p21 in Cancer Research

Abstract
:1. p21 and Cancer
1.1. p21 in Early Days
1.2. p21 and Cancer Evolution
1.3. LincRNA-p21 and Cancer
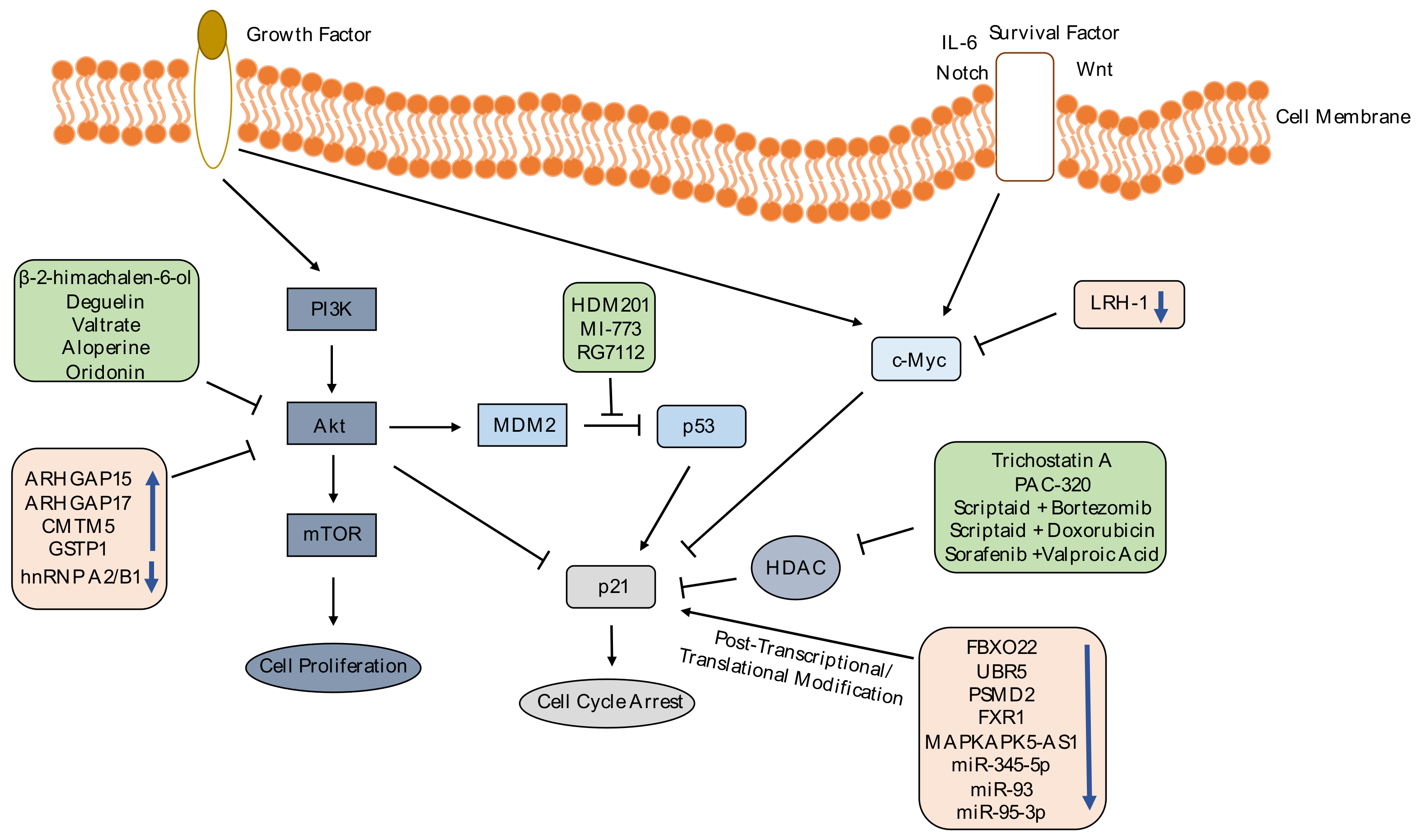
2. p21 as a Target in Cancer Treatment
2.1. Gene Editing of p21 in Cancer Cells
2.2. Targeting p21 Pathways
2.2.1. Chemicals
2.2.2. Gene Editing
2.2.3. Synthetic Lethality in p21-Mutant Cancers
3. p21 Regulation
3.1. Strict p21 Regulation
3.2. Targeting p21 Regulators for Therapy
4. p21 Role in Stemness
4.1. p21 Expression and Stemness
4.2. Non-Coding RNA and Stemness
4.3. PAK and Stemness
5. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Harper, J.W.; Adami, G.R.; Wei, N.; Keyomarsi, K.; Elledge, S.J. The p21 Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases. Cell 1993, 75, 805–816. [Google Scholar] [CrossRef]
- El-Deiry, W.S.; Tokino, T.; Velculescu, V.E.; Levy, D.B.; Parsons, R.; Trent, J.M.; Lin, D.; Mercer, W.E.; Kinzler, K.W.; Vogelstein, B. WAF1, a potential mediator of p53 tumor suppression. Cell 1993, 75, 817–825. [Google Scholar] [CrossRef]
- Mousses, S.; Ozçelik, H.; Lee, P.D.; Malkin, D.; Bull, S.B.; Andrulis, I.L. Two variants of the CIP1/WAF1 gene occur together and are associated with human cancer. Hum. Mol. Genet. 1995, 4, 1089–1092. [Google Scholar] [CrossRef] [PubMed]
- Wan, M.; Cofer, K.; Dubeau, L. WAF1/CIP1 structural abnormalities do not contribute to cell cycle deregulation in ovarian cancer. Br. J. Cancer 1996, 73, 1398–1400. [Google Scholar] [CrossRef] [Green Version]
- Diab, S.G.; Yu, Y.Y.; Hilsenbeck, S.G.; Allred, D.C.; Elledge, R.M. WAF1/CIP1 protein expression in human breast tumors. Breast Cancer Res. Treat. 1997, 43, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zou, M.; Farid, N.; al-Sedairy, S. Evidence of gene deletion of p21 (WAF1/CIP1), a cyclin-dependent protein kinase inhibitor, in thyroid carcinomas. Br. J. Cancer 1996, 74, 1336–1341. [Google Scholar] [CrossRef] [Green Version]
- Lukas, J.; Groshen, S.; Saffari, B.; Niu, N.; Reles, A.; Wen, W.H.; Felix, J.; Jones, L.A.; Hall, F.L.; Press, M.F. WAF1/Cip1 gene polymorphism and expression in carcinomas of the breast, ovary, and endometrium. Am. J. Pathol. 1997, 150, 167–175. [Google Scholar]
- Marchetti, A.; Doglioni, C.; Barbareschi, M.; Buttitta, F.; Pellegrini, S.; Bertacca, G.; Chella, A.; Merlo, G.; Angeletti, C.A.; Dalla Palma, P.; et al. p21 RNA and protein expression in non-small cell lung carcinomas: Evidence of p53-independent expression and association with tumoral differentiation. Oncogene 1996, 12, 1319–1324. [Google Scholar]
- Michieli, P.; Chedid, M.; Lin, D.; Pierce, J.H.; Mercer, W.E.; Givol, D. Induction of WAF1/CIP1 by a p53-independent pathway. Cancer Res. 1994, 54, 3391–3395. [Google Scholar]
- Zhang, W.; Grasso, L.; McClain, C.D.; Gambel, A.M.; Cha, Y.; Travali, S.; Deisseroth, A.B.; Mercer, W.E. p53-independent induction of WAF1/CIP1 in human leukemia cells is correlated with growth arrest accompanying monocyte/macrophage differentiation. Cancer Res. 1995, 55, 668–674. [Google Scholar]
- Li, C.-Y.; Suardet, L.; Little, J.B. Potential Role of WAF1 /Cip1/p21 as a Mediator of TGF-β Cytoinhibitory Effect. J. Biol. Chem. 1995, 270, 4971–4974. [Google Scholar] [CrossRef] [PubMed]
- Massagué, J. TGFβ in Cancer. Cell 2008, 134, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Givol, I.; Givol, D.; Rulong, S.; Resau, J.; Tsarfaty, I.; Hughes, S.H. Overexpression of human p21waf1/cip1 arrests the growth of chicken embryo fibroblasts transformed by individual oncogenes. Oncogene 1995, 11, 2609–2618. [Google Scholar] [PubMed]
- Gartenhaus, R.B.; Wang, P.; Hoffmann, P. Induction of the WAF1/CIP1 protein and apoptosis in human T-cell leukemia virus type I-transformed lymphocytes after treatment with adriamycin by using a p53-independent pathway. Proc. Natl. Acad. Sci. USA 1996, 93, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, M.; Tse, V.; Carmel, J.; Costanzi, E.; Strauss, B.; Haas, M.; Silverberg, G.D. Functional Expression of Human p21WAF1/CIP1Gene in Rat Glioma Cells Suppresses Tumor Growthin Vivoand Induces Radiosensitivity. Biochem. Biophys. Res. Commun. 1997, 233, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Georgakilas, A.G.; Martin, O.A.; Bonner, W.M. p21: A Two-Faced Genome Guardian. Trends Mol. Med. 2017, 23, 310–319. [Google Scholar] [CrossRef]
- Abbas, T.; Dutta, A. p21 in cancer: Intricate networks and multiple activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Chen, X. Tumor suppression by p53: Making cells senescent. Histol. Histopathol. 2010, 25, 515–526. [Google Scholar] [CrossRef]
- Romanov, V.S.; Rudolph, K.L. p21 shapes cancer evolution. Nat. Cell Biol. 2016, 18, 722–724. [Google Scholar] [CrossRef]
- Macleod, K.F.; Sherry, N.; Hannon, G.; Beach, D.; Tokino, T.; Kinzler, K.; Vogelstein, B.; Jacks, T. p53-dependent and independent expression of p21 during cell growth, differentiation, and DNA damage. Genes Dev. 1995, 9, 935–944. [Google Scholar] [CrossRef]
- Viale, A.; De Franco, F.; Orleth, A.; Cambiaghi, V.; Giuliani, V.; Bossi, D.; Ronchini, C.; Ronzoni, S.; Muradore, I.; Monestiroli, S.; et al. Cell-cycle restriction limits DNA damage and maintains self-renewal of leukaemia stem cells. Nature 2009, 457, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Marhenke, S.; Buitrago-Molina, L.E.; Endig, J.; Orlik, J.; Schweitzer, N.; Klett, S.; Longerich, T.; Geffers, R.; Sánchez Muñoz, A.; Dorrell, C.; et al. p21 promotes sustained liver regeneration and hepatocarcinogenesis in chronic cholestatic liver injury. Gut 2014, 63, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, W.; Chen, Y.; Huang, Y.; Zhang, J.; He, S.; Tan, Y.; Qiang, F.; Li, A.; Røe, O.D.; et al. The opposite prognostic significance of nuclear and cytoplasmic p21 expression in resectable gastric cancer patients. J. Gastroenterol. 2014, 49, 1441–1452. [Google Scholar] [CrossRef] [PubMed]
- Koster, R.; di Pietro, A.; Timmer-Bosscha, H.; Gibcus, J.H.; van den Berg, A.; Suurmeijer, A.J.; Bischoff, R.; Gietema, J.A.; de Jong, S. Cytoplasmic p21 expression levels determine cisplatin resistance in human testicular cancer. J. Clin. Investig. 2010, 120, 3594–3605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uszczynska-Ratajczak, B.; Lagarde, J.; Frankish, A.; Guigó, R.; Johnson, R. Towards a complete map of the human long non-coding RNA transcriptome. Nat. Rev. Genet. 2018, 19, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef]
- Huarte, M.; Guttman, M.; Feldser, D.; Garber, M.; Koziol, M.J.; Kenzelmann-Broz, D.; Khalil, A.M.; Zuk, O.; Amit, I.; Rabani, M.; et al. A Large Intergenic Noncoding RNA Induced by p53 Mediates Global Gene Repression in the p53 Response. Cell 2010, 142, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.-S.; Zheng, B.-Y.; Xiong, X.-D. LincRNA-p21: Implications in Human Diseases. Int. J. Mol. Sci. 2015, 16, 18732–18740. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Cai, J.; Han, Y.; Chen, J.; Huang, Z.-P.; Chen, C.; Cai, Y.; Huang, H.; Yang, Y.; Liu, Y.; et al. LincRNA-p21 Regulates Neointima Formation, Vascular Smooth Muscle Cell Proliferation, Apoptosis, and Atherosclerosis by Enhancing p53 Activity. Circulation 2014, 130, 1452–1465. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Wu, H.; Zhu, X.; Guo, X.; Hutchins, A.P.; Luo, Z.; Song, H.; Chen, Y.; Lai, K.; Yin, M.; et al. The p53-induced lincRNA-p21 derails somatic cell reprogramming by sustaining H3K9me3 and CpG methylation at pluripotency gene promoters. Cell Res. 2015, 25, 80–92. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, H.; Mei, Y.; Wu, M. Reciprocal Regulation of HIF-1α and LincRNA-p21 Modulates the Warburg Effect. Mol. Cell 2014, 53, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liang, H.; Yang, H.; Zhou, K.; Xu, L.; Liu, J.; Lai, B.; Song, L.; Luo, H.; Peng, J.; et al. LincRNa-p21: Function and mechanism in cancer. Med. Oncol. 2017, 34, 98. [Google Scholar] [CrossRef] [PubMed]
- Puvvula, P.K.; Desetty, R.D.; Pineau, P.; Marchio, A.; Moon, A.; Dejean, A.; Bischof, O. Long noncoding RNA PANDA and scaffold-attachment-factor SAFA control senescence entry and exit. Nat. Commun. 2014, 5, 5323. [Google Scholar] [CrossRef] [PubMed]
- Baldassarre, A.; Masotti, A. Long non-coding RNAs and p53 regulation. Int. J. Mol. Sci. 2012, 13, 16708–16717. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2014, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed]
- Christian, M.; Cermak, T.; Doyle, E.L.; Schmidt, C.; Zhang, F.; Hummel, A.; Bogdanove, A.J.; Voytas, D.F. Targeting DNA Double-Strand Breaks with TAL Effector Nucleases. Genetics 2010, 186, 757–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.G.; Cha, J.; Chandrasegaran, S. Hybrid restriction enzymes: Zinc finger fusions to Fok I cleavage domain. Proc. Natl. Acad. Sci. USA 1996, 93, 1156–1160. [Google Scholar] [CrossRef] [PubMed]
- Robbins, P.D.; Ghivizzani, S.C. Viral vectors for gene therapy. Pharmacol. Ther. 1998, 80, 35–47. [Google Scholar] [CrossRef]
- Fire, A. RNA-triggered gene silencing. Trends Genet. 1999, 15, 358–363. [Google Scholar] [CrossRef]
- Capecchi, M.R. Altering the genome by homologous recombination. Science 1989, 244, 1288–1292. [Google Scholar] [CrossRef]
- Martín-Caballero, J.; Flores, J.M.; García-Palencia, P.; Serrano, M. Tumor Susceptibility of p21 Waf1/Cip1-deficient Mice. Cancer Res. 2001, 61, 6234–6238. [Google Scholar] [PubMed]
- Poole, A.J.; Heap, D.; Carroll, R.E.; Tyner, A.L. Tumor suppressor functions for the Cdk inhibitor p21 in the mouse colon. Oncogene 2004, 23, 8128–8134. [Google Scholar] [CrossRef] [PubMed]
- Eastham, J.A.; Hall, S.J.; Wang, J.; Timme, T.L.; Yang, G.; Connell-crowley, L.; Elledge, S.J.; Zhang, W.; Harper, J.W.; Thompson, T.C. In Vivo Gene Therapy with p53 or p21 Adenovirus for Prostate Cancer. Cancer Res. 1995, 55, 5151–5155. [Google Scholar] [PubMed]
- Tsao, Y.P.; Huang, S.J.; Chang, J.L.; Hsieh, J.T.; Pong, R.C.; Chen, S.L. Adenovirus-mediated p21((WAF1/SDII/CIP1)) gene transfer induces apoptosis of human cervical cancer cell lines. J. Virol. 1999, 73, 4983–4990. [Google Scholar] [PubMed]
- Elliott, M.J.; Stilwell, A.; Dong, B.; Yang, H.L.; Wong, S.L.; Wrightson, W.R.; Martin, R.C.G.; Mcmasters, K.M. C-terminal deletion mutant p21WAF1/CIP1 enhances E2F-1–mediated apoptosis in colon adenocarcinoma cells. Cancer Gene Ther. 2002, 9, 453–463. [Google Scholar] [CrossRef]
- Gorospe, M.; Cirielli, C.; Wang, X.; Seth, P.; Capogrossi, M.C.; Holbrook, N.J. p21 Waf1/Cip1 protects against p53-mediated apoptosis of human melanoma cells. Oncogene 1997, 14, 929–935. [Google Scholar] [CrossRef]
- Jiang, D.; Wang, X.; Liu, X.; Li, F. Gene delivery of cyclin-dependent kinase inhibitors p21 Waf1 and p27 Kip1 suppresses proliferation of MCF-7 breast cancer cells in vitro. Breast Cancer 2014, 21, 614–623. [Google Scholar] [CrossRef]
- García-Fernández, R.A.; García-Palencia, P.; Sánchez, M.Á.; Gil-Gómez, G.; Sánchez, B.; Rollán, E.; Martín-Caballero, J.; Flores, J.M. Combined loss of p21 waf1/cip1 and p27 kip1 enhances tumorigenesis in mice. Lab. Investig. 2011, 91, 1634–1642. [Google Scholar] [CrossRef]
- Okuma, A.; Hanyu, A.; Watanabe, S.; Hara, E. P16Ink4a and p21Cip1/Waf1 promote tumour growth by enhancing myeloid-derived suppressor cells chemotaxis. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Ibnat, N.; Kamaruzman, N.I.; Ashaie, M.; Chowdhury, E.H. Transfection with p21 and p53 tumor suppressor plasmids suppressed breast tumor growth in syngeneic mouse model. Gene 2019, 701, 32–40. [Google Scholar] [CrossRef]
- Bearss, D.J.; Lee, R.J.; Troyer, D.A.; Pestell, R.G.; Windle, J.J. Differential effects of p21(WAF1/CIP1) deficiency on MMTV-ras and MMTV-myc mammary tumor properties. Cancer Res. 2002, 62, 2077–2084. [Google Scholar] [PubMed]
- Ehedego, H.; Boekschoten, M.V.; Hu, W.; Doler, C.; Haybaeck, J.; Gassler, N.; Muller, M.; Liedtke, C.; Trautwein, C. p21 Ablation in Liver Enhances DNA Damage, Cholestasis, and Carcinogenesis. Cancer Res. 2015, 75, 1144–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buitrago-Molina, L.E.; Marhenke, S.; Longerich, T.; Sharma, A.D.; Boukouris, A.E.; Geffers, R.; Guigas, B.; Manns, M.P.; Vogel, A. The degree of liver injury determines the role of p21 in liver regeneration and hepatocarcinogenesis in mice. Hepatology 2013, 58, 1143–1152. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Raina, K.; Agarwal, R. Deletion of p21/Cdkn1a confers protective effect against prostate tumorigenesis in transgenic adenocarcinoma of the mouse prostate model. Cell Cycle 2013, 12, 1598–1604. [Google Scholar] [CrossRef] [PubMed]
- Adnane, J.; Jackson, R.J.; Nicosia, S.V.; Cantor, A.B.; Pledger, W.J.; Sebti, S.M. Loss of p21WAF1/CIP1 accelerates Ras oncogenesis in a transgenic/knockout mammary cancer model. Oncogene 2000, 19, 5338–5347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.C.; Mathew, J.; Velcich, A.; Edelmann, W.; Kucherlapati, R.; Lipkin, M.; Yang, K.; Augenlicht, L.H. Targeted Inactivation of the p21 WAF1/cip1 Gene Enhances Apc-initiated Tumor Formation and the Tumor-promoting Activity of a Western-Style High-Risk Diet by Altering Cell Maturation in the Intestinal Mucosa 1. Cancer Res. 2001, 61, 565–569. [Google Scholar] [PubMed]
- Yeganeh, M.; Gui, Y.; Kandhi, R.; Bobbala, D.; Tobelaim, W.-S.; Saucier, C.; Yoshimura, A.; Ferbeyre, G.; Ramanathan, S.; Ilangumaran, S. Suppressor of cytokine signaling 1-dependent regulation of the expression and oncogenic functions of p21CIP1/WAF1 in the liver. Oncogene 2016, 35, 4200–4211. [Google Scholar] [CrossRef] [PubMed]
- Milewski, D.; Pradhan, A.; Wang, X.; Cai, Y.; Le, T.; Turpin, B.; Kalinichenko, V.V.; Kalin, T.V. FoxF1 and FoxF2 transcription factors synergistically promote Rhabdomyosarcoma carcinogenesis by repressing transcription of p21Cip1 CDK inhibitor David. Oncogene 2017, 36, 850–862. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Li, J.; Liu, Q.; Liu, C.; Li, C.; Song, G.; Zhu, H.; Gao, H.; Zhang, Y. P21 Waf1/Cip1 and p27 Kip1 are correlated with the development and invasion of prolactinoma. J. Neurooncol. 2018, 136, 485–494. [Google Scholar] [CrossRef]
- Kang, M.R.; Park, K.H.; Yang, J.-O.; Lee, C.W.; Oh, S.J.; Yun, J.; Lee, M.Y.; Han, S.-B.; Kang, J.S. miR-6734 Up-Regulates p21 Gene Expression and Induces Cell Cycle Arrest and Apoptosis in Colon Cancer Cells. PLoS ONE 2016, 11, e0160961. [Google Scholar] [CrossRef]
- Valente, L.J.; Grabow, S.; Vandenberg, C.J.; Strasser, A.; Janic, A. Combined loss of PUMA and p21 accelerates c-MYC-driven lymphoma development considerably less than loss of one allele of p53. Oncogene 2016, 35, 3866–3871. [Google Scholar] [CrossRef] [PubMed]
- Herůdková, J.; Paruch, K.; Khirsariya, P.; Souček, K.; Krkoška, M.; Vondálová Blanářová, O.; Sova, P.; Kozubík, A.; Hyršlová Vaculová, A. Chk1 Inhibitor SCH900776 Effectively Potentiates the Cytotoxic Effects of Platinum-Based Chemotherapeutic Drugs in Human Colon Cancer Cells. Neoplasia 2017, 19, 830–841. [Google Scholar] [CrossRef] [PubMed]
- Sohn, D.; Essmann, F.; Schulze-Osthoff, K.; Jänicke, R.U. P21 Blocks Irradiation-Induced Apoptosis Downstream of Mitochondria By Inhibition of Cyclin-Dependent Kinase-Mediated Caspase-9 Activation. Cancer Res. 2006, 66, 11254–11262. [Google Scholar] [CrossRef] [PubMed]
- Wendt, J.; Radetzki, S.; Von Haefen, C.; Hemmati, P.G.; Güner, D.; Schulze-Osthoff, K.; Dörken, B.; Daniel, P.T. Induction of p21CIP/WAF-1 and G2 arrest by ionizing irradiation impedes caspase-3-mediated apoptosis in human carcinoma cells. Oncogene 2006, 25, 972–980. [Google Scholar] [CrossRef] [PubMed]
- King, M.L.; Murphy, L.L. Role of cyclin inhibitor protein p21 in the inhibition of HCT116 human colon cancer cell proliferation by American ginseng (Panax quinquefolius) and its constituents. Phytomedicine 2010, 17, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Ferrandiz, N.; Martin-Perez, J.; Blanco, R.; Donertas, D.; Weber, A.; Eilers, M.; Dotto, P.; Delgado, M.D.; Leon, J. HCT116 cells deficient in p21Waf1 are hypersensitive to tyrosine kinase inhibitors and adriamycin through a mechanism unrelated to p21 and dependent on p53. DNA Repair 2009, 8, 390–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.D.; Honores, E.; Wu, B.; Johnson, S.; Powell, D.; Miranda, M.; McGinnis, J.P.; Discafani, C.; Rabindran, S.K.; Cheng, W.; et al. Synthesis, SAR study and biological evaluation of novel pyrazolo[1,5-a]pyrimidin-7-yl phenyl amides as anti-proliferative agents. Bioorg. Med. Chem. 2009, 17, 2091–2100. [Google Scholar] [CrossRef] [PubMed]
- Stewart, Z.A.; Mays, D.; Pietenpol, J.A. Defective G1-S cell cycle checkpoint function sensitizes cells to microtubule inhibitor-induced apoptosis. Cancer Res. 1999, 59, 3831–3837. [Google Scholar]
- Hauge, S.; Macurek, L.; Syljuåsen, R.G. p21 limits S phase DNA damage caused by the Wee1 inhibitor MK1775. Cell Cycle 2019, 18, 834–847. [Google Scholar] [CrossRef]
- Jackson, R.J.; Adnane, J.; Coppola, D.; Cantor, A.; Sebti, S.M.; Pledger, W.J. Loss of the cell cycle inhibitors p21 Cip1 and p27 Kip1 enhances tumorigenesis in knockout mouse models. Oncogene 2002, 21, 8486–8497. [Google Scholar] [CrossRef]
- Jung, Y.-S.; Qian, Y.; Chen, X. Examination of the expanding pathways for the regulation of p21 expression and activity. Cell. Signal. 2010, 22, 1003–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiffon, C. Histone Deacetylase Inhibition Restores Expression of Hypoxia-Inducible Protein NDRG1 in Pancreatic Cancer. Pancreas 2018, 47, 200–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Yang, Y.; Liu, S.; Lu, J.; Huang, B.; Zhang, Y. HDAC inhibitor PAC-320 induces G2/M cell cycle arrest and apoptosis in human prostate cancer. Oncotarget 2018, 9, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Janyst, K.; Janyst, M.; Siernicka, M.; Lasek, W. Synergistic antitumor effects of histone deacetylase inhibitor scriptaid and bortezomib against ovarian cancer cells. Oncol. Rep. 2018, 39, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Liang, Q.; Yang, X.; Yu, Y.; Shen, X.; Sun, G. Combination of sorafenib and Valproic acid synergistically induces cell apoptosis and inhibits hepatocellular carcinoma growth via down-regulating Notch3 and pAkt. Am. J. Cancer Res. 2017, 7, 2503–2514. [Google Scholar] [PubMed]
- Elbendary, A.A.; Cirisano, F.D.; Cevans, A.; Davis, P.L., Jr.; Iglehart, J.D.; Marks, J.R.; Berchuck, A. Relationship between p21 expression and mutation of the p53 tumor suppressor gene in normal and malignant ovarian epithelial cells. Clin. Cancer Res. 1996, 2, 1571–1575. [Google Scholar] [PubMed]
- Chen, J.; Wu, X.; Lin, J.; Levine, A.J. mdm-2 inhibits the G1 arrest and apoptosis functions of the p53 tumor suppressor protein. Mol. Cell. Biol. 1996, 16, 2445–2452. [Google Scholar] [CrossRef] [Green Version]
- Jeay, S.; Ferretti, S.; Holzer, P.; Fuchs, J.; Chapeau, E.A.; Wartmann, M.; Sterker, D.; Romanet, V.; Murakami, M.; Kerr, G.; et al. Dose and schedule determine distinct molecular mechanisms underlying the efficacy of the p53–MDM2 inhibitor HDM201. Cancer Res. 2018, 78, 6257–6267. [Google Scholar] [CrossRef]
- Andrews, A.; Warner, K.; Rodriguez-Ramirez, C.; Pearson, A.T.; Nor, F.; Zhang, Z.; Kerk, S.; Kulkarni, A.; Helman, J.I.; Brenner, J.C.; et al. Ablation of cancer stem cells by therapeutic inhibition of the MDM2–p53 interaction in mucoepidermoid carcinoma. Clin. Cancer Res. 2019, 25, 1588–1600. [Google Scholar] [CrossRef]
- Al-Ghabkari, A.; Narendran, A. In Vitro Characterization of a Potent p53-MDM2 Inhibitor, RG7112 in Neuroblastoma Cancer Cell Lines. Cancer Biother. Radiopharm. 2019, 34, 252–257. [Google Scholar] [CrossRef]
- Zhou, B.P.; Liao, Y.; Xia, W.; Spohn, B.; Lee, M.H.; Hung, M.C. Cytoplasmic localization of p21 CIP1/WAF1 by Akt-induced phosphorylation in HER-2/neu-overexpressing cells. Nat. Cell Biol. 2001, 3, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.-H.; Pi, J.; Jin, H.; Cai, J.-Y. Oridonin-induced mitochondria-dependent apoptosis in esophageal cancer cells by inhibiting PI3K/AKT/mTOR and Ras/Raf pathways. J. Cell. Biochem. 2019, 120, 3736–3746. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ren, X.; Zhang, L.; Li, Y.; Cheng, B.; Xia, J. Oridonin inhibits oral cancer growth and PI3K/Akt signaling pathway. Biomed. Pharmacother. 2018, 100, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Zheng, X.; Wang, P.; Guo, S. Deguelin exerts anticancer activity of human gastric cancer MGC-803 and MKN-45 cells in vitro. Int. J. Mol. Med. 2018, 41, 3157–3166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daaboul, H.E.; Dagher, C.; Taleb, R.I.; Bodman-Smith, K.; Shebaby, W.N.; El-Sibai, M.; Mroueh, M.A.; Daher, C.F. The chemotherapeutic effect of β-2-himachalen-6-ol in chemically induced skin tumorigenesis. Biomed. Pharmacother. 2018, 103, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Wang, Z.; Wu, Z.; Wei, Y.; Yang, B.; Lou, S. Valtrate from Valeriana jatamansi Jones induces apoptosis and inhibits migration of human breast cancer cells in vitro. Nat. Prod. Res. 2019, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Guan, H.; You, Z.; Wang, C.; Hu, L.; Zhang, L.; Wang, Y.; Chen, S.; Xu, B.; Chen, M. Aloperine executes antitumor effects through the induction of apoptosis and cell cycle arrest in prostate cancer in vitro and in vivo. OncoTargets Ther. 2018, 11, 2735–2743. [Google Scholar] [CrossRef]
- Lu, S.; Ren, C.; Liu, Y.; Epner, D.E. PI3K-Akt signaling is involved in the regulation of p21(WAF/CIP) expression and androgen-independent growth in prostate cancer cells. Int. J. Oncol. 2006, 28, 245–251. [Google Scholar]
- Wu, S.; Cetinkaya, C.; Munoz-Alonso, M.J.; Von Der Lehr, N.; Bahram, F.; Beuger, V.; Eilers, M.; Leon, J.; Larsson, L.G. Myc represses differentiation-induced p21CIP1 expression via Miz-1-dependent interaction with the p21 core promoter. Oncogene 2003, 22, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Pan, S.; Deng, Y.; Fu, J.; Zhang, Y.; Zhang, Z.; Ru, X.; Qin, X. Decreased expression of ARHGAP15 promotes the development of colorectal cancer through PTEN/AKT/FOXO1 axis. Cell Death Dis. 2018, 9. [Google Scholar] [CrossRef]
- Guo, Q.; Xiong, Y.; Song, Y.; Hua, K.; Gao, S. ARHGAP17 suppresses tumor progression and up-regulates P21 and P27 expression via inhibiting PI3K/AKT signaling pathway in cervical cancer. Gene 2019, 692, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Dang, C. CMTM5 is downregulated and suppresses tumour growth in hepatocellular carcinoma through regulating PI3K-AKT signalling. Cancer Cell Int. 2017, 17, 113. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tan, N.; Liao, H.; Pan, G.; Xu, Q.; Zhu, R. High GSTP1 inhibits cell proliferation by reducing Akt phosphorylation and is associated with a better prognosis in hepatocellular carcinoma. Oncotarget 2018, 9, 8957–8971. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Ran, L.; Liu, Y.; Zhong, S.; Zhou, P.; Liao, M.; Fang, W. Knockdown of hnRNP A2/B1 inhibits cell proliferation, invasion and cell cycle triggering apoptosis in cervical cancer via PI3K/AKT signaling pathway. Oncol. Rep. 2018, 39, 939–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeller, K.I.; Jegga, A.G.; Aronow, B.J.; O’Donnell, K.A.; Dang, C.V. An integrated database of genes responsive to the Myc oncogenic transcription factor: Identification of direct genomic targets. Genome Biol. 2003, 4, R69. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Wang, Y.; Liang, W.; Liu, L.; Pan, N.; Deng, H.; Li, L.; Zou, C.; Leung, F.; Zhou, Y. LRH-1 drives hepatocellular carcinoma partially through induction of c-myc and cyclin E1, and suppression of p21. Cancer Manag. Res. 2018, 10, 2389–2400. [Google Scholar] [CrossRef]
- Winchester, E. Bridges, Calvin Blackman. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Chichester, UK, 2002. [Google Scholar]
- Nijman, S.M.B. Synthetic lethality: General principles, utility and detection using genetic screens in human cells. FEBS Lett. 2011, 585, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Origanti, S.; Cai, S.; Munir, A.Z.; White, L.S.; Piwnica-Worms, H. Synthetic lethality of Chk1 inhibition combined with p53 and/or p21 loss during a DNA damage response in normal and tumor cells. Oncogene 2013, 32, 577–588. [Google Scholar] [CrossRef]
- Hofmann, C.; Shepelev, M.; Chernoff, J. The genetics of Pak. J. Cell Sci. 2004, 117, 4343–4354. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.-S.; Manser, E. PAK family kinases: Physiological roles and regulation. Cell. Logist. 2012, 2, 59–68. [Google Scholar] [CrossRef]
- Rane, C.K.; Minden, A. P21 activated kinases: Structure, regulation, and functions. Small GTPases 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Li, D.-Q. PAKs in Human Cancer Progression: From Inception to Cancer Therapeutic to Future Oncobiology. Adv. Cancer Res. 2016, 130, 137–209. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.L.; Chernoff, J. Mouse models of PAK function. Cell. Logist. 2012, 2, 84–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rane, C.K.; Minden, A. P21 activated kinase signaling in cancer. Semin. Cancer Biol. 2019, 54, 40–49. [Google Scholar] [CrossRef]
- Ye, D.Z.; Field, J. PAK signaling in cancer. Cell. Logist. 2012, 2, 105–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandolfi, A.; Stanley, R.F.; Yu, Y.; Bartholdy, B.; Pendurti, G.; Gritsman, K.; Boultwood, J.; Chernoff, J.; Verma, A.; Steidl, U. PAK1 is a therapeutic target in acute myeloid leukemia and myelodysplastic syndrome. Blood 2015, 126, 1118–1127. [Google Scholar] [CrossRef] [Green Version]
- Wells, C.M.; Whale, A.D.; Parsons, M.; Masters, J.R.W.; Jones, G.E. PAK4: A pluripotent kinase that regulates prostate cancer cell adhesion. J. Cell Sci. 2010, 123, 1663–1673. [Google Scholar] [CrossRef]
- Wang, T.; Yu, H.; Hughes, N.W.; Liu, B.; Kendirli, A.; Klein, K.; Chen, W.W.; Lander, E.S.; Sabatini, D.M. Gene Essentiality Profiling Reveals Gene Networks and Synthetic Lethal Interactions with Oncogenic Ras. Cell 2017, 168, 890–903. [Google Scholar] [CrossRef]
- Ahn, H.K.; Jang, J.; Lee, J.; Se, H.P.; Park, J.O.; Park, Y.S.; Lim, H.Y.; Kim, K.-M.; Kang, W.K. P21-Activated Kinase 4 Overexpression in Metastatic Gastric Cancer Patients. Transl. Oncol. 2011, 4, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Chen, X.; Hu, C.; Zhang, D.; Shao, Z.; Jin, Q.; Yang, J.; Xie, H.; Liu, B.; Hu, M.; et al. Synthetic Lethality-based Identification of Targets for Anticancer Drugs in the Human Signaling Network. Sci. Rep. 2018, 8, 8440. [Google Scholar] [CrossRef]
- Xia, W.; Chen, J.-S.; Zhou, X.; Sun, P.-R.; Lee, D.-F.; Liao, Y.; Zhou, B.P.; Hung, M.-C. Phosphorylation/Cytoplasmic Localization of p21Cip1/WAF1 Is Associated with HER2/neu Overexpression and Provides a Novel Combination Predictor for Poor Prognosis in Breast Cancer Patients. Clin. Cancer Res. 2004, 10, 3815–3824. [Google Scholar] [CrossRef] [PubMed]
- Peters, D.; Radine, C.; Reese, A.; Budach, W.; Sohn, D.; Jänicke, R.U. The DEAD-box RNA helicase DDX41 is a novel repressor of p21 WAF1/CIP1 mRNA translation. J. Biol. Chem. 2017, 292, 8331–8341. [Google Scholar] [CrossRef] [PubMed]
- Deng, T.; Yan, G.; Song, X.; Xie, L.; Zhou, Y.; Li, J.; Hu, X.; Li, Z.; Hu, J.; Zhang, Y.; et al. Deubiquitylation and stabilization of p21 by USP11 is critical for cell-cycle progression and DNA damage responses. Proc. Natl. Acad. Sci. USA 2018, 115, 4678–4683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, E.K.; Mungamuri, S.K.; Attie, O.; Kracikova, M.; Sachidanandam, R.; Manfredi, J.J.; Aaronson, S.A. p53-dependent gene repression through p21 is mediated by recruitment of E2F4 repression complexes. Oncogene 2014, 33, 3959–3969. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Quaas, M.; Steiner, L.; Engeland, K. The p53-p21-DREAM-CDE/CHR pathway regulates G2/M cell cycle genes. Nucleic Acids Res. 2016, 44, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, M.; Qian, Y.; Xu, E.; Zhang, J.; Chen, X. Rbm24, an RNA-binding Protein and a Target of p53, Regulates p21 Expression via mRNA Stability. J. Biol. Chem. 2014, 289, 3164–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechara, E.; Davidovic, L.; Melko, M.; Bensaid, M.; Tremblay, S.; Grosgeorge, J.; Khandjian, E.W.; Lalli, E.; Bardoni, B. Fragile X related protein 1 isoforms differentially modulate the affinity of fragile X mental retardation protein for G-quartet RNA structure. Nucleic Acids Res. 2007, 35, 299–306. [Google Scholar] [CrossRef]
- Majumder, M.; House, R.; Palanisamy, N.; Qie, S.; Day, T.A.; Neskey, D.; Diehl, J.A.; Palanisamy, V. RNA-Binding Protein FXR1 Regulates p21 and TERC RNA to Bypass p53-Mediated Cellular Senescence in OSCC. PLOS Genet. 2016, 12, e1006306. [Google Scholar] [CrossRef]
- Hussain, T.; Saha, D.; Purohit, G.; Kar, A.; Kishore Mukherjee, A.; Sharma, S.; Sengupta, S.; Dhapola, P.; Maji, B.; Vedagopuram, S.; et al. Transcription regulation of CDKN1A (p21/CIP1/WAF1) by TRF2 is epigenetically controlled through the REST repressor complex. Sci. Rep. 2017, 7, 11541. [Google Scholar] [CrossRef] [Green Version]
- Ji, H.; Hui, B.; Wang, J.; Zhu, Y.; Tang, L.; Peng, P.; Wang, T.; Wang, L.; Xu, S.; Li, J.; et al. Long noncoding RNA MAPKAPK5-AS1 promotes colorectal cancer proliferation by partly silencing p21 expression. Cancer Sci. 2019, 110, 72–85. [Google Scholar] [CrossRef]
- Tinay, I.; Tan, M.; Gui, B.; Werner, L.; Kibel, A.S.; Jia, L. Functional roles and potential clinical application of miRNA-345-5p in prostate cancer. Prostate 2018, 78, 927–937. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yu, B. MicroRNA-93 promotes cell proliferation by directly targeting P21 in osteosarcoma cells. Exp. Ther. Med. 2017, 13, 2003–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Yao, Y.; Song, Q.; Li, S.; Hu, Z.; Yu, Y.; Hu, C.; Da, X.; Li, H.; Chen, Q.; et al. Up-regulation of miR-95-3p in hepatocellular carcinoma promotes tumorigenesis by targeting p21 expression. Sci. Rep. 2016, 6, 34034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Balan, V.; Kho, D.; Hogan, V.; Nangia-Makker, P.; Raz, A. Galectin-3 regulates p21 stability in human prostate cancer cells. Oncogene 2013, 32, 5058–5065. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, X.; Tang, H.; Jiang, B.; Dou, Y.; Gorospe, M.; Wang, W. NSUN2-Mediated m5C Methylation and METTL3/METTL14-Mediated m6A Methylation Cooperatively Enhance p21 Translation. J. Cell. Biochem. 2017, 118, 2587–2598. [Google Scholar] [CrossRef] [PubMed]
- Lehman, S.L.; Cerniglia, G.J.; Johannes, G.J.; Ye, J.; Ryeom, S.; Koumenis, C. Translational Upregulation of an Individual p21Cip1 Transcript Variant by GCN2 Regulates Cell Proliferation and Survival under Nutrient Stress. PLOS Genet. 2015, 11, e1005212. [Google Scholar] [CrossRef]
- Lee, M.-S.; Seo, J.; Choi, D.Y.; Lee, E.-W.; Ko, A.; Ha, N.-C.; Yoon, J.B.; Lee, H.-W.; Kim, K.P.; Song, J. Stabilization of p21 (Cip1/WAF1) following Tip60-dependent acetylation is required for p21-mediated DNA damage response. Cell Death Differ. 2013, 20, 620. [Google Scholar] [CrossRef]
- Nakakido, M.; Deng, Z.; Suzuki, T.; Dohmae, N.; Nakamura, Y.; Hamamoto, R. PRMT6 increases cytoplasmic localization of p21CDKN1A in cancer cells through arginine methylation and makes more resistant to cytotoxic agents. Oncotarget 2015, 6, 30957–30967. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Chen, J.; Ning, D.; Liu, Q.; Wang, C.; Zhang, Z.; Chu, L.; Yu, C.; Liang, H.; Zhang, B.; et al. FBXO22 promotes the development of hepatocellular carcinoma by regulating the ubiquitination and degradation of p21. J. Exp. Clin. Cancer Res. 2019, 38. [Google Scholar] [CrossRef]
- Ji, S.Q.; Zhang, Y.X.; Yang, B.H. UBR5 promotes cell proliferation and inhibits apoptosis in colon cancer by destablizing P21. Pharmazie 2017, 72, 408–413. [Google Scholar] [CrossRef]
- Li, Y.; Huang, J.; Zeng, B.; Yang, D.; Sun, J.; Yin, X.; Lu, M.; Qiu, Z.; Peng, W.; Xiang, T.; et al. PSMD2 regulates breast cancer cell proliferation and cell cycle progression by modulating p21 and p27 proteasomal degradation. Cancer Lett. 2018, 430, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Stier, S.; Cheng, T.; Forkert, R.; Lutz, C.; Dombkowski, D.M.; Zhang, J.L.; Scadden, D.T. Ex vivo targeting of p21Cip1/Waf1 permits relative expansion of human hematopoietic stem cells. Blood 2003, 102, 1260–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, T.; Rodrigues, N.; Shen, H.; Yang, Y.; Dombkowski, D.; Sykes, M.; Scadden, D.T. Hematopoietic Stem Cell Quiescence Maintained by p21cip1/waf1. Science 2000, 287, 1804–1808. [Google Scholar] [CrossRef] [PubMed]
- Herreros-Villanueva, M.; Zhang, J.S.; Koenig, A.; Abel, E.V.; Smyrk, T.C.; Bamlet, W.R.; De Narvajas, A.A.; Gomez, T.S.; Simeone, D.M.; Bujanda, L.; et al. SOX2 promotes dedifferentiation and imparts stem cell-like features to pancreatic cancer cells. Oncogenesis 2013, 2, e61. [Google Scholar] [CrossRef]
- Han, S.; Woo, J.K.; Jung, Y.; Jeong, D.; Kang, M.; Yoo, Y.-J.; Lee, H.; Oh, S.H.; Ryu, J.-H.; Kim, W.-Y. Evodiamine selectively targets cancer stem-like cells through the p53-p21-Rb pathway. Biochem. Biophys. Res. Commun. 2016, 469, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Baharuddin, P.; Satar, N.; Fakiruddin, K.S.; Zakaria, N.; Lim, M.N.; Yussof, N.M.; Zakaria, Z.; Yahayah, B.H. Curcumin improves the efficacy of cisplatin by targeting cancer stem-like cells through p21 and cyclin D1-mediated tumour cell inhibition in non-small cell lung cancer cell lines. Oncol. Rep. 2016, 35, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Singh, S.V. Role of Krüppel-like Factor 4-p21 CIP1 Axis in Breast Cancer Stem-like Cell Inhibition by Benzyl Isothiocyanate. Cancer Prev. Res. 2019, 12, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Benard, O.; Qian, X.; Liang, H.; Ren, Z.; Suyama, K.; Norton, L.; Hazan, R.B. p21CIP1 Promotes Mammary Cancer-Initiating Cells via Activation of Wnt/TCF1/CyclinD1 Signaling. Mol. Cancer Res. 2019. [Google Scholar] [CrossRef]
- Yang, W.; Yu, H.; Shen, Y.; Liu, Y.; Yang, Z.; Sun, T. MiR-146b-5p overexpression attenuates stemness and radioresistance of glioma stem cells by targeting HuR/lincRNA-p21/β-catenin pathway. Oncotarget 2016, 7, 41505–41526. [Google Scholar] [CrossRef]
- Wang, J.; Lei, Z.; Guo, Y.; Wang, T.; Qin, Z.; Xiao, H.; Fan, L.; Chen, D.; Bian, X.; Liu, J.; et al. miRNA-regulated delivery of lincRNA-p21 suppresses β-catenin signaling and tumorigenicity of colorectal cancer stem cells. Oncotarget 2015, 6, 37852–37870. [Google Scholar] [CrossRef]
- Chang, Y.-L.; Zhou, P.-J.; Wei, L.; Li, W.; Ji, Z.; Fang, Y.-X.; Gao, W.-Q. MicroRNA-7 inhibits the stemness of prostate cancer stem-like cells and tumorigenesis by repressing KLF4/PI3K/Akt/p21 pathway. Oncotarget 2015, 6, 24017–24031. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-Y.; Yang, M.-C.; Ding, L.-Y.; Chen, C.S.; Chu, P.-C. p21-Activated kinase 3 promotes cancer stem cell phenotypes through activating the Akt-GSK3β-β-catenin signaling pathway in pancreatic cancer cells. Cancer Lett. 2019, 456, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Huynh, N.; Shulkes, A.; Baldwin, G.; He, H. Up-regulation of stem cell markers by P21-activated kinase 1 contributes to 5-fluorouracil resistance of colorectal cancer. Cancer Biol. Ther. 2016, 17, 813–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, N.; Marimuthu, S.; Bhardwaj, A.; Deshmukh, S.K.; Srivastava, S.K.; Singh, A.P.; McClellan, S.; Carter, J.E.; Singh, S. p-21 activated kinase 4 (PAK4) maintains stem cell-like phenotypes in pancreatic cancer cells through activation of STAT3 signaling. Cancer Lett. 2016, 370, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Gómez, A.; Kedward, T.; Simões, B.M.; Dragoni, I.; NicAmhlaoibh, R.; Trivier, E.; Sabin, V.; Gee, J.M.; Sims, A.H.; Howell, S.J.; et al. PAK4 regulates stemness and progression in endocrine resistant ER-positive metastatic breast cancer. Cancer Lett. 2019, 458, 66–75. [Google Scholar] [CrossRef]
- Gartel, A.L.; Tyner, A.L. The role of the cyclin-dependent kinase inhibitor p21 in apoptosis. Mol. Cancer Ther. 2002, 1, 639–649. [Google Scholar]
- Parveen, A.; Akash, M.S.H.; Rehman, K.; Kyunn, W.W. Dual Role of p21 in the Progression of Cancer and Its Treatment. Crit. Rev. Eukaryot. Gene Expr. 2016, 26, 49–62. [Google Scholar] [CrossRef]
- Palmer, A.C.; Sorger, P.K. Combination Cancer Therapy Can Confer Benefit via Patient-to-Patient Variability without Drug Additivity or Synergy. Cell 2017, 171, 1678–1691.e13. [Google Scholar] [CrossRef] [Green Version]
- Galanos, P.; Vougas, K.; Walter, D.; Polyzos, A.; Maya-Mendoza, A.; Haagensen, E.J.; Kokkalis, A.; Roumelioti, F.-M.; Gagos, S.; Tzetis, M.; et al. Chronic p53-independent p21 expression causes genomic instability by deregulating replication licensing. Nat. Cell Biol. 2016, 18, 777–789. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Kwiatkowski, D.J. Combined CDKN1A/TP53 Mutation in Bladder Cancer Is a Therapeutic Target. Mol. Cancer Ther. 2015, 14, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Idogawa, M.; Sasaki, Y.; Suzuki, H.; Mita, H.; Imai, K.; Shinomura, Y.; Tokino, T. A Single Recombinant Adenovirus Expressing p53 and p21-targeting Artificial microRNAs Efficiently Induces Apoptosis in Human Cancer Cells. Clin. Cancer Res. 2009, 15, 3725–3732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, Y.; Tazawa, H.; Hashimoto, Y.; Kojima, T.; Kuroda, S.; Yano, S.; Yoshida, R.; Uno, F.; Mizuguchi, H.; Ohtsuru, A.; et al. A novel apoptotic mechanism of genetically engineered adenovirus-mediated tumour-specific p53 overexpression through E1A-dependent p21 and MDM2 suppression. Eur. J. Cancer 2012, 48, 2282–2291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccolo, M.T.; Crispi, S. The Dual Role Played by p21 May Influence the Apoptotic or Anti-Apoptotic Fate in Cancer. J. Can. Res. Updates 2012, 1, 189–202. [Google Scholar] [Green Version]
- Gawriluk, T.R.; Simkin, J.; Thompson, K.L.; Biswas, S.K.; Clare-Salzler, Z.; Kimani, J.M.; Kiama, S.G.; Smith, J.J.; Ezenwa, V.O.; Seifert, A.W. Comparative analysis of ear-hole closure identifies epimorphic regeneration as a discrete trait in mammals. Nat. Commun. 2016, 7, 11164. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Phenotype of Mice | Cancer Model | Description | Reference |
|---|---|---|---|
| p21−/− | Hematopoietic tumor Epithelial tumor Endothelial tumor | Susceptible to spontaneous tumors development | [41] |
| p21−/− | Colon cancer | Increased putative premalignant lesions development | [42] |
| p21−/− p16−/− | Lewis lung carcinoma | Inhibition of MO-MDSC, accumulation in tumors, and suppression of tumor progression | [49] |
| p21−/− p27−/− | Pituitary adenomas, Pheochromocytomas Thyroid adenomas | Aggressive tumor and decreased lifespan | [48] |
| p21−/− | HCT116 | Combination of Chk1 inhibitors and cisplatin treatment enhances cancer cell vulnerability | [62] |
| p21−/− | HCT116 | Caspase-9 and caspase-3 dependent apoptosis after ionizing radiation | [63] |
| p21−/− | HCT116 | Enhanced caspase-3-dependent apoptosis after irradiation | [64] |
| p21−/− | HCT116 | Increased apoptosis and cell death with treatment of ginseng, imatinib and gefitinib, pyrazolo(1,5-a)pyrimidine, microtubule inhibitors, MK1775 checkpoint inhibitor | [65] [66] [67] [68] [69] |
| p21−/− | MMTV-ras | Higher S-phase fractions, increased spontaneous apoptosis | [51] |
| p21−/− | MMTV-myc | Lower S-phase fractions, no effect on apoptosis | [51] |
| NEMOΔhepa/p21−/− | HCC | Higher DNA damage and higher number of HCC | [52] |
| p21−/− | Lung tumors | Accelerated tumor onset, increased tumor multiplicity | [70] |
| Fah−/−/p21−/− | HCC | Rapid tumor formation, continuous hepatocyte proliferation | [53] |
| p21−/− | Prostate cancer | Less aggressiveness in prostates, lower adenocarcinoma incidence and prostate tumorigenesis | [54] |
| p21−/− | MMTV/v-Ha-ras | Increase in aggressiveness and tumor multiplicity, earlier tumor appearance | [55] |
| Apc1638+/− / p21−/− | Intestinal tumor | Increased tumor formation | [56] |
| p21−/− | miR-6734 expressing HCT-116 cells | Tumor growth and not induction of cell cycle arrest and apoptosis, phosphorylation of Rb and cleavage of PARP and caspase 3 | [60] |
| (SOCS)1−/− /p21−/− | HCC | No increased resistance to apoptosis and no increased proliferation to growth factor stimulation | [57] |
| FoxF1−/− / FoxF2−/− / p21−/− | RMS | Restored cell cycle progression | [58] |
| p21−/− | c-MYC-driven lymphoma | No change on the number and survival of pre-leukemic Eμ-Myc B-lymphoid cells | [61] |
| p21−/− | PRL | Increase in migration and tumor formation | [59] |
| Drug/Chemical | Target/Pathway | Cancer Type | Reference |
|---|---|---|---|
| β-2-himachalen-6-ol | Akt/PI3K-Akt | Skin Carcinogenesis | [85] |
| Valtrate | Akt/PI3K-Akt | Breast Cancer | [86] |
| Deguelin | Akt/PI3K-Akt | Gastric Cancer | [84] |
| Aloperine | Akt/PI3K-Akt | Prostate Cancer | [87] |
| Oridonin Oridonin | Akt/PI3K-Akt Akt/PI3K-Akt | Esophageal Cancer Oral Squamous Cell Carcinoma | [82] [83] |
| HDM201 | MDM2-p53 binding | p53 Wild- Type Cancers | [78] |
| MI-773 | MDM2-p53 binding | Mucoepidermoid Carcinoma | [79] |
| RG7112 | MDM2-p53 binding | Neuroblastoma Cancer | [80] |
| Trichostatin A | HDAC | Pancreatic Cancer | [72] |
| PAC-320 | HDAC | Prostate Cancer | [73] |
| Scriptaid+Bortezomib | HDAC | Ovarian Cancer | [74] |
| Scriptaid + Doxorubicin | HDAC | Ovarian Cancer | [74] |
| Sorafenib+Valproic Acid | HDAC | HCC | [75] |
| Gene/Protein/lncRNA/miRNA | Expression in Cancer Cells | Target/Pathway | Cancer Type | Reference |
|---|---|---|---|---|
| ARHGAP15 | Downregulated | Akt/PI3K-Akt | CRC | [90] |
| ARHGAP17 | Downregulated | Akt/PI3K-Akt | Cervical Cancer | [91] |
| CMTM5 | Downregulated | Akt/PI3K-Akt | HCC | [92] |
| GSTP1 | Downregulated | Akt/PI3K-Akt | HCC | [93] |
| hnRNP A2/B1 | Overexpressed | Akt/PI3K-Akt | Cervical Cancer | [94] |
| LRH-1 | Overexpressed | c-Myc/c-Myc-p21 | HCC | [96] |
| FBXO22 | Overexpressed | p21/ubiquitylation | HCC | [130] |
| UBR5 | Overexpressed | p21/ubiquitylation | Colon Cancer | [131] |
| PSMD2 FXR1 MAPKAPK5-AS1 miR-345-5p miR-93 miR-95-3p | Overexpressed Overexpressed Overexpressed Overexpressed Overexpressed Overexpressed | p21/ubiquitylation p21/Posttranscriptional p21/Posttranscriptional p21/Posttranscriptional p21/Translational p21/Posttranscriptional | Breast Cancer HNSCC CRC Prostate Cancer Osteosarcoma Cells HCC | [132] [119] [121] [122] [123] [124] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shamloo, B.; Usluer, S. p21 in Cancer Research. Cancers 2019, 11, 1178. https://doi.org/10.3390/cancers11081178
Shamloo B, Usluer S. p21 in Cancer Research. Cancers. 2019; 11(8):1178. https://doi.org/10.3390/cancers11081178
Chicago/Turabian StyleShamloo, Bahar, and Sinem Usluer. 2019. "p21 in Cancer Research" Cancers 11, no. 8: 1178. https://doi.org/10.3390/cancers11081178
APA StyleShamloo, B., & Usluer, S. (2019). p21 in Cancer Research. Cancers, 11(8), 1178. https://doi.org/10.3390/cancers11081178