associated with Helicobacter pylori. Int. J. Cancer 2017, 140, 686–695. [Google Scholar] [CrossRef]
- Lauren, P. The two histological main type of gastric carcinoma: Diffuse and so-called intestinal-typecarcinoma. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar] [CrossRef]
- Sobin, L.H.; Gospodarowicz, M.; Wittekind, C. TNM Classification of Malignant Tumours, 7th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2009. [Google Scholar]
- Di Sabatino, A.; Biagi, F.; Lenzi, M.; Frulloni, L.; Lenti, M.V.; Giuffrida, P.; Corazza, G.R. Clinicalusefulness of serum antibodies as biomarkers of gastrointestinal and liver diseases. Dig. Liver Dis. 2017, 49, 947–956. [Google Scholar] [CrossRef]
- Lenti, M.V.; Miceli, E.; Padula, D.; Di Sabatino, A.; Corazza, G.R. The challenging diagnosis ofautoimmune atrophic gastritis. Scand. J. Gastroenterol. 2017, 52, 471–472. [Google Scholar]
- Forstneric, V.; Ivicak-Kocjan, K.; Plaper, T.; Jerala, R.; Bencina, M. The role of the C-terminal D0 domainof flagellin in activation of Toll like receptor 5. PLoS Pathog. 2017, 13, e1006574. [Google Scholar] [CrossRef]
- Chi, H.; Flavell, R.A. Innate recognition of non-self nucleic acids. Genome Biol. 2008, 9, 211–219. [Google Scholar] [CrossRef]
- De Re, V.; De Zorzi, M.; Caggiari, L.; Lauletta, G.; Tornesello, M.L.; Fognani, E.; Miorin, M.; Racanelli, V.; Quartuccio, L.; Gragnani, L.; et al. HCV-related liver and lymphoproliferative diseases: Association withpolymorphisms of IL28B and TLR2. Oncotarget 2016, 7, 37487–37497. [Google Scholar] [CrossRef]
- Tomasini, M.L.; Zanussi, S.; Sozzi, M.; Tedeschi, R.; Basaglia, G.; De, P.P. Heterogeneity of cag genotypesin Helicobacter pylori isolates from human biopsy specimens. J. Clin. Microbiol. 2003, 41, 976–980. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic features | Age at Inclusion | Mean ± SEM (Range) | p | |
| GC | 61.45 ± 1.04 (19–85) | GC vs. HD, p < 0.001 | ||
| AAG | 54.59 ± 1.79 (31–70) | GC vs. AAG, p = 0.019 | ||
| HD | 42.03 ± 1.66 (24–64) | AAG vs. HD, p < 0.001 | ||
| Gender | Male | |||
| GC | 71 (62.3%) | GC vs. HD | ||
| AAG | 15 (22.4%) | GC vs. AAG, p < 0.001 | ||
| HD | 55 (56.7%) | AAG vs. HD, p < 0.001 | ||
| H. pylori status | IgG Anti-H. Pylori Antibodies | positive | ||
| GC | 34 (56.7%) | GC vs. HD, p < 0.001 | ||
| AAG | 16 (40.0%) | GC vs. AAG, p = 0.021 | ||
| HD | 18 (18.6%) | AAG vs. HD | ||
| Missed data | 101 | |||
| Pepsinogen | Pepsinogen I (ng/mL) | Mean ± SD | ||
| GC | 153.85 ± 137 | GC vs. HD, p < 0.001 | ||
| AAG | 37.29 ± 75 | GC vs. AAG, p = 0.001 | ||
| HD | 73.7 ± 89 | AAG vs. HD | ||
| Missed data | 50 | |||
| Pepsinogen II (ng/mL) | Mean ± SD | |||
| GC | 22.58 ± 19 | GC vs. HD, p < 0.001 | ||
| AAG | 12.13 ± 6 | GC vs. AAG, p = 0.007 | ||
| HD | 7.2 ± 5 | AAG vs. HD, p < 0.001 | ||
| Missed data | 50 | |||
| PG I/PG II Ratio | Mean ± SD | |||
| GC | 6.96 ± 4.17 | GC vs. HD | ||
| AAG | 2.43 ± 3.95 | GC vs. AAG | ||
| HD | 9.57 ± 2.78 | AAG vs. HD, p < 0.001 | ||
| Gastrin | Gastrin G17 (pg/mL) | Mean ± SD | ||
| GC | 20.47 ± 21 | GC vs. HD, p = 0.003 | ||
| AAG | 431.8 ± 504 | GC vs. AAG, p < 0.001 | ||
| HD | 7.7 ± 29 | AAG vs. HD, p < 0.001 | ||
| Missed data | 50 | |||
| Tumor characteristics (n = 60 GC) | GC Lauren Classification | |||
| Intestinal | 30 (50.0%) | |||
| Diffuse | 17 (28.3%) | |||
| Mixed | 10 (16.7%) | |||
| unspecified | 3 (5.0%) | |||
| GC Anatomical Site | ||||
| Proximal | 21 (35.0%) | |||
| Distal | 38 (63.3%) | |||
| unspecified | 1 (1.7%) | |||
| Clinical Stage | ||||
| T | T1–T2 (26.7%) | T3–T4 (73.3%) | ||
| N | no (30.0%) | yes (70.0%) | ||
| M | no (82.5%) | yes (17.5%) | ||
| Surgery | ||||
| Total gastrectomy | yes (40.7%) | |||
| Partial gastrectomy | yes (44.4%) | |||
| No surgery | yes (18.8%) | |||
| Autoimmune characteristics (n = 47 AAG) | Pernicious Anemia | yes (29.4%) | ||
| Familiarity for GC | yes (12.7%) | |||
| Gene | Chromosomal Position | Locus | SNP ID | Change Nucleotide AA | Reference | |
|---|---|---|---|---|---|---|
| TLR1 | 4p14 | exon 4 | rs4833095 | T > C | N248S | [8] |
| TLR2 | 4q32 | 5’UTR | -196 to -174del (rs111200466) | 22-bp ins/del | --- | [9] |
| exon 3 | rs3804099 | T > C | N199N | [10] | ||
| TLR4 | 9q33.1 | exon 3 | rs4986790 | A > G | D299G | [9,11] |
| exon 3 | rs4986791 | C > T | T399I | [9,11] | ||
| TLR5 | 1q41 | Exon 4 | rs5744174 | T > C | F616L | [12] |
| TLR8 | Xp22.2 | 5´UTR | rs3764880 | A > G | --- | [13] |
| TLR9 | 3p21.3 | 5´UTR | rs5743836 | T > C | --- | [14] |
| Primers ID | Sequence (bp) | Size Products | References |
|---|---|---|---|
| TLR2-196, -174 | FOR: CTCGGAGGCAGCGAGAAA | 286/264 | [60] |
| REV: CTGGGCCGTGCAAAGAAG | |||
| TLR4-EXON-3 | FOR: TCTGCTCTAGAGGGCCTGTGC | 632 | (designed by the authors) |
| REV: TCCTGGAAAGAATTGCCAGCCA | |||
| TLR5-5744174 | FOR: TGTCACTATAGCTGGGCCTC | 104 | (designed by the authors) |
| REV: CCTCTTCATCACAACCTTCCG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Re, V.; Repetto, O.; De Zorzi, M.; Casarotto, M.; Tedeschi, M.; Giuffrida, P.; Lenti, M.V.; Magris, R.; Miolo, G.; Mazzon, C.; et al. Polymorphism in Toll-Like Receptors and Helicobacter Pylori Motility in Autoimmune Atrophic Gastritis and Gastric Cancer. Cancers 2019, 11, 648. https://doi.org/10.3390/cancers11050648
De Re V, Repetto O, De Zorzi M, Casarotto M, Tedeschi M, Giuffrida P, Lenti MV, Magris R, Miolo G, Mazzon C, et al. Polymorphism in Toll-Like Receptors and Helicobacter Pylori Motility in Autoimmune Atrophic Gastritis and Gastric Cancer. Cancers. 2019; 11(5):648. https://doi.org/10.3390/cancers11050648
Chicago/Turabian StyleDe Re, Valli, Ombretta Repetto, Mariangela De Zorzi, Mariateresa Casarotto, Massimo Tedeschi, Paolo Giuffrida, Marco Vincenzo Lenti, Raffaella Magris, Gianmaria Miolo, Cinzia Mazzon, and et al. 2019. "Polymorphism in Toll-Like Receptors and Helicobacter Pylori Motility in Autoimmune Atrophic Gastritis and Gastric Cancer" Cancers 11, no. 5: 648. https://doi.org/10.3390/cancers11050648
APA StyleDe Re, V., Repetto, O., De Zorzi, M., Casarotto, M., Tedeschi, M., Giuffrida, P., Lenti, M. V., Magris, R., Miolo, G., Mazzon, C., Zanette, G., Alessandrini, L., Canzonieri, V., Caggiari, L., Zanussi, S., Steffan, A., Di Sabatino, A., & Cannizzaro, R. (2019). Polymorphism in Toll-Like Receptors and Helicobacter Pylori Motility in Autoimmune Atrophic Gastritis and Gastric Cancer. Cancers, 11(5), 648. https://doi.org/10.3390/cancers11050648