Association between Circulating Fibroblast Growth Factor 21 and Aggressiveness in Thyroid Cancer

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
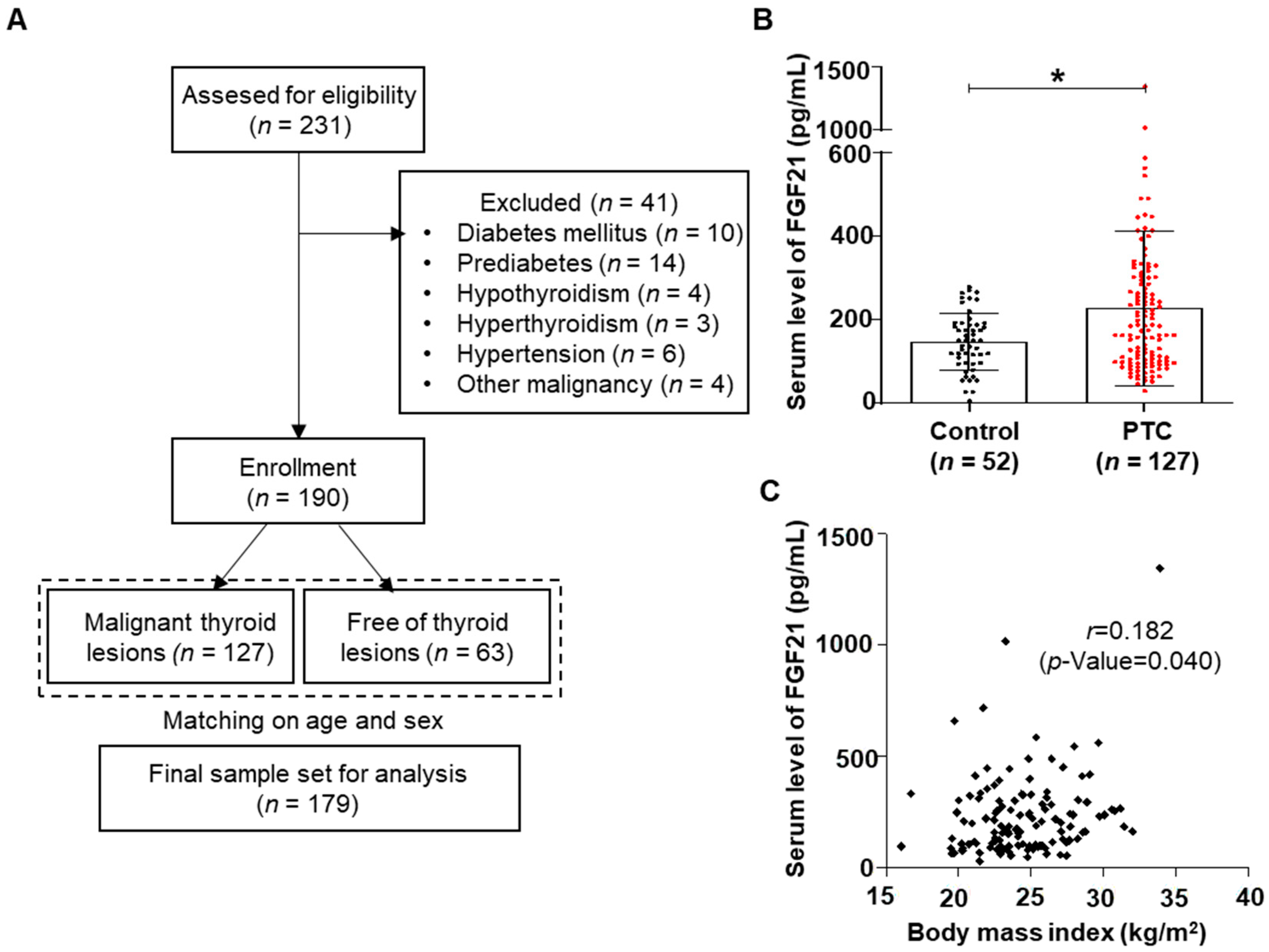
2.1. Clinical Features of PTC Patients
2.2. Higher Levels of FGF21 in Patients with PTC
2.3. Relationship between FGF21 Levels and Clinicopathological Parameters
2.4. Multivariate Analysis of the Relationships between FGF21 Levels and Tumor Aggressiveness
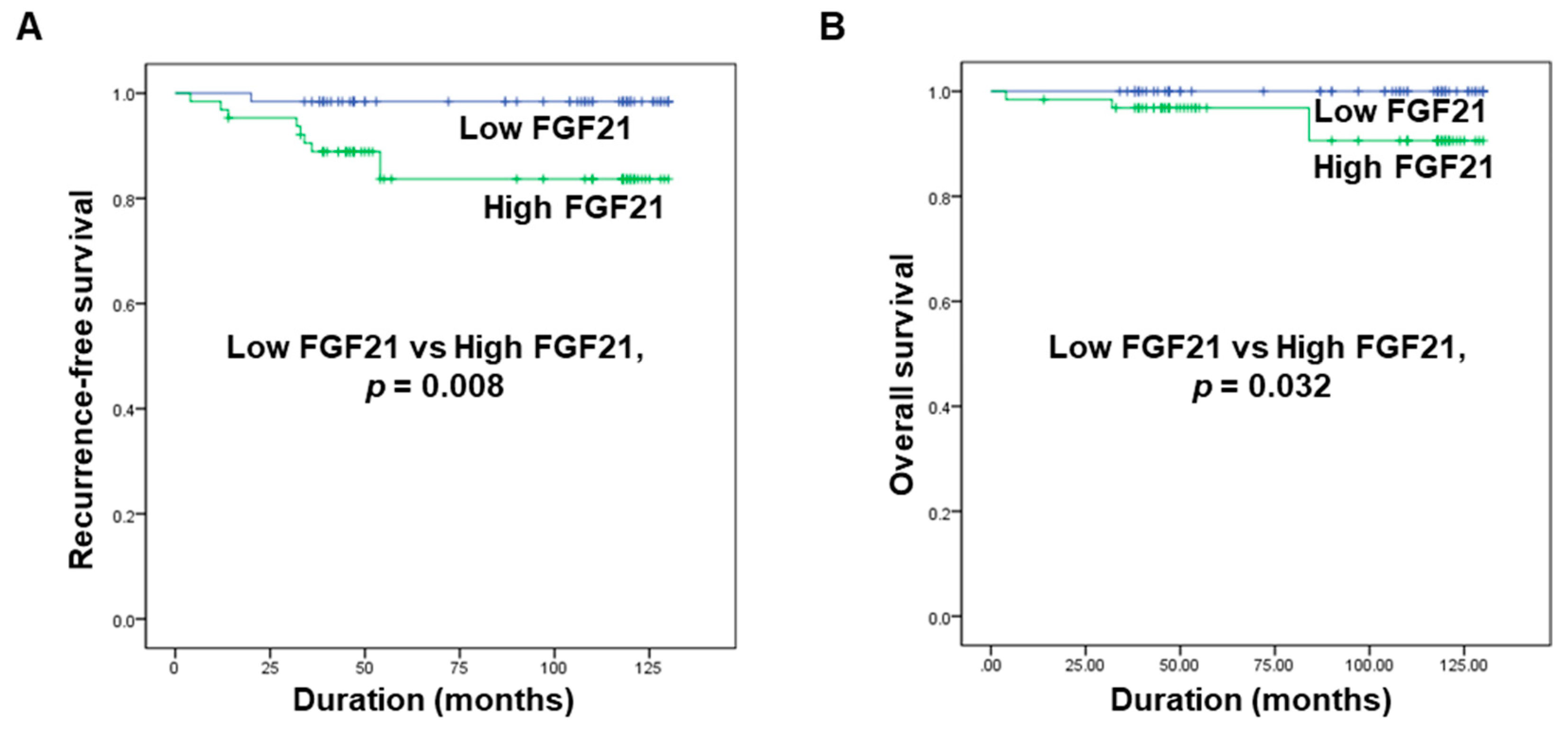
2.5. Correlations between FGF21 Levels and Recurrence-Free Survival and Overall Survival in Patients with PTC
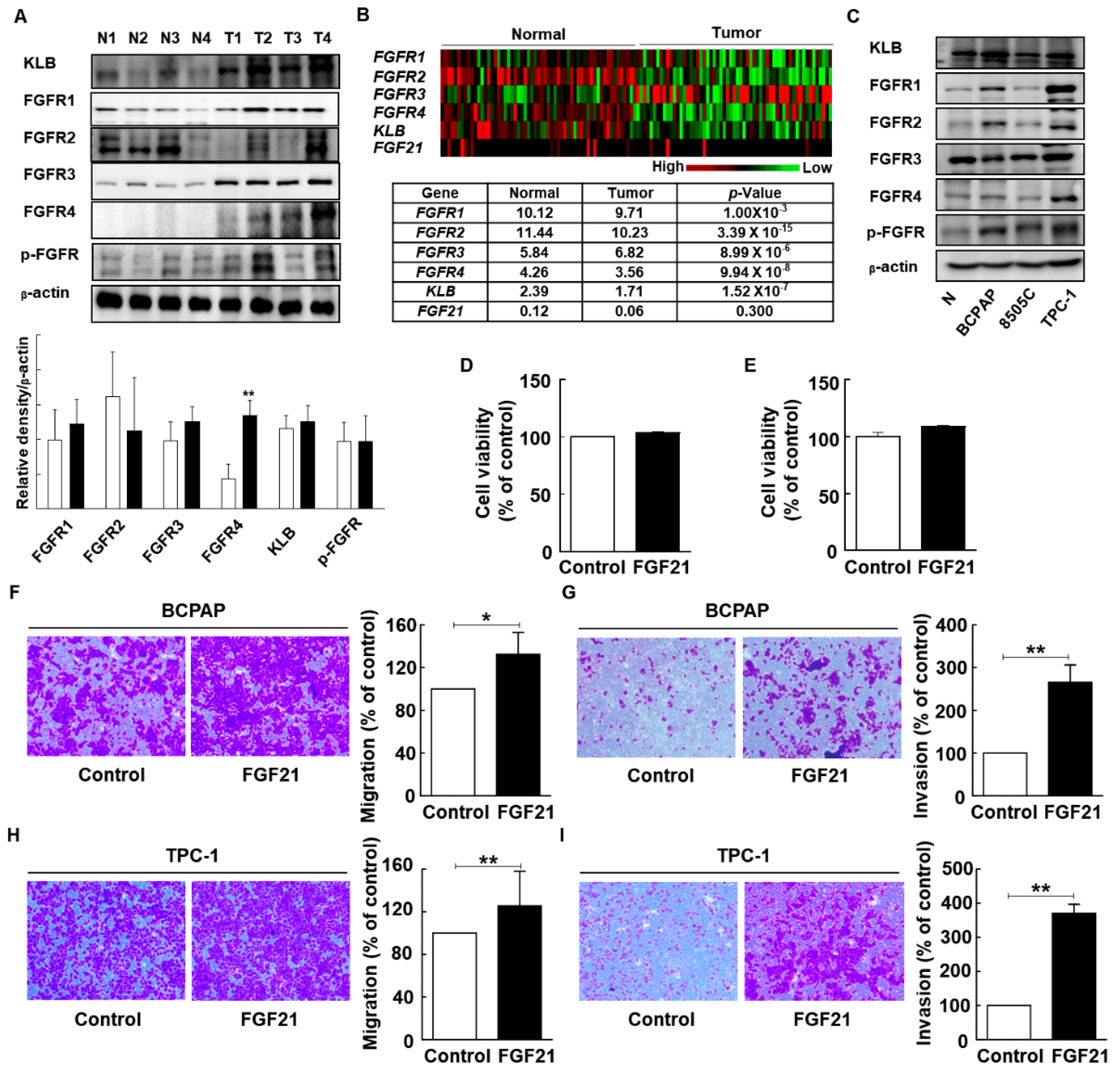
2.6. FGF21 Promotes Migration and Invasion of Thyroid Cancer Cell Lines
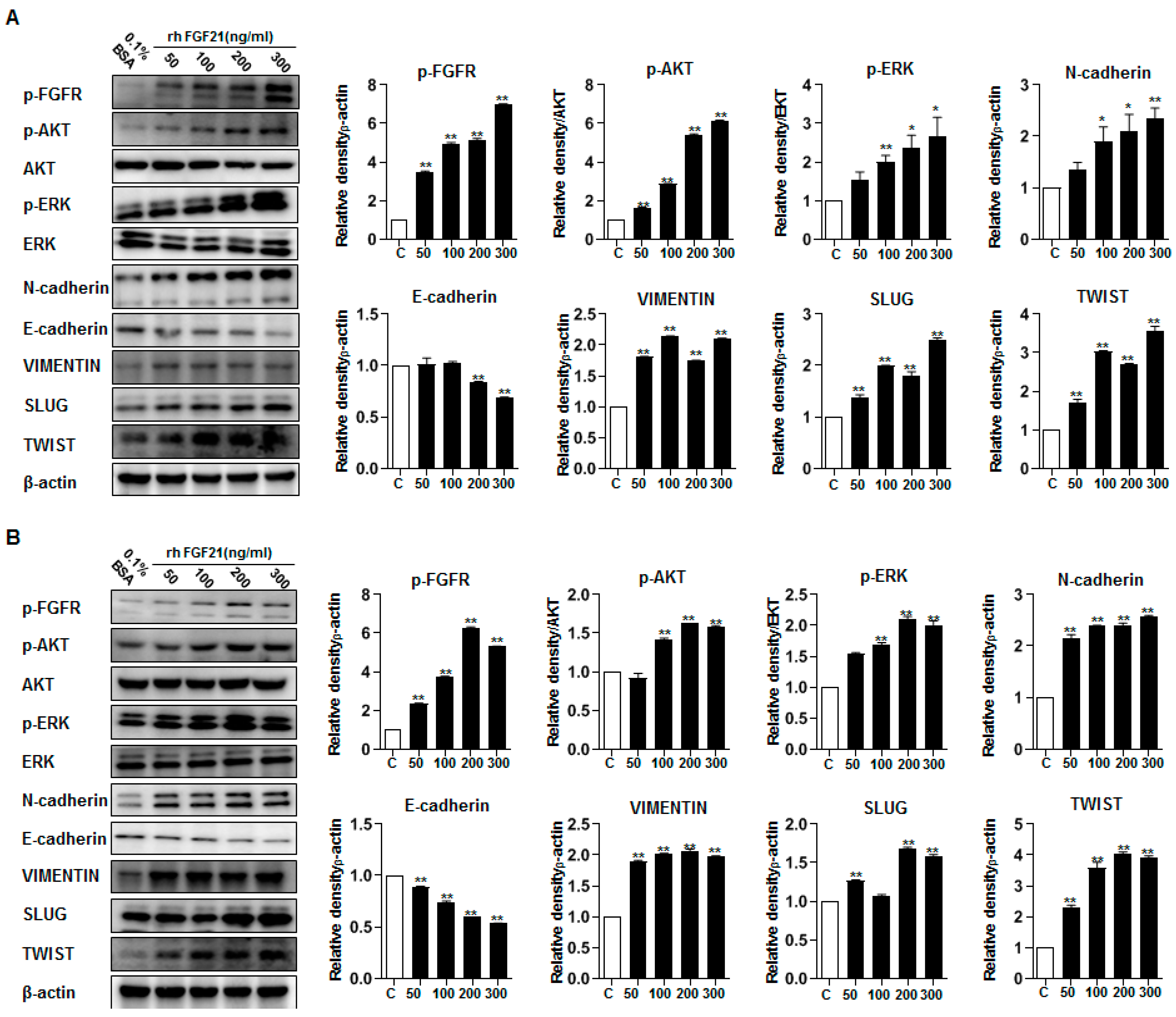
2.7. FGF21 Activates Epithelial–Mesenchymal Transition (EMT) Signaling and Upregulates FGFR Signaling
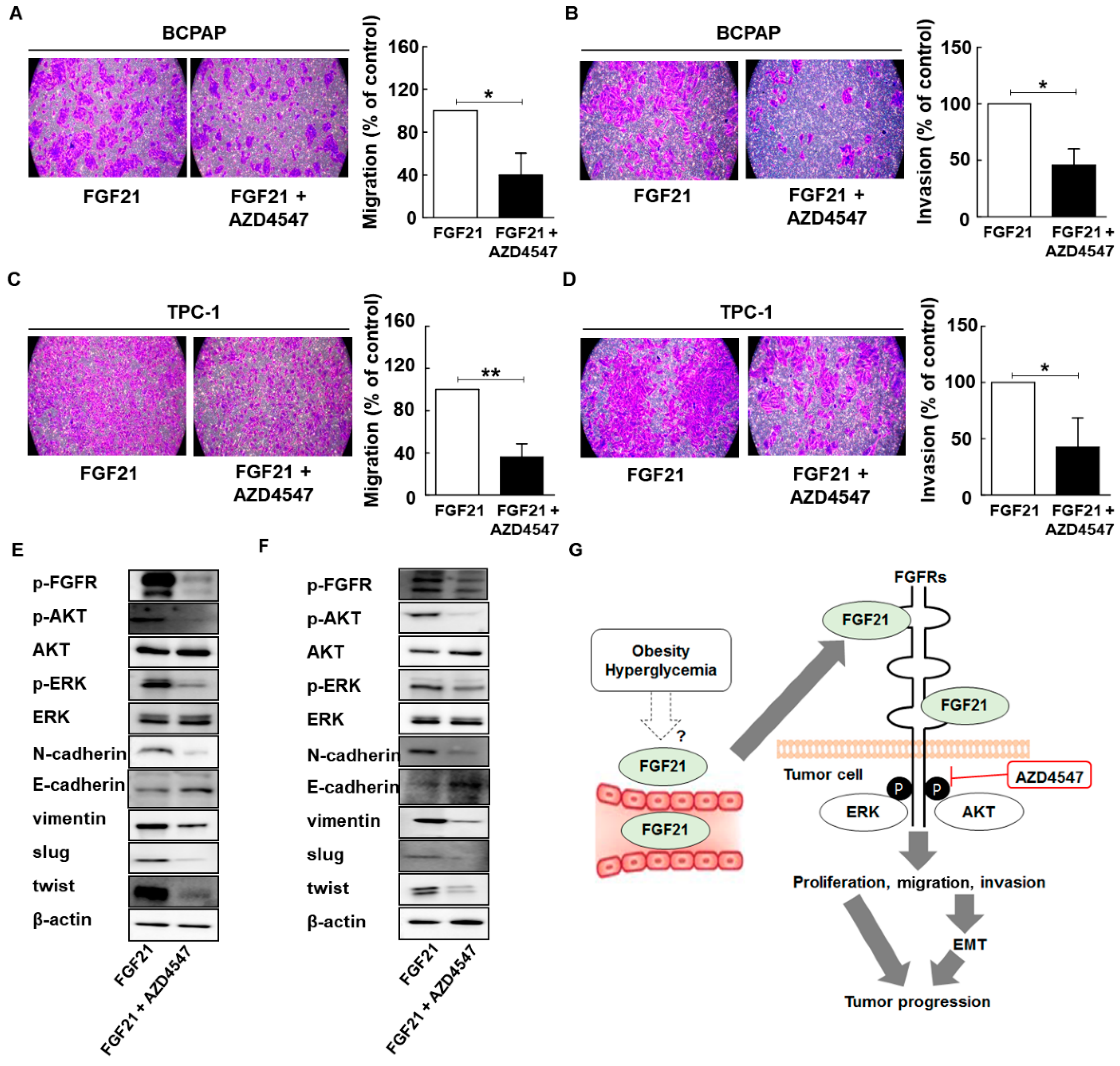
2.8. AZD4547 Attenuates the Effects of rFGF21 in Thyroid Cancer via Downregulation of FGFR Signaling
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Clinical and Biochemical Parameters
4.3. Serum Levels of FGF21
4.4. Genomic and Clinical Data Sets
4.5. Cell Lines and Cell Cultures
4.6. Recombinant FGF21 Treatment
4.7. Western Blot Analysis
4.8. Cell Viability Assay
4.9. Cell Migration and Invasion Assay
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dieci, M.V.; Arnedos, M.; Andre, F.; Soria, J.C. Fibroblast growth factor receptor inhibitors as a cancer treatment: From a biologic rationale to medical perspectives. Cancer Discov. 2013, 3, 264–279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ibrahimi, O.A.; Olsen, S.K.; Umemori, H.; Mohammadi, M.; Ornitz, D.M. Receptor specificity of the fibroblast growth factor family, part II. J. Biol. Chem. 2006, 271, 15292–15297. [Google Scholar]
- Turner, N.; Grose, R. Fibroblast growth factor signalling: From development to cancer. Nat. Rev. Cancer 2010, 10, 116. [Google Scholar] [CrossRef] [PubMed]
- Wesche, J.; Haglund, K.; Haugsten, E.M. Fibroblast growth factors and their receptors in cancer. Biochem. J. 2011, 437, 199–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo, V.D.; Gangula, R.D.; Freeman, K.W.; Li, R.; Zhang, Y.; Wang, F.; Ayala, G.E.; Peterson, L.E.; Ittmann, M.; Spencer, D.M. Inducible FGFR-1 activation leads to irreversible prostate adenocarcinoma and an epithelial-to-mesenchymal transition. Cancer Cell 2007, 12, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Marek, L.; Ware, K.E.; Fritzsche, A.; Hercule, P.; Helton, W.R.; Smith, J.E.; McDermott, L.A.; Coldren, C.D.; Nemenoff, R.A.; Merrick, D.T. Fibroblast growth factor (FGF) and FGF receptor-mediated autocrine signaling in non-small-cell lung cancer cells. Mol. Pharmacol. 2009, 75, 196–207. [Google Scholar] [CrossRef]
- Beenken, A.; Mohammadi, M. The FGF family: Biology, pathophysiology and therapy. Nat. Rev. Drug Discov. 2009, 8, 235. [Google Scholar] [CrossRef]
- Inagaki, T.; Dutchak, P.; Zhao, G.; Ding, X.; Gautron, L.; Parameswara, V.; Li, Y.; Goetz, R.; Mohammadi, M.; Esser, V. Endocrine regulation of the fasting response by PPARα-mediated induction of fibroblast growth factor 21. Cell Metabol. 2007, 5, 415–425. [Google Scholar] [CrossRef]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.J.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A. FGF-21 as a novel metabolic regulator. J. Clin. Investig. 2005, 115, 1627–1635. [Google Scholar] [CrossRef] [Green Version]
- Foltz, I.N.; Hu, S.; King, C.; Wu, X.; Yang, C.; Wang, W.; Weiszmann, J.; Stevens, J.; Chen, J.S.; Nuanmanee, N. Treating diabetes and obesity with an FGF21-mimetic antibody activating the βKlotho/FGFR1c receptor complex. Sci. Transl. Med. 2012, 4, ra153–ra162. [Google Scholar] [CrossRef]
- Yang, C.; Jin, C.; Li, X.; Wang, F.; McKeehan, W.L.; Luo, Y. Differential specificity of endocrine FGF19 and FGF21 to FGFR1 and FGFR4 in complex with KLB. PLoS ONE 2012, 7, e33870. [Google Scholar] [CrossRef]
- Yang, C.; Wang, C.; Ye, M.; Jin, C.; He, W.; Wang, F.; McKeehan, W.L.; Luo, Y. Control of lipid metabolism by adipocyte FGFR1-mediated adipohepatic communication during hepatic stress. Nutr. Metabol. 2012, 9, 94. [Google Scholar] [CrossRef]
- Kurosu, H.; Kuro-o, M. The Klotho gene family as a regulator of endocrine fibroblast growth factors. Mol. Cell. Endocrinol. 2009, 299, 72–78. [Google Scholar] [CrossRef]
- Badman, M.K.; Pissios, P.; Kennedy, A.R.; Koukos, G.; Flier, J.S.; Maratos-Flier, E. Hepatic fibroblast growth factor 21 is regulated by PPARα and is a key mediator of hepatic lipid metabolism in ketotic states. Cell Metabol. 2007, 5, 426–437. [Google Scholar] [CrossRef]
- Lundåsen, T.; Hunt, M.C.; Nilsson, L.-M.; Sanyal, S.; Angelin, B.; Alexson, S.E.; Rudling, M. PPARα is a key regulator of hepatic FGF21. Biochem. Biophys. Res. Commun. 2007, 360, 437–440. [Google Scholar] [CrossRef]
- Mai, K.; Andres, J.; Biedasek, K.; Weicht, J.; Bobbert, T.; Sabath, M.; Meinus, S.; Reinecke, F.; Möhlig, M.; Weickert, M.O. Free fatty acids link metabolism and regulation of the insulin-sensitizing fibroblast growth factor-21. Diabetes 2009, 58, 1532–1538. [Google Scholar] [CrossRef]
- Hondares, E.; Iglesias, R.; Giralt, A.; Gonzalez, F.J.; Giralt, M.; Mampel, T.; Villarroya, F. Thermogenic activation induces FGF21 expression and release in brown adipose tissue. J. Biol. Chem. 2011, 286, 12983–12990. [Google Scholar] [CrossRef]
- Muise, E.S.; Kuo, D.W.; El-Sherbeini, M.; Tan, Y.; Yuan, X.; Mu, J.; Thompson, J.R.; Berger, J.P.; Wong, K.K. Adipose fibroblast growth factor 21 is up-regulated by PPARγ and altered metabolic states. Mol. Pharmacol. 2008, 74, 403–412. [Google Scholar] [CrossRef]
- Izumiya, Y.; Bina, H.A.; Ouchi, N.; Akasaki, Y.; Kharitonenkov, A.; Walsh, K. FGF21 is an Akt-regulated myokine. FEBS Lett. 2008, 582, 3805–3810. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Lu, W.; Lin, T.; You, P.; Ye, M.; Huang, Y.; Jiang, X.; Wang, C.; Wang, F.; Lee, M.-H. Activation of Liver FGF21 in hepatocarcinogenesis and during hepatic stress. BMC Gastroenterol. 2013, 13, 67. [Google Scholar] [CrossRef]
- Kim, K.H.; Jeong, Y.T.; Oh, H.; Kim, S.H.; Cho, J.M.; Kim, Y.-N.; Kim, S.S.; Kim, D.H.; Hur, K.Y.; Kim, H.K. Autophagy deficiency leads to protection from obesity and insulin resistance by inducing Fgf21 as a mitokine. Nat. Med. 2013, 19, 83. [Google Scholar] [CrossRef]
- Schaap, F.G.; Kremer, A.E.; Lamers, W.H.; Jansen, P.L.; Gaemers, I.C. Fibroblast growth factor 21 is induced by endoplasmic reticulum stress. Biochimie 2013, 95, 692–699. [Google Scholar] [CrossRef]
- Suomalainen, A.; Elo, J.M.; Pietiläinen, K.H.; Hakonen, A.H.; Sevastianova, K.; Korpela, M.; Isohanni, P.; Marjavaara, S.K.; Tyni, T.; Kiuru-Enari, S. FGF-21 as a biomarker for muscle-manifesting mitochondrial respiratory chain deficiencies: A diagnostic study. Lancet Neurol. 2011, 10, 806–818. [Google Scholar] [CrossRef]
- Crasto, C.; Semba, R.D.; Sun, K.; Ferrucci, L. Serum fibroblast growth factor 21 is associated with renal function and chronic kidney disease in community-dwelling adults. J. Am. Geriatr. Soc. 2012, 60, 792. [Google Scholar] [CrossRef]
- Han, S.H.; Choi, S.H.; Cho, B.J.; Lee, Y.; Lim, S.; Park, Y.J.; Moon, M.K.; Lee, H.K.; Kang, S.-W.; Han, D.S. Serum fibroblast growth factor–21 concentration is associated with residual renal function and insulin resistance in end-stage renal disease patients receiving long-term peritoneal dialysis. Metabol. Clin. Exp. 2010, 59, 1656–1662. [Google Scholar] [CrossRef]
- Lin, Z.; Zhou, Z.; Liu, Y.; Gong, Q.; Yan, X.; Xiao, J.; Wang, X.; Lin, S.; Feng, W.; Li, X. Circulating FGF21 levels are progressively increased from the early to end stages of chronic kidney diseases and are associated with renal function in Chinese. PLoS ONE 2011, 6, e18398. [Google Scholar] [CrossRef]
- Chou, R.-H.; Huang, P.-H.; Hsu, C.-Y.; Chang, C.-C.; Leu, H.-B.; Huang, C.-C.; Chen, J.-W.; Lin, S.-J. Circulating fibroblast growth factor 21 is associated with diastolic dysfunction in heart failure patients with preserved ejection fraction. Sci. Rep. 2016, 6, 33953. [Google Scholar] [CrossRef]
- Yaqoob, U.; Jagavelu, K.; Shergill, U.; De Assuncao, T.; Cao, S.; Shah, V.H. FGF21 promotes endothelial cell angiogenesis through a dynamin-2 and Rab5 dependent pathway. PLoS ONE 2014, 9, e98130. [Google Scholar] [CrossRef]
- Dey, J.H.; Bianchi, F.; Voshol, J.; Bonenfant, D.; Oakeley, E.J.; Hynes, N.E. Targeting fibroblast growth factor receptors blocks PI3K/AKT signaling, induces apoptosis, and impairs mammary tumor outgrowth and metastasis. Cancer Res. 2010, 70, 4151–4162. [Google Scholar] [CrossRef]
- Koziczak, M.; Holbro, T.; Hynes, N.E. Blocking of FGFR signaling inhibits breast cancer cell proliferation through downregulation of D-type cyclins. Oncogene 2004, 23, 3501. [Google Scholar] [CrossRef]
- Stjepanovic, N.; Capdevila, J. Multikinase inhibitors in the treatment of thyroid cancer: Specific role of lenvatinib. Biol. Targets Ther. 2014, 8, 129. [Google Scholar]
- St. Bernard, R.; Zheng, L.; Liu, W.; Winer, D.; Asa, S.L.; Ezzat, S. Fibroblast growth factor receptors as molecular targets in thyroid carcinoma. Endocrinology 2005, 146, 1145–1153. [Google Scholar] [CrossRef]
- Shingu, K.; Fujimori, M.; Ito, K.-I.; Hama, Y.; Kasuga, Y.; Kobayashi, S.; Itoh, N.; Amano, J. Expression of fibroblast growth factor-2 and fibroblast growth factor receptor-1 in thyroid diseases. Endocr. J. 1998, 45, 35–43. [Google Scholar] [CrossRef]
- Nishimura, T.; Nakatake, Y.; Konishi, M.; Itoh, N. Identification of a novel FGF, FGF-21, preferentially expressed in the liver1. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 2000, 1492, 203–206. [Google Scholar] [CrossRef]
- Singhal, G.; Chee, M.J.; Tan, T.G.; El Ouaamari, A.; Adams, A.C.; Najarian, R.; Kulkarni, R.N.; Benoist, C.; Flier, J.S.; Maratos-Flier, E. Fibroblast growth factor 21 (FGF21) protects against high fat diet induced inflammation and islet hyperplasia in pancreas. PLoS ONE 2016, 11, e0148252. [Google Scholar] [CrossRef]
- Lin, Z.; Pan, X.; Wu, F.; Ye, D.; Zhang, Y.; Wang, Y.; Jin, L.; Lian, Q.; Huang, Y.; Ding, H. Fibroblast growth factor 21 prevents atherosclerosis by suppression of hepatic sterol regulatory element-binding protein-2 and induction of adiponectin in mice. Circulation 2015, 131, 1861–1871. [Google Scholar] [CrossRef]
- Ye, D.; Wang, Y.; Li, H.; Jia, W.; Man, K.; Lo, C.M.; Wang, Y.; Lam, K.S.; Xu, A. Fibroblast growth factor 21 protects against acetaminophen-induced hepatotoxicity by potentiating peroxisome proliferator-activated receptor coactivator protein-1α-mediated antioxidant capacity in mice. Hepatology 2014, 60, 977–989. [Google Scholar] [CrossRef]
- Coate, K.C.; Hernandez, G.; Thorne, C.A.; Sun, S.; Le, T.D.; Vale, K.; Kliewer, S.A.; Mangelsdorf, D.J. FGF21 is an exocrine pancreas secretagogue. Cell Metabol. 2017, 25, 472–480. [Google Scholar] [CrossRef]
- Owen, B.M.; Mangelsdorf, D.J.; Kliewer, S.A. Tissue-specific actions of the metabolic hormones FGF15/19 and FGF21. Trends Endocrinol. Metabol. 2015, 26, 22–29. [Google Scholar] [CrossRef]
- Tyynismaa, H.; Carroll, C.J.; Raimundo, N.; Ahola-Erkkilä, S.; Wenz, T.; Ruhanen, H.; Guse, K.; Hemminki, A.; Peltola-Mjøsund, K.E.; Tulkki, V. Mitochondrial myopathy induces a starvation-like response. Hum. Mol. Genet. 2010, 19, 3948–3958. [Google Scholar] [CrossRef] [Green Version]
- Hojman, P.; Pedersen, M.; Nielsen, A.R.; Krogh-Madsen, R.; Yfanti, C.; Åkerstrom, T.; Nielsen, S.; Pedersen, B.K. Fibroblast growth factor-21 is induced in human skeletal muscles by hyperinsulinemia. Diabetes 2009, 58, 2797–2801. [Google Scholar] [CrossRef]
- Lee, M.H.; Lee, S.E.; Kim, D.W.; Ryu, M.J.; Kim, S.J.; Kim, S.J.; Kim, Y.K.; Park, J.H.; Kweon, G.R.; Kim, J.M. Mitochondrial localization and regulation of BRAFV600E in thyroid cancer: A clinically used RAF inhibitor is unable to block the mitochondrial activities of BRAFV600E. J. Clin. Endocrinol. Metabol. 2011, 96, E19–E30. [Google Scholar] [CrossRef]
- Yi, H.-S.; Chang, J.Y.; Kim, K.S.; Shong, M. Oncogenes, mitochondrial metabolism, and quality control in differentiated thyroid cancer. Korean J. Internal Med. 2017, 32, 780. [Google Scholar] [CrossRef]
- Zhao, Z.; Guo, X.; Ba, C.; Wang, W.; Yang, Y.; Wang, J.; Cao, H. Overweight, obesity and thyroid cancer risk: A meta-analysis of cohort studies. J. Int. Med. Res. 2012, 40, 2041–2050. [Google Scholar] [CrossRef]
- Kitahara, C.M.; Platz, E.A.; Freeman, L.E.B.; Hsing, A.W.; Linet, M.S.; Park, Y.; Schairer, C.; Schatzkin, A.; Shikany, J.M.; de González, A.B. Obesity and thyroid cancer risk among US men and women: A pooled analysis of five prospective studies. Cancer Epidemiol. Prev. Biomark 2011, 20, 464–472. [Google Scholar] [CrossRef]
- Markan, K.R.; Naber, M.C.; Ameka, M.K.; Anderegg, M.D.; Mangelsdorf, D.J.; Kliewer, S.A.; Mohammadi, M.; Potthoff, M.J. Circulating FGF21 is liver derived and enhances glucose uptake during refeeding and overfeeding. Diabetes 2014, 63, 4057–4063. [Google Scholar] [CrossRef]
- Knott, M.E.; Minatta, J.N.; Roulet, L.; Gueglio, G.; Pasik, L.; Ranuncolo, S.M.; Nuñez, M.; Puricelli, L.I.; De Lorenzo, M.S. Circulating fibroblast growth factor 21 (Fgf21) as diagnostic and prognostic biomarker in renal cancer. J. Mol. Biomark. Diagn. 2016, 1, 015. [Google Scholar]
- Knott, M.E.; Ranuncolo, S.M.; Nuñez, M.; Armanasco, E.; Puricelli, L.I.; De Lorenzo, M.S. Levels of Fibroblast Growth Factor 21 (FGF21) in serum as diagnostic biomarker in patients with breast cancer. Clin. Res. 2015. [Google Scholar] [CrossRef]
- Gaich, G.; Chien, J.Y.; Fu, H.; Glass, L.C.; Deeg, M.A.; Holland, W.L.; Kharitonenkov, A.; Bumol, T.; Schilske, H.K.; Moller, D.E. The effects of LY2405319, an FGF21 analog, in obese human subjects with type 2 diabetes. Cell Metabol. 2013, 18, 333–340. [Google Scholar] [CrossRef]
- Adams, A.C.; Yang, C.; Coskun, T.; Cheng, C.C.; Gimeno, R.E.; Luo, Y.; Kharitonenkov, A. The breadth of FGF21’s metabolic actions are governed by FGFR1 in adipose tissue. Mol. Metabol. 2013, 2, 31–37. [Google Scholar] [CrossRef]
- Motylewska, E.; Stępień, T.; Borkowska, M.; Kuzdak, K.; Siejka, A.; Komorowski, J.; Stępień, H.; Ławnicka, H. Alteration in the serum concentrations of FGF19, FGFR4 and βKlotho in patients with thyroid cancer. Cytokine 2018, 105, 32–36. [Google Scholar] [CrossRef]
- Pai, R.; Dunlap, D.; Qing, J.; Mohtashemi, I.; Hotzel, K.; French, D.M. Inhibition of fibroblast growth factor 19 reduces tumor growth by modulating β-catenin signaling. Cancer Res. 2008, 68, 5086–5095. [Google Scholar] [CrossRef]
- Nicholes, K.; Guillet, S.; Tomlinson, E.; Hillan, K.; Wright, B.; Frantz, G.D.; Pham, T.A.; Dillard-Telm, L.; Tsai, S.P.; Stephan, J.-P. A mouse model of hepatocellular carcinoma: Ectopic expression of fibroblast growth factor 19 in skeletal muscle of transgenic mice. Am. J. Pathol. 2002, 160, 2295–2307. [Google Scholar] [CrossRef]
- Xie, M.-H.; Holcomb, I.; Deuel, B.; Dowd, P.; Huang, A.; Vagts, A.; Foster, J.; Liang, J.; Brush, J.; Gu, Q. FGF-19, a novel fibroblast growth factor with unique specificity for FGFR4. Cytokine 1999, 11, 729–735. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, Y.; Berglund, E.D.; Coate, K.C.; He, T.T.; Katafuchi, T.; Xiao, G.; Potthoff, M.J.; Wei, W.; Wan, Y. The starvation hormone, fibroblast growth factor-21, extends lifespan in mice. Elife 2012, 1, e00065. [Google Scholar] [CrossRef]
- Ables, G.P.; Perrone, C.E.; Orentreich, D.; Orentreich, N. Methionine-restricted C57BL/6J mice are resistant to diet-induced obesity and insulin resistance but have low bone density. PLoS ONE 2012, 7, e51357. [Google Scholar] [CrossRef]
- Harlid, S.; Myte, R.; Van Guelpen, B. The metabolic syndrome, inflammation, and colorectal cancer risk: An evaluation of large panels of plasma protein markers using repeated, prediagnostic samples. Mediat. Inflamm. 2017, 2017, 4803156. [Google Scholar] [CrossRef]
- Babina, I.S.; Turner, N.C. Advances and challenges in targeting FGFR signalling in cancer. Nat. Rev. Cancer 2017, 17, 318. [Google Scholar] [CrossRef]
- Dienstmann, R.; Rodon, J.; Prat, A.; Perez-Garcia, J.; Adamo, B.; Felip, E.; Cortes, J.; Iafrate, A.; Nuciforo, P.; Tabernero, J. Genomic aberrations in the FGFR pathway: Opportunities for targeted therapies in solid tumors. Ann. Oncol. 2013, 25, 552–563. [Google Scholar] [CrossRef]
- Schlumberger, M.; Tahara, M.; Wirth, L.J.; Robinson, B.; Brose, M.S.; Elisei, R.; Habra, M.A.; Newbold, K.; Shah, M.H.; Hoff, A.O. Lenvatinib versus placebo in radioiodine-refractory thyroid cancer. New Engl. J. Med. 2015, 372, 621–630. [Google Scholar] [CrossRef]
- Matsui, J.; Funahashi, Y.; Uenaka, T.; Watanabe, T.; Tsuruoka, A.; Asada, M. Multi-kinase inhibitor E7080 suppresses lymph node and lung metastases of human mammary breast tumor MDA-MB-231 via inhibition of vascular endothelial growth factor-receptor (VEGF-R) 2 and VEGF-R3 kinase. Clin. Cancer Res. 2008, 14, 5459–5465. [Google Scholar] [CrossRef]
- Grande, E.; Díez, J.J.; Zafon, C.; Capdevila, J. Thyroid cancer: Molecular aspects and new therapeutic strategies. J. Thyroid Res. 2012, 2012, 847108. [Google Scholar] [CrossRef]
- Abbas, A.K.; Murphy, K.M.; Sher, A. Functional diversity of helper T lymphocytes. Nature 1996, 383, 787. [Google Scholar] [CrossRef]
- TCGA Data Portal. Available online: https://tcga-data.nci.nih.gov (accessed on 1 January 2018).
- Cancer Browser. Available online: https://genome-cancer.ucsc.edu (accessed on 1 January 2018).
- Chang, J.W.; Jung, S.-N.; Kim, J.-H.; Shim, G.-A.; Park, H.S.; Liu, L.; Kim, J.M.; Park, J.; Koo, B.S. Carboxyl-terminal modulator protein positively acts as an oncogenic driver in head and neck squamous cell carcinoma via regulating Akt phosphorylation. Sci. Rep. 2016, 6, 28503. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Control Group (n = 52) | PTC Group (n = 127) | p-Value |
|---|---|---|---|
| Age (years) | 52.0 ± 12.3 | 50.5 ± 12.2 | 0.488 |
| Sex (male:female) | 9:31 | 18:109 | 0.212 |
| Body weight (kg) | 62.5 ± 11.4 | 61.2 ± 9.5 | 0.519 |
| Height (cm) | 160.4 ± 9.5 | 158.0 ± 8.1 | 0.102 |
| BMI (kg/m2) | 23.5 ± 2.7 | 24.5 ± 3.1 | 0.060 |
| Systolic blood pressure (mmHg) | 118.7 ± 9.4 | 117.7 ± 8.0 | 0.053 |
| Diastolic blood pressure (mmHg) | 74.0 ± 8.9 | 75.2 ± 5.9 | 0.318 |
| Fasting glucose (mm) | 99.9 ± 9.9 | 96.4 ± 15.4 | 0.174 |
| Triglycerides (mm) | 137.2 ± 116.1 | 136.2 ± 83.7 | 0.961 |
| Total cholesterol (mm) | 184.1 ± 32.7 | 187.9 ± 36.7 | 0.532 |
| LDL cholesterol (mm) | 111.3 ± 29.9 | 112.1 ± 36.0 | 0.895 |
| HDL cholesterol (mm) | 52.8 ± 13.1 | 51.6 ± 12.4 | 0.623 |
| AST (IU/L) | 21.1 ± 6.2 | 19.5 ± 7.2 | 0.185 |
| ALT (IU/L) | 20.3 ± 14.1 | 20.3 ± 12.4 | 0.895 |
| TSH (IU/mL) | 2.2 ± 1.9 | 2.2 ± 3.8 | 0.993 |
| Serum level of FGF21 (pg/mL) | 147.4 ± 68.7 | 227.2 ± 184.0 | 0.017 * |
| Variables | Low FGF21 Group (n = 63) | High FGF21 Group (n = 64) | p-Value | |
|---|---|---|---|---|
| Age (years) | <45 | 22 | 20 | 0.66 |
| ≥45 | 41 | 44 | ||
| Sex | Male | 6 | 12 | 0.136 |
| Female | 57 | 52 | ||
| Tumor size (cm) | ≤2 | 57 | 51 | 0.088 |
| >2 | 6 | 13 | ||
| T stage | T1–T2 | 33 | 15 | 0.001 * |
| T3–T4 | 30 | 49 | ||
| Multicentricity | No | 47 | 46 | 0.728 |
| Yes | 16 | 18 | ||
| Microscopic capsular invasion | No | 27 | 14 | 0.011 * |
| Yes | 36 | 50 | ||
| Extrathyroid extension | No | 37 | 21 | 0.003 * |
| Yes | 26 | 43 | ||
| Lymphovascular invasion | No | 15 | 12 | 0.486 |
| Yes | 48 | 52 | ||
| Central lymph node metastasis | No | 42 | 43 | 0.95 |
| Yes | 21 | 21 | ||
| Lateral lymph node metastasis | No | 57 | 57 | 0.793 |
| Yes | 6 | 6 | ||
| Recurrence | No | 62 | 56 | 0.017 * |
| Yes | 1 | 8 | ||
| Survival | No | 0 | 4 | 0.044 * |
| Yes | 63 | 60 | ||
| BRAFV600E mutation | No | 2 | 1 | 0.09 |
| Yes | 5 | 15 | ||
| Unknown | 55 | 48 | ||
| Body weight (kg) | 48.5 ± 12.1 | 52.45 ± 12.1 | 0.068 | |
| Height (cm) | 158.2 ± 6.7 | 157.8 ± 9.4 | 0.773 | |
| BMI (kg/m2) | 22.8 ± 2.8 | 25.1 ± 3.3 | 0.013 * | |
| Fasting glucose (mmol/L) | 93.5 ± 12.3 | 99.3 ± 17.5 | 0.031 * | |
| Triglycerides (mmol/L) | 118.7 ± 70.8 | 153.7 ± 92.1 | 0.026 * | |
| Total cholesterol (mmol/L) | 187.2 ± 40.7 | 188.7 ± 32.5 | 0.825 | |
| LDL cholesterol (mmol/L) | 118.5 ± 35.4 | 105.5 ± 35.6 | 0.079 | |
| HDL cholesterol (mmol/L) | 53.8 ± 12.8 | 49.2 ± 11.8 | 0.071 | |
| AST (IU/L) | 19.5 ± 8.2 | 19.6 ± 6.2 | 0.918 | |
| ALT (IU/L) | 19.1 ± 11.0 | 21.5 ± 13.7 | 0.279 | |
| TSH (IU/mL) | 2.6 ± 5.2 | 1.9 ± 1.4 | 0.283 | |
| Factors | Exp (β) | SE | 95% CI | p-Value |
|---|---|---|---|---|
| T stage (stage 3–4) | 3.671 | 0.776 | (0.802, 16.811) | 0.094 |
| Microscopic capsular invasion | 0.907 | 0.630 | (0.264, 3.120) | 0.877 |
| Extrathyroid extension | 1.121 | 0.637 | (0.321, 3.908) | 0.858 |
| Recurrence | 9.985 | 1.110 | (1.135, 87.868) | 0.038 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, Y.E.; Kim, J.T.; Lim, M.A.; Oh, C.; Liu, L.; Jung, S.-N.; Won, H.-R.; Lee, K.; Chang, J.W.; Yi, H.-S.; et al. Association between Circulating Fibroblast Growth Factor 21 and Aggressiveness in Thyroid Cancer. Cancers 2019, 11, 1154. https://doi.org/10.3390/cancers11081154
Kang YE, Kim JT, Lim MA, Oh C, Liu L, Jung S-N, Won H-R, Lee K, Chang JW, Yi H-S, et al. Association between Circulating Fibroblast Growth Factor 21 and Aggressiveness in Thyroid Cancer. Cancers. 2019; 11(8):1154. https://doi.org/10.3390/cancers11081154
Chicago/Turabian StyleKang, Yea Eun, Jung Tae Kim, Mi Ae Lim, Chan Oh, Lihua Liu, Seung-Nam Jung, Ho-Ryun Won, Kyungmin Lee, Jae Won Chang, Hyon-Seung Yi, and et al. 2019. "Association between Circulating Fibroblast Growth Factor 21 and Aggressiveness in Thyroid Cancer" Cancers 11, no. 8: 1154. https://doi.org/10.3390/cancers11081154
APA StyleKang, Y. E., Kim, J. T., Lim, M. A., Oh, C., Liu, L., Jung, S. -N., Won, H. -R., Lee, K., Chang, J. W., Yi, H. -S., Kim, H. J., Ku, B. J., Shong, M., & Koo, B. S. (2019). Association between Circulating Fibroblast Growth Factor 21 and Aggressiveness in Thyroid Cancer. Cancers, 11(8), 1154. https://doi.org/10.3390/cancers11081154