APOBEC3B Gene Expression in Ductal Carcinoma In Situ and Synchronous Invasive Breast Cancer

 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. General Clinicopathological Data
2.2. APOBEC3B Expression in Synchronous Normal, DCIS and IBC Cells
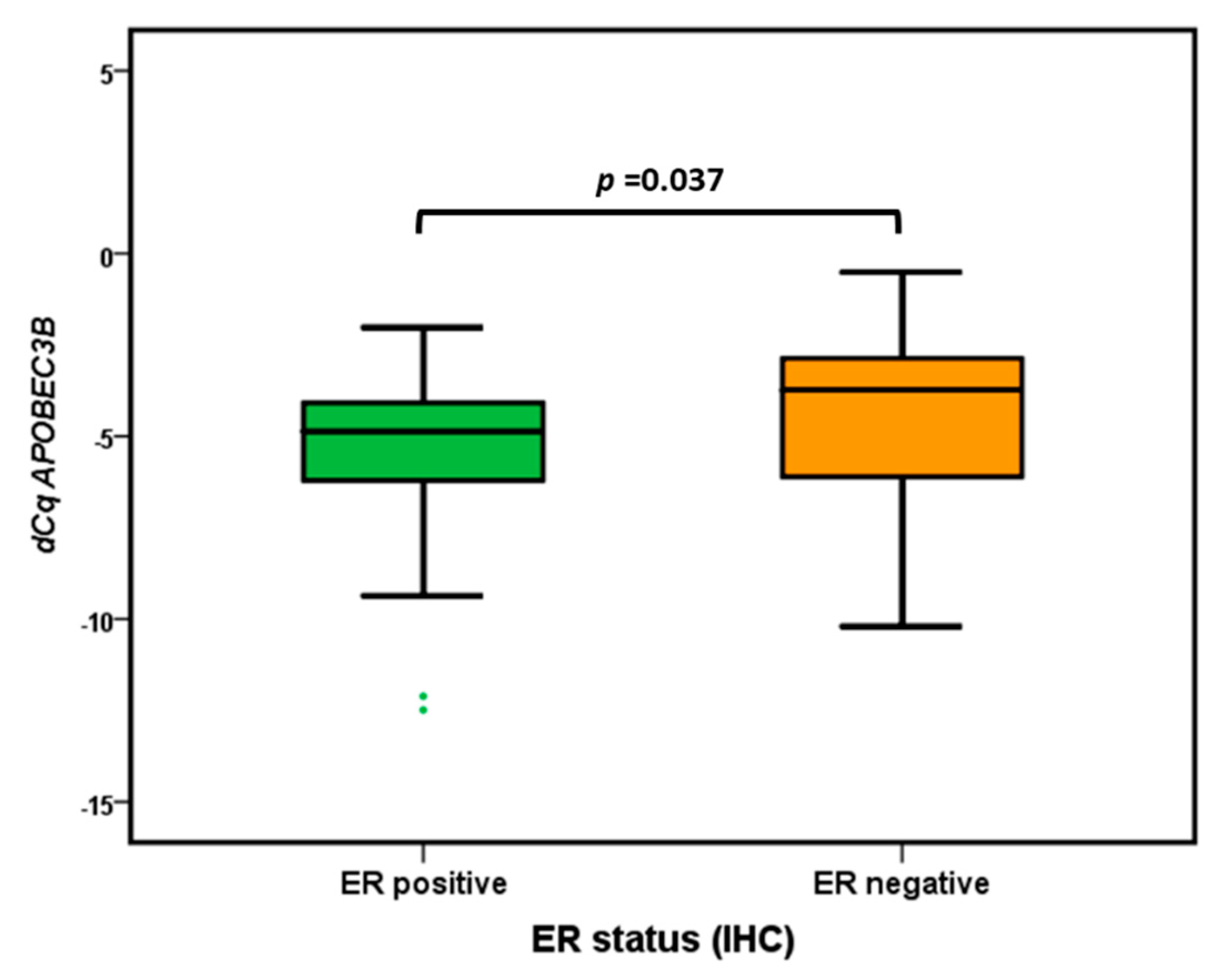
2.3. APOBEC3B mRNA Subgroup Analysis
2.4. APOBEC3B Expression in Epithelial Versus Inflammatory Cells
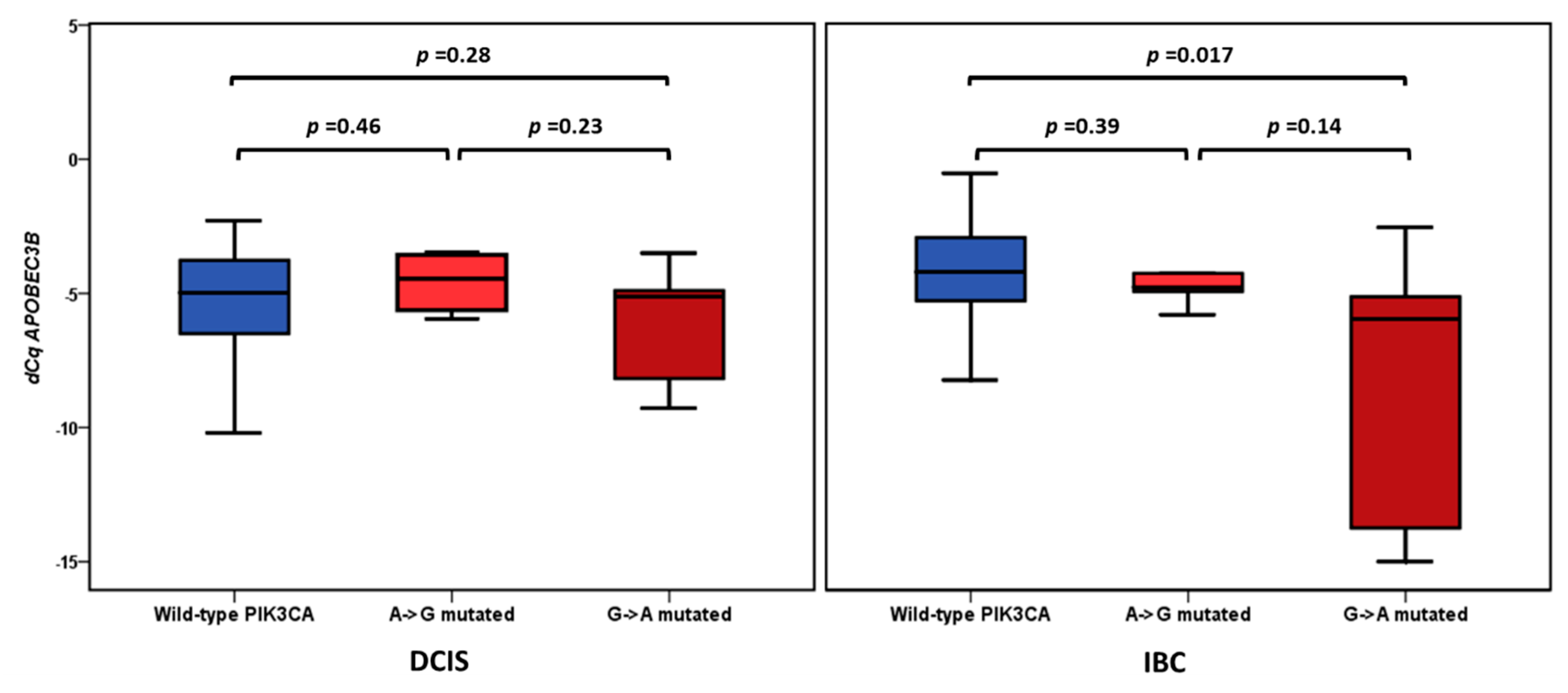
2.5. APOBEC3B Expression and PIK3CA Mutation Status
3. Discussion
4. Materials and Methods
4.1. Patient Materials
4.2. RT-qPCR
4.3. PIK3CA Mutation Status
4.4. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cowell, C.F.; Weigelt, B.; Sakr, R.A.; Ng, C.K.; Hicks, J.; King, T.A.; Reis-Filho, J.S. Progression from ductal carcinoma in situ to invasive breast cancer: Revisited. Mol. Oncol. 2013, 7, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Casasent, A.K.; Edgerton, M.; Navin, N.E. Genome evolution in ductal carcinoma in situ: Invasion of the clones. J. Pathol. 2017, 241, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Jung, S.H.; Kim, M.S.; Baek, I.P.; Lee, S.H.; Kim, T.M.; Chung, Y.J.; Lee, S.H. Genomic differences between pure ductal carcinoma in situ and synchronous ductal carcinoma in situ with invasive breast cancer. Oncotarget 2015, 6, 7597–7607. [Google Scholar] [CrossRef] [PubMed]
- Casasent, A.K.; Schalck, A.; Gao, R.; Sei, E.; Long, A.; Pangburn, W.; Casasent, T.; Meric-Bernstam, F.; Edgerton, M.E.; Navin, N.E. Multiclonal Invasion in Breast Tumors Identified by Topographic Single Cell Sequencing. Cell 2018, 172, 205–217.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, L.; Wilkerson, P.M.; Lambros, M.B.; Campion-Flora, A.; Rodrigues, D.N.; Gauthier, A.; Cabral, C.; Pawar, V.; Mackay, A.; A’Hern, R.; et al. Genomic and mutational profiling of ductal carcinomas in situ and matched adjacent invasive breast cancers reveals intra-tumour genetic heterogeneity and clonal selection. J. Pathol. 2012, 227, 42–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, H.C.; Bennett, R.P.; Kizilyer, A.; McDougall, W.M.; Prohaska, K.M. Functions and regulation of the APOBEC family of proteins. Semin. Cell Dev. Biol. 2012, 23, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.A.; Lawrence, M.S.; Klimczak, L.J.; Grimm, S.A.; Fargo, D.; Stojanov, P.; Kiezun, A.; Kryukov, G.V.; Carter, S.L.; Saksena, G.; et al. An APOBEC cytidine deaminase mutagenesis pattern is widespread in human cancers. Nat. Genet. 2013, 45, 970–976. [Google Scholar] [CrossRef]
- Harris, R.S. Molecular mechanism and clinical impact of APOBEC3B-catalyzed mutagenesis in breast cancer. Breast Cancer Res. 2015, 17, 8. [Google Scholar] [CrossRef]
- Ng, J.C.F.; Quist, J.; Grigoriadis, A.; Malim, M.H.; Fraternali, F. Pan-cancer transcriptomic analysis dissects immune and proliferative functions of APOBEC3 cytidine deaminases. Nucleic Acids Res. 2019, 47, 1178–1194. [Google Scholar] [CrossRef] [Green Version]
- Kosumi, K.; Baba, Y.; Ishimoto, T.; Harada, K.; Nakamura, K.; Ohuchi, M.; Kiyozumi, Y.; Izumi, D.; Tokunaga, R.; Taki, K.; et al. APOBEC3B is an enzymatic source of molecular alterations in esophageal squamous cell carcinoma. Med. Oncol. 2016, 33, 26. [Google Scholar] [CrossRef]
- Henderson, S.; Chakravarthy, A.; Su, X.; Boshoff, C.; Fenton, T.R. APOBEC-mediated cytosine deamination links PIK3CA helical domain mutations to human papillomavirus-driven tumor development. Cell Rep. 2014, 7, 1833–1841. [Google Scholar] [CrossRef] [PubMed]
- Burns, M.B.; Lackey, L.; Carpenter, M.A.; Rathore, A.; Land, A.M.; Leonard, B.; Refsland, E.W.; Kotandeniya, D.; Tretyakova, N.; Nikas, J.B.; et al. APOBEC3B is an enzymatic source of mutation in breast cancer. Nature 2013, 494, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Sieuwerts, A.M.; Willis, S.; Burns, M.B.; Look, M.P.; Meijer-Van Gelder, M.E.; Schlicker, A.; Heideman, M.R.; Jacobs, H.; Wessels, L.; Leyland-Jones, B.; et al. Elevated APOBEC3B correlates with poor outcomes for estrogen-receptor-positive breast cancers. Horm. Cancer 2014, 5, 405–413. [Google Scholar] [CrossRef]
- Sieuwerts, A.M.; Schrijver, W.A.; Dalm, S.U.; de Weerd, V.; Moelans, C.B.; Ter Hoeve, N.; van Diest, P.J.; Martens, J.W.; van Deurzen, C.H. Progressive APOBEC3B mRNA expression in distant breast cancer metastases. PLoS ONE 2017, 12, e0171343. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, M.; Yamane, A.; Horiguchi, J.; Yokobori, T.; Kawabata-Iwakawa, R.; Yoshiyama, S.; Rokudai, S.; Odawara, H.; Tokiniwa, H.; Oyama, T.; et al. APOBEC3B high expression status is associated with aggressive phenotype in Japanese breast cancers. Breast Cancer 2016, 23, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Siriwardena, S.U.; Chen, K.; Bhagwat, A.S. Functions and Malfunctions of Mammalian DNA-Cytosine Deaminases. Chem. Rev. 2016, 116, 12688–12710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agahozo, M.C.; Sieuwerts, A.M.; Doebar, S.C.; Verhoef, E.; Beaufort, C.M.; Ruigrok-Ritstier, K.; de Weerd, V.; Sleddens, H.; Dinjens, W.N.; Martens, J.; et al. PIK3CA mutations in ductal carcinoma in situ and adjacent invasive breast cancer. Endocr. Relat. Cancer 2019, 26, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Aubele, M.; Mattis, A.; Zitzelsberger, H.; Walch, A.; Kremer, M.; Welzl, G.; Hofler, H.; Werner, M. Extensive ductal carcinoma In situ with small foci of invasive ductal carcinoma: Evidence of genetic resemblance by CGH. Int. J. Cancer 2000, 85, 82–86. [Google Scholar] [CrossRef]
- Pang, B.; Cheng, S.; Sun, S.P.; An, C.; Liu, Z.Y.; Feng, X.; Liu, G.J. Prognostic role of PIK3CA mutations and their association with hormone receptor expression in breast cancer: A meta-analysis. Sci. Rep. 2014, 4, 6255. [Google Scholar] [CrossRef]
- Olson, M.E.; Harris, R.S.; Harki, D.A. APOBEC Enzymes as Targets for Virus and Cancer Therapy. Cell Chem. Biol. 2018, 25, 36–49. [Google Scholar] [CrossRef]
- Li, M.; Shandilya, S.M.; Carpenter, M.A.; Rathore, A.; Brown, W.L.; Perkins, A.L.; Harki, D.A.; Solberg, J.; Hook, D.J.; Pandey, K.K.; et al. First-in-class small molecule inhibitors of the single-strand DNA cytosine deaminase APOBEC3G. ACS Chem. Biol. 2012, 7, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Elston, C.W.; Ellis, I.O. Pathological prognostic factors in breast cancer. I. The value of histological grade in breast cancer: Experience from a large study with long-term follow-up. Histopathology 1991, 19, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Consensus Conference Committee. Consensus Conference on the classification of ductal carcinoma in situ. Thomas Jefferson University in Philadelphia, Pennsylvania, USA, Nov 1. Cancer 1997, 80, 1798–1802. [Google Scholar] [CrossRef]
- Prat, A.; Cheang, M.C.; Martín, M.; Parker, J.S.; Carrasco, E.; Caballero, R.; Tyldesley, S.; Gelmon, K.; Bernard, P.S.; Nielsen, T.O.; et al. Prognostic significance of progesterone receptor-positive tumor cells within immunohistochemically defined luminal A breast cancer. J. Clin. Oncol. 2013, 31, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Wolff, A.C.; Hammond, M.E.; Hicks, D.G.; Dowsett, M.; McShane, L.M.; Allison, K.H.; Allred, D.C.; Bartlett, J.M.; Bilous, M.; Fitzgibbons, P.; et al. Recommendations for human epidermal growth factor receptor 2 testing in breast cancer: American Society of Clinical Oncology/College of American Pathologists clinical practice guideline update. J. Clin. Oncol. 2013, 31, 3997–4013. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ishikawa, T.; Michiue, T.; Zhu, B.L.; Guan, D.W.; Maeda, H. Stability of endogenous reference genes in postmortem human brains for normalization of quantitative real-time PCR data: Comprehensive evaluation using geNorm, NormFinder, and BestKeeper. Int. J. Legal Med. 2012, 126, 943–952. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | n | (%) |
|---|---|---|
| Age at diagnosis | 53 | |
| years, median (range) | (28–102) | |
| Type of surgery | ||
| Breast-conserving surgery | 24 | 45.3 |
| Mastectomy | 29 | 54.7 |
| Grade DCIS | ||
| 1 | 1 | 49.1 |
| 2 | 19 | 39.6 |
| 3 | 33 | 7.5 |
| Grade IBC | ||
| 1 | 1 | 49.1 |
| 2 | 21 | 39.6 |
| 3 | 31 | 7.5 |
| Tumor size | ||
| ≤2 cm | 28 | 49.1 |
| >2–5 cm | 21 | 39.6 |
| >5 cm | 4 | 7.5 |
| Missing | 0 | 3.8 |
| Subtypes based on immunohistochemistry | ||
| ER+/PR high/Her2− | 13 | 24.5 |
| ER+/PR− or low/Her2− | 12 | 22.6 |
| ER+/any PR/Her2+ | 11 | 20.8 |
| ER−/PR−/Her2+ | 8 | 15.1 |
| ER−/PR−/Her2− | 9 | 17.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sieuwerts, A.M.; Doebar, S.C.; de Weerd, V.; Verhoef, E.I.; Beauford, C.M.; Agahozo, M.C.; Martens, J.W.M.; van Deurzen, C.H.M. APOBEC3B Gene Expression in Ductal Carcinoma In Situ and Synchronous Invasive Breast Cancer. Cancers 2019, 11, 1062. https://doi.org/10.3390/cancers11081062
Sieuwerts AM, Doebar SC, de Weerd V, Verhoef EI, Beauford CM, Agahozo MC, Martens JWM, van Deurzen CHM. APOBEC3B Gene Expression in Ductal Carcinoma In Situ and Synchronous Invasive Breast Cancer. Cancers. 2019; 11(8):1062. https://doi.org/10.3390/cancers11081062
Chicago/Turabian StyleSieuwerts, Anieta M., Shusma C. Doebar, Vanja de Weerd, Esther I. Verhoef, Corine M. Beauford, Marie C. Agahozo, John W.M. Martens, and Carolien H.M. van Deurzen. 2019. "APOBEC3B Gene Expression in Ductal Carcinoma In Situ and Synchronous Invasive Breast Cancer" Cancers 11, no. 8: 1062. https://doi.org/10.3390/cancers11081062
APA StyleSieuwerts, A. M., Doebar, S. C., de Weerd, V., Verhoef, E. I., Beauford, C. M., Agahozo, M. C., Martens, J. W. M., & van Deurzen, C. H. M. (2019). APOBEC3B Gene Expression in Ductal Carcinoma In Situ and Synchronous Invasive Breast Cancer. Cancers, 11(8), 1062. https://doi.org/10.3390/cancers11081062